Carnotaurus sastrei
Carnotaurus sastrei (lat. "toro carnívoro de Anselmo Sastre"),[1] o carnotauro en castellano, es la única especie conocida del género extinto Carnotaurus de dinosaurio terópodo de la familia Abelisauridae, del período Cretácico, hace aproximadamente entre 72 y 69,9 millones de años, desde el Campaniense al Maastrichtiense, en lo que hoy es Sudamérica.[2] Lo más notable de este dinosaurio son sus dos pequeños cuernos sobre los ojos en su pequeña cabeza, siendo el primer carnívoro que mostrara cuernos bien formados y definidos. También se hallaron impresiones de la piel que presentaban pequeños osteodermos, escudos óseos.[1] Carnotaurus es un miembro avanzado de la familia Abelisauridae, un grupo de grandes terópodos que ocuparon el nicho de gran depredador en las masas de tierra que conformaron Gondwana durante finales del Cretácico, ocupando el lugar de los tiranosáuridos luego de la desaparición de los carcarodontosáuridos y neovenatóridos.[3]
| Carnotaurus sastrei | ||
|---|---|---|
| Rango temporal: 72 Ma - 69,9 Ma Cretácico Superior | ||
.png.webp) Reconstrucción en vida. | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Sauropsida | |
| Superorden: | Dinosauria | |
| Orden: | Saurischia | |
| Suborden: | Theropoda | |
| Infraorden: | Ceratosauria | |
| Superfamilia: | Abelisauroidea | |
| Familia: | Abelisauridae | |
| Subfamilia: | Carnotaurinae | |
| Tribu: | Carnotaurini | |
| Género: |
Carnotaurus Bonaparte, 1985 | |
| Especie: |
C. sastrei Bonaparte, 1985 | |
Junto con el género Abelisaurus, dieron a conocer una familia completamente nueva de predadores mesozoicos, los abelisáuridos, lo que llevó a la revisión taxonómica de muchos otros géneros de difícil clasificación con el engrandecimiento del clado ceratosauria y a una visión distinta sobre la fauna gondwánica a finales de la época de los dinosaurios.[1] Su nombre proviene del latín carō, carnis, "carne", y taurus, "toro", y le fue dado por ser un carnívoro con aspecto de toro, debido a los apéndices de su cabeza en forma de cuernos. El nombre de la especie "sastrei" le fue otorgado en honor a Anselmo Sastre, dueño de la estancia en la provincia argentina de Chubut donde fue hallado el primer y único espécimen.[1]
Los hábitos alimentarios de Carnotaurus aún no están claros del todo para los investigadores, aunque algunos estudios de la morfología craneal sugieren que el animal era capaz de cazar presas muy grandes, como los saurópodos. Otros estudios encontraron que se alimentaba principalmente de animales relativamente pequeños. Lo que sí se sabe es que Carnotaurus se adaptaba muy bien para correr y fue posiblemente uno de los terópodos más rápidos de gran tamaño.
Descripción

Carnotaurus era un terópodo de tamaño medio a grande, pero de constitución ligera,[3] de entre ocho y nueve metros de longitud,[4] alrededor de 3,5 de altura, y pesaba según los estudios entre 1350 y 2100 kilogramos.[5][6][7] Se estima que su masa fue de 1350 kilogramos,[5] 1500 kilogramos,[6] 2000 kilogramos,[8] 2100 kilogramos,[7] y entre 1306 y 1743 kilogramos[9] en estudios separados que utilizaron diferentes métodos de estimación. Esto lo vuelve uno de los abelisáuridos más grandes conocidos; solo Pycnonemosaurus y posiblemente Abelisaurus pudieron ser de mayor tamaño, aunque los restos incompletos de estos géneros hacen que las estimaciones sean imprecisas.[10][11][4] Su cabeza era muy pequeña en comparación con las de los otros predadores de su tamaño; alrededor de sesenta centímetros de largo.[1][5] Las características más distintivas del Carnotaurus son dos cuernos gruesos sobre los ojos y unos miembros delanteros extremadamente reducidos y atrofiados, provistos de cuatro dedos.[1] Estudios separados, que utilizan diferentes métodos de estimación, muestran a Carnotaurus un altamente especializado terópodo, lo que se denota en características del cráneo, las vértebras y las extremidades anteriores.[12] La pelvis y extremidades posteriores, por otra parte, mantuvieron una estructura relativamente conservadora, parecida a las del más basal Ceratosaurus. Tanto la pelvis como los huesos de los miembros posteriores eran largos y delgados. El hueso del muslo izquierdo mide 103 cm de longitud, pero muestra un diámetro promedio de solo once centímetros.[5]
Cabeza

Tenía un cráneo fuertemente construido, pero pequeño, con espacios vacíos a sus lados para disminuir el peso, un pecho grueso, y una cola gruesa y larga que le proveía de equilibrio. Los pequeños ojos de Carnotaurus, que se encontraban en unas órbitas oculares fuertemente protegidas, miraban hacia el frente, algo que es inusual en un dinosaurio, lo que puede indicar que estos animales tenían visión binocular y percepción de profundidad. El masivo hocico sugiere que habrían presentado órganos olfativos de gran tamaño, lo que apoya la idea de que el animal tenía un olfato refinado. Las mandíbulas poseían una articulación que le permitía ampliar el área de la mordida.[5]
El cráneo, que mide 59,6 cm de longitud, era proporcionalmente más corto y profundo que en cualquier otro gran dinosaurio carnívoro.[5][11] El hocico era amplio, a diferencia del visto en terópodos más basales como Ceratosaurus, y las mandíbulas estaban curvadas hacia arriba.[13] Al igual que en otros abelisáuridos, los huesos de la cara, especialmente los huesos nasales, estaban esculpidos con numerosos pequeños agujeros y espigas. En vida, una piel gruesa y posiblemente queratinosa habría cubierto estos huesos.[5] Un par de prominentes cuernos sobresalían oblicuamente por encima de los ojos. Estos cuernos, formados por los huesos frontales,[5] eran gruesos, aplanados en sus lados superiores, y medían quince cm de longitud.[14] En vida, probablemente se han formado los núcleos óseos de cuernos de queratina mucho más grandes.[5] Mauricio Cerroni y sus colegas, en 2020, acordaron que los cuernos soportaban vainas queratinosas, pero argumentaron que estas vainas no habrían sido mucho más largas que los núcleos óseos.[15] Los ojos, proporcionalmente pequeños,[16] estaban situados en la parte superior de la órbita ocular con forma de ojo de cerradura.[5] La parte superior estaba ligeramente girada hacia delante, probablemente permitiendo cierto grado de visión binocular.[6] La mandíbula inferior de Carnotaurus muestra un débil contacto de los huesos dentario y postdentario, formando una larga ventana en la mandíbula. Los caracteres derivados de las vértebras cervicales, tales como procesos espinosos reducidos y epipofísis grandes y altas. Características únicas de Carnotaurus son el yugal ancho, corto y alto, y los cuernos visibles hechos por una extensión latero dorsal corta y saliente de cada frontal.[17][5][18]

Como en otros dinosaurios, el cráneo estaba perforado por seis aberturas principales en cada lado. La más frontal de estas aberturas, la cavidad nasal externa, fosa nasal ósea, era subrectangular y estaba dirigida hacia los lados y hacia delante, pero no se inclinaba en la vista lateral como en algunos otros ceratosaurianos, como Ceratosaurus. Esta abertura estaba formada solo por el nasal y el premaxilar, mientras que en algunos ceratosaurianos relacionados el maxilar también contribuía a esta abertura. Entre la fosa nasal ósea y la órbita, abertura del ojo, estaba la fenestra antorbitaria. En Carnotaurus, esta abertura era más alta que larga, mientras que era más larga que alta en formas relacionadas como Skorpiovenator y Majungasaurus. La fenestra antorbitaria estaba limitada por una depresión más grande, la fosa antorbitaria, que estaba formada por partes rebajadas del maxilar al frente y el lagrimal por detrás. Como en todos los abelisáuridos, esta depresión fue pequeña en Carnotaurus. La esquina frontal inferior de la fosa antorbitaria contenía una abertura más pequeña, la fenestra promaxilar, que conducía a una cavidad llena de aire dentro del maxilar.[15] El ojo estaba situado en la parte superior de la órbita en forma de ojo de cerradura.[5] Esta parte superior era proporcionalmente pequeña y subcircular, y estaba separada de la parte inferior de la órbita por el hueso postorbitario que se proyectaba hacia delante.[1] Estaba ligeramente girado hacia delante, probablemente permitiendo cierto grado de visión binocular.[15] La forma de ojo de cerradura de la órbita posiblemente estaba relacionada con el marcado acortamiento del cráneo, y también se encuentra en abelisáuridos de hocico corto relacionados.[15] Como en todos los abelisáuridos, el hueso frontal, en el techo del cráneo entre los ojos, fue excluido de la órbita. Detrás de la órbita había dos aberturas, la fenestra infratemporal en el lateral y la fenestra supratemporal en la parte superior del cráneo. La fenestra infratemporal era alta, corta y con forma de riñón, mientras que la fenestra supratemporal era corta y de forma cuadrada. Otra abertura, la fenestra mandibular, estaba ubicada en la mandíbula inferior; en Carnotaurus, esta abertura era comparativamente grande.[15]
Los dientes eran largos y delgados,[16] en comparación con los dientes por lo general muy cortos vistos en otros abelisáuridos.[13] A cada lado de la mandíbula superior había cuatro dientes premaxilares y doce maxilares,[12] mientras que la mandíbula inferior se equipaba con quince dientes dentarios por lado.[5] En contraste con el robusto cráneo, la mandíbula inferior era delgada y débilmente construida, con el dentario conectado a los huesos de la mandíbula inferior por solo dos puntos de contacto.[16][5] La mandíbula inferior se encontró con huesos hioides en la posición que tendrían en caso de que el animal estuviera vivo. Estos huesos delgados son el apoyo a la musculatura de la lengua y varios otros músculos, y rara vez se encuentran en los dinosaurios, ya que no están conectados a otros huesos y por lo tanto se pierden fácilmente.[5][19] Sin embargo, Cerroni y sus colegas, en su descripción de 2020 del cráneo, afirmaron que todos los dientes erupcionados habían sido severamente dañados durante la excavación y luego fueron reconstruidos con yeso. Bonaparte, en 1990, solo notó que algunos dientes de la mandíbula inferior se habían fragmentado.[15][5] Por lo tanto, la información confiable sobre la forma de los dientes se limita a los dientes de reemplazo y las raíces de los dientes que aún están encerrados en la mandíbula, y se puede estudiar mediante imágenes de tomografía computarizada.[15] Los dientes de reemplazo tenían coronas bajas y aplanadas, estaban poco espaciados e inclinados hacia adelante, aproximadamente a 45 °.[15] En su descripción de 1990, Bonaparte notó que la mandíbula inferior era poco profunda y de estructura débil, con el dentario, el hueso de la mandíbula más adelantado, conectado a los huesos de la mandíbula posterior por sólo dos puntos de contacto; esto contrasta con el cráneo de aspecto robusto.[5][19][15] En cambio, Cerroni y sus colegas encontraron conexiones múltiples pero sueltas entre el hueso dentario y el último de la mandíbula. Esta articulación, por tanto, fue muy flexible pero no necesariamente débil.[15] El margen inferior del dentario era convexo, mientras que en Majungasaurus era recto.[15]
La mandíbula inferior se encontró con huesos hioides osificados en la posición en la que estarían si el animal estuviera vivo. Estos huesos delgados, que sostienen la musculatura de la lengua y varios otros músculos, rara vez se encuentran en los dinosaurios porque a menudo son cartilaginosos y no están conectados a otros huesos; por lo tanto, se pierden fácilmente.[5][19][15] En Carnotaurus se conservan tres huesos hioides, un par de ceratobranquiales curvos en forma de varilla que se articulan con un único elemento trapezoidal, el basihyal. Carnotaurus es el único terópodo no aviar conocido del que se conoce un basihyal.[15] La parte posterior del cráneo tenía cámaras bien desarrolladas y llenas de aire que rodeaban la caja del cerebro, como en otros abelisáuridos. Estaban presentes dos sistemas de cámara separados: el sistema paratimpánico, que estaba conectado a la cavidad del oído medio, así como cámaras resultantes de excrecencias de los sacos aéreos del cuello.[9]
Se pueden encontrar varias autapomorfias, características distintivas, en el cráneo, incluido el par de cuernos y el cráneo muy corto y profundo. El maxilar tenía excavaciones por encima de la fenestra promaxilar, que habría sido excavada por el seno aéreo antorbitario, conductos de aire en el hocico. El conducto nasolagrimal, que transportaba el líquido ocular, salía por la superficie medial, interior, del lagrimal a través de un canal de función incierta. Otras autapomorfias propuestas incluyen una profunda y larga excavación llena de aire en el cuadrado y una depresión alargada en el pterigoideo del paladar.[15]
Vértebras
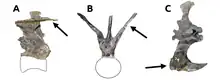
La columna vertebral consistió en diez vértebras cervicales formando el cuello, doce dorsales en la espalda, seis sacras fusionadas[11] y un número desconocido de vértebras de la cola.[5] El cuello era casi recto, en vez de tener la curva en S como se ve en otros terópodos, y también inusualmente ancho, sobre todo hacia su base.[20] La parte superior de la columna vertebral del cuello ofrece una doble fila de procesos óseos agrandados, dirigidos hacia arriba, llamados epipófisis, haciendo que la parte superior del mismo sea plana. Estos procesos fueron los puntos más altos de la columna vertebral; se elevan por encima de las apófisis espinosas inusualmente bajas.[5][19] Las epipófisis probablemente fueron zonas de fijación para una musculatura del cuello marcadamente fuerte.[12] Una fila doble similar también estuvo presente en la cola, formada allí por los procesos transversos altamente modificados, llamados costillas caudales, que sobresalen en forma de V, resultando en una parte superior plana de la cola. El extremo de cada costilla caudal estaba equipado con una expansión que se proyecta hacia delante que conecta con la de la vértebra anterior.[19][21]
Piel

Solo un esqueleto, aunque casi completo, ha sido encontrado, incluyendo impresiones de la piel a lo largo de casi todo el costado derecho, que no muestra evidencia de plumas u otras estructuras filamentosas, como en otros terópodos. La piel está incrustada por pequeños osteodermos, delineados en columnas de barriles paralelos al eje central del cuerpo. Estos tienen forma de prominencias cónicas de unos cinco centímetros de diámetro, que aumentan en tamaño al acercarse a la línea central de la espalda, lo que le otorga un aspecto del estilo de un cocodrilo actual, y sugiere que el Carnotaurus presentaba una piel áspera y gruesa.[1]
Carnotaurus fue el primer dinosaurio terópodo descubierto con impresiones completas de piel fósil.[22] Estas impresiones, que se encuentran por debajo de la parte derecha del esqueleto, provienen de diferentes partes del cuerpo, incluyendo la mandíbula inferior,[22] la parte frontal del cuello, la cintura escapular y la caja torácica.[5] La mayor parte de la piel corresponde a la parte anterior de la cola.[5] Originalmente, el lado derecho del cráneo también estaba cubierto de grandes parches de la piel; esto no fue reconocido cuando el cráneo estaba siendo preparado, y estos fueron destruidos accidentalmente. Sin embargo, la textura de la superficie de la mayor parte del lado derecho del cráneo es muy diferente de la parte izquierda, y probablemente muestra algunas de las características del patrón de escamas de la cabeza.[22]

La piel estaba constituida por un mosaico de escamas poligonales, que no se superponen entre sí y miden aproximadamente cinco mm de diámetro. Este mosaico se dividió por ranuras paralelas, finas.[5] El patrón de escamas fue similar a través de las diferentes partes del cuerpo con la excepción de la cabeza, que al parecer mostró un patrón diferente, con escamas irregulares.[5][23] No hay evidencia de plumas.[22] De manera única para los terópodos, tenía osteodermos, escudos óseos, a lo largo de los lados del cuello, la espalda y la cola en filas irregulares. Cada uno de estos mostró una cresta baja y mide cuatro-cinco cm de diámetro, estando espaciados entre ocho y diez cm de distancia el uno del otro y siendo más grandes hacia la parte superior del animal. Las protuberancias probablemente representan racimos de condensados escudos, similares a los observados a lo largo de la línea media del cuerpo en hadrosáuridos.[5][22] Stephen Czerkas sugiere que estas estructuras podrían proteger los lados del animal mientras luchaba contra miembros de su misma especie u otros terópodos, con el argumento de que las estructuras similares se pueden encontrar en el cuello de la moderna iguana, donde proporcionan una protección limitada en el combate.[22]
Brazos
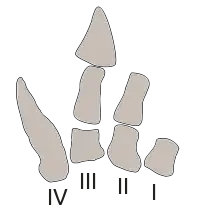
Sus extremidades anteriores eran más cortas en comparación con las del Tyrannosaurus,[24] con cuatro dedos en cada mano,[5] cosa considerada un rasgo primitivo entre los dinosaurios. No había carpo en la mano, de modo que los metacarpianos articulaban directamente con el antebrazo.[24] La mano mostraba los cuatro dígitos básicos,[5] aunque aparentemente solo los dos medios de estos terminaron en huesos de los dedos con el cuarto dedo constituido solo por el metacarpiano con forma de espolón. Se cree que este último era usado para sujetar a las presas.[1] Los dedos mismos estaban fusionados e inmóviles, y pueden haber carecido de garras.[25] Carnotaurus se diferenciaba de todos los demás abelisáuridos en tener patas delanteras proporcionalmente más cortas y robustas, y en tener el cuarto metacarpo como el hueso más largo en la mano.[24]

La proporción de los miembros superiores del Carnotaurus con respecto de los inferiores es del 14 % mientras que en el Albertosaurus esta relación era del 20 %, 46 % en Struthiomimus, 61 % en Deinonychus, y 96 % en Archaeopteryx. Las medidas de los huesos del brazo eran de 285 milímetros el húmero, 73 mm el radio y 37 mm el metacarpo, comparados con los 324, 156 y 98 del Albertosaurus y 357, 171 y 120 del Daspletosaurus respectivamente,[26] los dos tiranosáuridos de dimensiones similares y misma edad. Carnotaurus era el terópodo con la reducción más acentuada de los miembros superiores, mostrando una mayor reducción de los huesos distales del miembro. En proporción, el húmero representaba el 72 %, el radio el 19 % y el metacarpo solo el 9 %, comparados con otros terópodos con miembros reducidos que oscilaban alrededor de las siguientes proporciones: 55 %, 27 % y 19 % respectivamente.[26] Además, se ha visto que las falanges proximales de los dedos II y III eran mayores que sus respectivos metacarpianos.[24] Se ha sugerido otra extraña adaptación en los miembros superiores; según algunos investigadores, los huesos del brazo giraban 180°, dejando las palmas de las manos hacia fuera y no contra el cuerpo como en otros grandes terópodos.[27] Un estudio de 2009 sugiere que los brazos eran vestigiales en abelisáuridos, porque las fibras nerviosas responsables de la transmisión de estímulos nerviosos se redujeron en un grado visto hoy en emúes y kiwis, que también tienen extremidades anteriores vestigiales.[28]
Descubrimiento e investigación

Se lo encontró en la Patagonia de Argentina, en la Provincia del Chubut.[1] El esqueleto del holotipo, MACN-CH 894, y hasta ahora el único conocido fue descubierto en 1984 por una expedición dirigida por el paleontólogo argentino José Fernando Bonaparte.[5] Sus restos pertenecen a sedimentos de la Formación La Colonia. Fue un hallazgo muy interesante porque estos restos proporcionaron a los científicos una pista de cómo era la piel de los dinosaurios. En esta expedición también se recuperó al saurópodo con espinas Amargasaurus.[29] Fue la octava expedición dentro del proyecto denominado "Vertebrados Terrestres de América del Sur del Jurásico y Cretácico", que comenzó en 1976 y que fue patrocinada por la National Geographic Society.[29][5] El esqueleto está bien conservado y articulado, es decir todavía con los huesos conectados entre sí, sólo faltándole los dos tercios posteriores de la cola, la parte inferior de la pierna y pies destruidos por la intemperie.[5] Durante la fosilización, el cráneo y especialmente el hocico fueron aplastados lateralmente, mientras que los premaxillares fueron empujados hacia arriba sobre los huesos nasales. Como resultado, la curvatura hacia arriba de la mandíbula superior es artificialmente exagerada en el holotipo.[11] El esqueleto pertenecía a un individuo adulto, tal como lo indican las suturas fusionadas en la cintura.[14] Se encontró acostado en su lado derecho, mostrando una típica pose de muerte con el cuello doblado hacia atrás sobre el torso.[22] Inusualmente, se conserva con impresiones extensas de la piel.[1] En vista de la importancia de estas impresiones, una segunda expedición fue comenzada para reinvestigar el sitio original de la excavación, conduciendo a la recuperación de varios remiendos adicionales de la piel.[5]
El ejemplar fue colectado en la estancia "Pocho Sastre", cerca de Bajada Moreno, departamento de Telsen, Chubut, Argentina, en sedimentos correspondientes a la sección inferior de la formación La Colonia, Cretácico superior entre el Campaniano y el Maastrichtiano, con una antigüedad de aproximadamente 72 a 68 millones de años. El material holotípico del Carnotaurus sastrei se encuentra depositado en el Museo Argentino de Ciencias Naturales Bernardino Rivadavia, donde también puede apreciarse una réplica de su esqueleto de tamaño natural.[1] Debido a que estaba incrustado en una gran concreción de hematites, un tipo muy duro de roca, la preparación fue complicada y progresó lentamente.[1][16] En 1985, Bonaparte publicó una nota presentando a Carnotaurus sastrei como un nuevo género y especie, y brevemente describen el cráneo y la mandíbula inferior.[1] El nombre genérico deriva del latín carnis "carne" y taurus "toro" se refiere a los cuernos,[30] mientras que el nombre específico sastrei honra a Ángel Sastre, el dueño del rancho donde él encontró el esqueleto.[31] Una descripción completa de todo el esqueleto se realizó en 1990.[5] Después de Abelisaurus, Carnotaurus fue el segundo miembro de la familia Abelisauridae que fue descubierto.[32] Durante años fue, por mucho, el miembro más completo de su familia, y también el terópodo mejor entendido del hemisferio sur.[18][33] No fue hasta el siglo XXI que se describieron abelisáuridos tan bien conservados, incluyendo Aucasaurus, Majungasaurus y Skorpiovenator, permitiendo a los científicos revaluar ciertos aspectos de la anatomía del Carnotaurus.[11] Réplicas se pueden ver en este y otros museos de todo el mundo.[23] Los escultores Stephen y Sylvia Czerkas crearon una escultura de tamaño natural del Carnotaurus que ahora está expuesta en el Museo de Historia Natural del Condado de Los Ángeles. Esta escultura, ordenada por el museo a mediados de la década de 1980, es probablemente la primera restauración de vida de un terópodo que muestra una piel precisa.[22][34]
¿Cretácico inferior o superior?
Originalmente fue datado en mediados del Cretácico, hace cien millones de años, durante el Albiense, en la Formación Gorro Frigio.[1] Ya en los primeros trabajos publicados se hacía la salvedad de que las dataciones estratigráficas de la zona donde se encontraron los restos no eran muy precisas y se exponía la posibilidad que fuese más joven. En 2002 M. C. Lamanna y colaboradores publicaron una datación definitiva que coloca a Carnotaurus a finales del Cretácico superior, en la Formación La Colonia, siendo contemporáneo de otros abelisáuridos con cuernos de la India y de Madagascar.[35]

Clasificación
Carnotaurus es uno de los géneros mejor conocidos dentro de Abelisauridae, una familia de grandes terópodos restringida al antiguo supercontinente Gondwana y algunas partes de lo que es hoy Europa.[36] Los abelisáuridos eran los depredadores dominantes en el Cretácico Tardío de estos lugares, en sustitución de los carcharodontosáuridos y ocupando el nicho ecológico ocupado por los tiranosáuridos en los continentes del norte.[3]
Aunque el cráneo muestra unas proporciones muy distintas de las de Abelisaurus comahuensis,[1] posee los caracteres diagnósticos de Abelisauridae, tales como la ventana infratemporal grande, el cuadrado alargado, el cuadrado yugal fusionado con el cuadrado, el contacto del postorbital sobre la órbita, la ventana preantorbital reducida, con el escamoso posteriormente dirigido hacia atrás y un proceso dorsal estrecho en el maxilar.[17] A partir de las características anteriormente mencionadas, José Fernando Bonaparte, Fernando Novas y Rodolfo Coria[5] sugirieron que ambos, Abelisaurus y Carnotaurus, pertenecen a una nueva familia a la que llamaron Abelisauridae, según la descripción realizada por Bonaparte y Novas en 1985.[37] Se observa fuerte diferencia entre los terópodos de Laurasia y los de Gondwana, especialmente en el cráneo, esqueleto axial y la llamativa reducción de los miembros superiores. Estas diferencias anatómicas han sido interpretadas en consecuencia de la separación geográfica a finales del Cretácico de los supercontinentes .[1][38][37][6] Carnotaurus sirve de base para el nombre de dos subgrupos dentro de Abelisauridae, la subfamilia Carnotaurinae y la tribu Carnotaurini. Los paleontólogos no aceptan universalmente estos grupos. Carnotaurinae se definió para incluir todos los abelisáuridos derivados con la exclusión de Abelisaurus, que se considera un miembro basal en la mayoría de los estudios.[39] Sin embargo, una revisión de 2008 sugirió que Abelisaurus era un miembro derivado en su familia.[11] Se propuso nombrar Carnotaurini al clado formado por Carnotaurus y Aucasaurus;[40] sólo aquellos paleontólogos que consideran Aucasaurus como el pariente más cercano de Carnotaurus utilizan este grupo.[41]
Taxonomía
Con una única especie conocida, Carnotaurus sastrei, fue descubierto en 1985 por el paleontólogo José F. Bonaparte,[1] en la provincia argentina de Chubut en sedimentos de la formación La Colonia. Pertenece a la familia Abelisauridae.[32] Aunque las relaciones dentro de los Abelisauridae se encuentran en debate, a Carnotaurus se lo considera constantemente como uno de los miembros más derivados de la familia en los análisis cladisticos.[11] Su pariente más cercano podría haber sido Aucasaurus[42][40][43] o Majungasaurus;[44][45][46] esta ambigüedad es en gran parte debida a lo incompleto del material del cráneo de Aucasaurus.[12][11] Una revisión reciente sugiere que Carnotaurus no estaba estrechamente relacionado con Aucasaurus o Majungasaurus, y en su lugar se propone a Ilokelesia como su taxón hermano.[11]
En 2008 Canale et al. erigieron el clado Brachyrostra, que incluye a Carnotaurus junto con los otros carnotaurinos menos derivados de América del Sur.[42][36] separado de las formas más basales como Rugops y Kryptops de África, Xenotarsosaurus de Sudamérica y Genusaurus y Tarascosaurus de Europa. Dentro de Brachyrostra se encuentran formas más basales como Ilokelesia, Ekrixinatosaurus y Skorpiovenator pero sin formar parte de la subfamilia Carnotaurinae a la que Carnotaurus pertenece junto a Pycnonemosaurus, Quilmesaurus y Abelisaurus, todos sudamericanos.[36] La otra subfamilia de Abelisauridae es Majungasaurinae, que incluye formas de la India como Indosuchus, Rajasaurus y Rahiolisaurus, de Madagascar, Majungatholus, y europeas como Arcovenator. Dentro de Carnotaurinae su pariente más cercano es Aucasaurus, también de Sudamérica; estos dinosaurios forman la tribu Carnotaurini,[40][36] En 2015 Filippi et al., en la descripción de Viavenator, crearon un nuevo clado, Furileusauria, que incluye a los Brachyrostra más avanzados, colocando a Carnotaurus solo en Carnotaurinae, con Aucasaurus y Abelisaurus en la subfamilia Abelisaurinae, por lo que la tribu Carnotaurini quedaría monotípica y sería redundante.[47]
Filogenia
| Carnotaurinae |
| ||||||||||||||||||||||||||||||
Cladograma de Carnotaurinae, de acuerdo con Canale (2008).[42]
| Abelisauridae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Cladograma de Abelisauridae, de acuerdo con Tortosa et. al. (2013).
Paleobiología

A pesar de ser conocido el hecho de que algunos terópodos presentan estructuras óseas en el cráneo como crestas longitudinales en Megapnosaurus, Dilophosaurus y Cryolophosaurus, o un cuerno nasal como en Ceratosaurus, éste es el único ejemplo descrito de un vertebrado bípedo corredor carnívoro provisto de un par de robustos cuernos frontales. Se ha propuesto que el Carnotaurus experimentaba grandes fuerzas en su cráneo tales como las producidas por impactos de alta velocidad sobre la presa, siendo esta su estrategia de caza. Otra explicación del aparente sobrediseño del cráneo del Carnotaurus, probablemente complementaria más que alternativa, surge de la consideración de fuertes impactos a nivel de los cuernos frontales, los que resultarían de comportamientos de combate con los de su misma especie, ya sea por el territorio, reproducción o liderazgo de grupo.[6] Un estudio más reciente demostró que un Carnotaurus no pudo haber sobrevivido a un embate con sus cuernos. Se ha estudiado que era muy rápido, cubría catorce metros por segundo, alrededor de cincuenta km/h y era de desplazamientos muy ágiles,[48] con una resistencia femoral veintinueve GPa.[6] Se le considera como el terópodo más veloz que haya existido.[21] Acompañando la fortaleza del cráneo están los poderosos músculos del cuello y la rigidez extra proporcionada por el alto número de vértebras sacras. Al igual que Allosaurus, Carnotaurus posee adaptaciones para cazar presas con un tamaño corporal mayor que el suyo. Se ven articulaciones en la mandíbula que le permiten una apertura de la boca excepcional, inserciones de los músculos del cuello que incrementan los movimientos ventrales aumentando la palanca junto con músculos temporales reducidos y pequeños dientes aserrados. En el Cretácico sudamericano la fauna de grandes saurópodos llevó a Carnotaurus a poseer adaptaciones convergentes con los predadores del Jurásico Superior norteamericano, con un hocico reducido, el anillo superior del occipucio reforzado, pero con un arreglo distinto de los huesos. Esto junto con lo pequeño de los dientes hace pensar que usaba la mandíbula superior para embestir.[49]
Cinética del cráneo y mordida
El análisis del diseño de la mandíbula de Carnotaurus por Mazzetta et al. sugiere que el animal era capaz de mordidas rápidas, pero no fuertes.[6][7][50] Estos movimientos de picoteo son más importantes que las fuertes mordidas al capturar pequeñas presas, como lo demuestran los estudios de cocodrilos de hoy en día.[50] Además, estos investigadores observaron un alto grado de flexibilidad del cráneo, especialmente en la mandíbula inferior, algo similar a las serpientes modernas. Esta elasticidad de la mandíbula le habría permitido al Carnotaurus tragar pequeñas presas enteras.[50] Además, la parte anterior de la mandíbula inferior posee una articulación, siendo capaz de moverse hacia arriba y hacia abajo. Cuando se presiona hacia abajo, los dientes se proyectan hacia delante, permitiendo a Carnotaurus atrapar pequeñas presas, y cuando se presiona hacia arriba, los dientes ahora se proyectan hacia atrás, lo que habría obstaculizado que la presa atrapada se escapara.[6] Por otro lado los mismos investigadores también encontraron que el cráneo era capaz de soportar las fuerzas que aparecen cuando se tira de grandes presas.[50] Carnotaurus, por lo tanto, no solo puede haberse alimentado principalmente de relativamente pequeñas presas, sino que también fue capaz de cazar grandes dinosaurios.[50]

A pesar del aspecto grácil del cráneo, este mostraba una resistencia mecánica superlativa. Existe una gran diferencia entre la máxima fuerza que resiste y la generada por una mordida. Esto le da un factor de seguridad de 92, que sugiere que el cráneo de Carnotaurus podría haber estado sobrediseñado.[51] Esto es sensiblemente más alto que el estimado para Allosaurus, que fue de 62,[52][53] calculados usando el método de análisis de elementos finitos. Este se basa en el análisis de elementos finitos de un modelo del cráneo obtenido a partir de un barrido topográfico del cráneo y de la mandíbula. En este sentido los cuernos de Carnotaurus son anchos y planos en dorsal, lo que proporciona una amplia base de contacto. La presencia de cuernos y la fuerte fusión observada entre los huesos frontales y parietales sugieren que este sector del cráneo presenta un diseño para resistir elevadas cargas compresivas. Este modelo de elementos finitos fue utilizado para determinar la fuerza ejercida por la musculatura mandibular. La musculatura fue agrupada en cuatro unidades funcionales: el grupo muscular temporal, el grupo muscular pterigoideo, el músculo aductor mandibular posterior y el músculo intramandibular. La fuerza de mordida de origen muscular máxima estimada en los dientes posteriores es de 3341 N, que resulta relativamente débil al ser comparada con la de otras especies y no habría sido suficiente para triturar el hueso de presas de mediano a gran tamaño.[7] Esto es especialmente cierto para el neurocráneo, en donde la acción muscular minimiza marcadamente las tensiones y deformaciones en respuesta a las fuerzas aplicadas sobre la fila dentaria. El gran rango de movimientos y el apropiado diseño de sus dientes, conjuntamente con la fuerza originada por su musculatura del cráneo y del cuello, le habrían permitido a Carnotaurus cortar y desgarrar grandes trozos de alimento que tragaría sin masticar.[7]
La interpretación de cazador de presas pequeñas fue cuestionada por François Therrien et al., quienes encontraron que la fuerza de mordida de Carnotaurus era dos veces mayor que la del cocodrilo americano, que puede tener la mordida más fuerte de un tetrápodo viviente. Estos investigadores también observaron analogías con los modernos dragones de Komodo. La resistencia a la flexión de la mandíbula inferior disminuye hacia la punta de forma lineal, lo que indica que las mordidas no eran adecuadas para la alta precisión necesaria para la captura de presas pequeñas, pero podía ser usada causando heridas para debilitar a una gran presa. Como consecuencia de ello, de acuerdo con este estudio, Carnotaurus debía de haber hecho presa principalmente en animales grandes, posiblemente por emboscada.[54] Cerroni y sus colegas, en 2020, argumentaron que la flexibilidad estaba restringida a la mandíbula inferior, mientras que el techo del cráneo engrosado y la osificación de varias articulaciones craneales sugieren que el cráneo tenía poca o ninguna cinesis.[15]
Por otro lado, Robert Bakker encontró que el Carnotaurus se alimentaba principalmente de presas muy grandes, especialmente de saurópodos. Como se señala anteriormente, varias adaptaciones del cráneo-hocico corto, los relativamente pequeños dientes y la fuerte parte de atrás del cráneo evolucionaron independientemente también en Allosaurus. Estas características sugieren que la mandíbula superior se utilizó como un hacha serrada para infligir heridas a grandes saurópodos que se habrían debilitado por los ataques repetidos.[53]
Función de los cuernos
Por lo que se sabe hasta hoy, Carnotaurus es el único carnívoro conocido con un par de cuernos en el hueso frontal.[50] El uso de estos cuernos no está del todo claro, existiendo varias interpretaciones que han girado en torno a su uso en luchas con sus congéneres, para exhibirse o para matar presas.

Gregory S. Paul propuso que los cuernos se usaron en batallas de empujones y que la órbita ocular se ha reducido para minimizar la posibilidad de daño a los ojos mientras ocurría la lucha.[55] Gerardo Mazzetta et al. sugirieron que Carnotaurus utiliza los cuernos de una manera similar a carneros actuales. Calcularon que la musculatura del cuello era lo suficientemente fuerte como para absorber la fuerza de dos individuos que chocaran con la cabeza frontalmente a una velocidad de 5,7 m/s cada uno.[6] Fernando Novas interpretó varias características esqueléticas como adaptaciones para la entrega de los golpes con la cabeza. Sugirió que el cráneo pequeño podría haber realizado movimientos de la cabeza más rápidos, reduciendo el momento de inercia, mientras que el cuello musculoso habría permitido fuertes golpes con la cabeza. También señaló una rigidez mejorada y la fuerza de la columna vertebral puede haber evolucionado para soportar choques realizados por la cabeza y el cuello.[12]
Otros estudios sugieren que Carnotaurus no podía dar golpes rápidos con la cabeza, pero sí empujarse lentamente el uno contra el otro con los lados superiores de sus cráneos.[50][56] Por lo tanto, los cuernos pueden haber sido un dispositivo para disipar las fuerzas de compresión y no dañar el cerebro.[50] Esto es apoyado por los lados superiores aplanados de los cuernos, los huesos fusionados fuertemente de la parte superior del cráneo y la incapacidad del cráneo para soportar golpes rápidos en la cabeza.[50]
Mazzetta et al. proponen que los cuernos también podrían haber sido usados para herir o matar a pequeñas presas. Aunque los núcleos óseos son romos, pueden haber tenido una cubierta queratinosa de forma similar a los modernos bóvidos. Sin embargo, esta teoría pierde credibilidad, ya que sería el único ejemplo informado de cuernos siendo utilizados como armas de caza en animales.[6]
Locomoción
Mazzetta et al. presumen que Carnotaurus era un corredor veloz; como argumento muestran que el hueso del muslo estaba adaptado para soportar altos momentos de flexión mientras corría. La capacidad de la pierna de un animal para soportar estas fuerzas limita su velocidad máxima. Las adaptaciones que se ven en Carnotaurus son superiores a las de un ser humano, aunque no tan buenas como las de un avestruz, ambos seres bipedos.[6][48] Los científicos calculan que Carnotaurus tenía una velocidad máxima de hasta 48-56 kilómetros por hora.[57]

En los dinosaurios, el músculo locomotor más importante se encuentra en la cola. Este músculo, llamado caudofemoralis, se une al cuarto trocánter, una cresta prominente en el fémur, hueso del muslo, y tira del hueso del muslo hacia atrás cuando se contrae. Scott Persons y Philip J. Currie señalan que en las vértebras de la cola de Carnotaurus las costillas caudales no sobresalen horizontalmente, en forma de "T", sino en ángulo contra el eje vertical de las vértebras, formando una "V". Esto habría proporcionado un espacio adicional para un músculo caudofemoralis mayor que en cualquier otro terópodo. La masa muscular se calculó entre 111 y 137 kilogramos por pierna. Por lo tanto, Carnotaurus podría haber sido uno de los más rápidos y grandes terópodos conocidos.[21] Mientras que el músculo caudofemoralis se amplía, los músculos epiaxiales situados por encima de las costillas caudales habrían sido proporcionalmente menores. Estos músculos, llamados longísimo y espinoso, eran responsables del movimiento de la cola y la estabilidad. Para mantener la estabilidad de la cola a pesar de la reducción de estos músculos, las costillas caudales llevan hacia delante la proyección de procesos entrelazados con las vértebras entre sí y con la pelvis, aumentando la rigidez de la cola. Como consecuencia de ello, la capacidad de tomar curvas cerradas habría disminuido, debido a que la cadera y la cola tuvieron que moverse simultáneamente, a diferencia de otros terópodos.[21]
Cerebros y sentidos
Cerroni y Paulina-Carabajal, en 2019, utilizaron una tomografía computarizada para estudiar la cavidad endocraneal que contenía el cerebro. El volumen de la cavidad endocraneal era de 168,8 cm³, aunque el cerebro sólo habría llenado una fracción de este espacio. Los autores utilizaron dos estimaciones diferentes del tamaño del cerebro, asumiendo un tamaño del cerebro del 50% y el 37% de la cavidad endocraneal, respectivamente. Esto da como resultado un cociente de encefalización de reptiles, una medida de inteligencia, mayor que el del relacionado Majungasaurus, pero menor que en los tiranosáuridos. La glándula pineal, que produce hormonas, podría haber sido más pequeña que en otros abelisáuridos, como indica una expansión dural baja, un espacio en la parte superior del prosencéfalo en el que se cree que se encuentra la glándula pineal.[9]
Los bulbos olfativos, que albergaban el sentido del olfato, eran grandes, mientras que los lóbulos ópticos, responsables de la vista, eran relativamente pequeños. Esto indica que el sentido del olfato podría haberse desarrollado mejor que el sentido de la vista, mientras que ocurre lo contrario en las aves modernas. La parte frontal de los tractos olfatorios y los bulbos se curvó hacia abajo, una característica que solo comparte Indosaurus; en otros abelisáuridos, estas estructuras estaban orientadas horizontalmente. Según la hipótesis de Cerroni y Paulina-Carabajal, esta curvatura hacia abajo, junto con el gran tamaño de los bulbos, podría indicar que Carnotaurus dependía más del sentido del olfato que otros abelisáuridos. El flóculo, un lóbulo del cerebro que se cree que se correlaciona con la estabilización de la mirada, coordinación entre los ojos y el cuerpo, era grande en Carnotaurus y otros abelisáuridos sudamericanos. Esto podría indicar que estas formas utilizan con frecuencia movimientos rápidos de la cabeza y el cuerpo. La audición podría haber estado poco desarrollada en Carnotaurus y otros abelisáuridos, como indica la lagena corta del oído interno. Se estimó que el rango de audición era inferior a 3 kHz.[9]
Paleoecología

La formación La Colonia se interpreta como depósitos de un ambiente de estuarios, con costa baja e intercambio de agua dulce de continente con la de las mareas marinas, donde se hallan moluscos bivalvos similares a ostras. Esta porción de mar es conocida como el mar de Káwas, también llamada Transgresión de Schiller.[58] El análisis sedimentológico sugiere que la deposición habría sido sobre todo en ambientes de baja energía restringidos, como llanuras fangosas, pantanos y estanques cortados por canales serpenteantes, probablemente la zona central de energía mixta dentro de un estuario. Esta interpretación es coherente con el hábitat inferido de los reptiles recuperados, así como con foraminíferos asociados y con el origen probable de gastrolitos encontrados y asociados con los plesiosaurios.[59] La mayoría de los vertebrados recogidos incluyen peces, tortugas, crocodilomorfos, plesiosauros, dinosaurios, serpientes y mamíferos.[60][59] Entre los dinosaurios, además de Carnotaurus, se encontraron fragmentos de los metatarsianos de un terópodo, restos de otro abelisáurido distinto a Carnotaurus, una vértebra de un titanosauriano, partes de los miembros de un hadrosauroide y osteodermos de un anquilosauriano.[59] Entre las serpientes se pueden hallar de las familias Boidae y Madtsoidae, como Alamitophis argentinus.[60] Las tortugas están representadas por al menos cinco taxones de Chelidae, miembro de Pleurodira, entre ellos tres ejemplares de Yaminuechelys gasparinii y dieciséis de la Meiolaniidae, perteneciente a Cryptodira, Patagoniaemys gasparinae.[61][59] Dentro de los plesiosaurios encontrados allí se incluye a Sulcusuchus erraini,[62] un reptil marino longirrostro de la familia Polycotylidae y elasmosauridos, inclusive dos con gastrolitos asociados.[61][63][59] Entre los mamíferos aparece Reigitherium bunodontum, el cual fue considerado como el primer registro de un docodonte en Sudamérica[58] y actualmente es de ubicación incierta, y Argentodites coloniensis, un posible Cimolodonta.[64] Se encontraron restos de peces dipnoos, otros peces de agua dulce y anuros.[59] A excepción de los plesiosaurios, todos los reptiles son taxones terrestres o de agua dulce. Sin embargo, el análisis de los elasmosáuridos indica que son ejemplares adultos de pequeño tamaño, lo que podría estar relacionado con formas que vivían en ambientes acuáticos restringidos. Asimismo, el policotílido posee un profundo rostral y ranuras inferiores como una vascularización e inervación visible, que es consistente con la presencia de algunas estructuras sensoriales especiales similares a los conocidos en algunos cetáceos modernos que habitan en ríos y estuarios.[59]
Entre los dinosaurios, así como Carnotaurus fue datado erróneamente en un principio, también lo podría haber sido Chubutisaurus, que pertenecería a la misma formación.[65] Igualmente en la Patagonia y siendo contemporáneos, habitaron los titanosaurianos Antarctosaurus, Puertasaurus, Neuquensaurus, Epachthosaurus, Pellegrinisaurus, Rocasaurus, Aeolosaurus, Bonatitan, Laplatasaurus y Saltasaurus, los euiguanodóntidos Talenkauen y Gasparinisaura, el hadrosáurido Secernosaurus y el posible ceratopsiano Notoceratops, todos ellos herbívoros y posibles presas. También compartió el territorio con el megarraptórido de gran tamaño Orkoraptor, los abelisáuridos Quilmesaurus y Aucasaurus y probablemente con el dromeosáurido de gran tamaño Austroraptor. La paleoflora era conocida por sus componentes acuáticas, Paleoazolla[66] y Regnellidium. Sin embargo, los descubrimientos paleobotánicos recientes han revelado la presencia de una variedad más diversa de las plantas asociadas a estos cuerpos acuáticos, incluyendo pteridofitas, gimnospermas y varias angiospermas.[67][68] Entre estas últimas aparecen los fósiles de frutos de una Nelumbonaceae, siendo la más antigua para América del Sur de este grupo.[69]
En la cultura popular
Hasta mediados de los años 1990 la aparición del Carnotaurus en los medios había sido ocasional. Una de sus primeras apariciones importantes fue en 1995, en la novela El mundo perdido, la secuela de la exitosa novela Parque Jurásico de Michael Crichton. En ella aparecía con la habilidad de cambiar de color como un camaleón o una sepia,[70] lo que permitía al animal camuflarse con su entorno. Esto no es más que una licencia literaria del escritor, y desde luego no está basada en evidencia de campo, a pesar de ser omitido en la versión cinematográfica dirigida por Steven Spielberg en 1997.
En el cine
Aparece en el film de Disney, Dinosaurio, donde una pareja de Carnotaurus actúan como principales antagonistas acosando a una manada de dinosaurios herbívoros. En la película aparece mucho más grande de lo que fue, con proporciones parecidas a las de un Tiranosaurio, cuando en realidad el Carnotauro tenía un tamaño similar al Iguanodonte.[71]
Galería
 Montura del esqueleto de un carnotauro.
Montura del esqueleto de un carnotauro. Cráneo de un carnotauro desde un punto de vista lateral.
Cráneo de un carnotauro desde un punto de vista lateral. Montura anacrónica de un carnotauro y un amargasaurio.
Montura anacrónica de un carnotauro y un amargasaurio.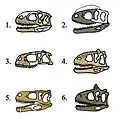 Comparativa de cráneos de abelisáuridos: 1)Indefinido 2)Rugops 3)Abelisaurus 4)Majungasaurus 5)Aucasaurus 6)Carnotauro.
Comparativa de cráneos de abelisáuridos: 1)Indefinido 2)Rugops 3)Abelisaurus 4)Majungasaurus 5)Aucasaurus 6)Carnotauro. Dibujos de dos carnotauros.
Dibujos de dos carnotauros.
Véase también
 Portal:Dinosaurios. Contenido relacionado con Dinosaurios.
Portal:Dinosaurios. Contenido relacionado con Dinosaurios.- El mundo perdido (novela)
- Dinosaurio (película)
- Abelisauridae
- Abelisaurus
- José Fernando Bonaparte
- Anexo:Géneros válidos de dinosaurios
Referencias
- Bonaparte, J. F. (1985). "A horned Cretaceous carnosaur from Patagonia." National Geographic Research, 1: 149-151.
- Sereno, P. C.; Wilson J. A.; Conrad J. L. (2004).New dinosaurs link southern landmasses in the Mid-Cretaceous; Proc. R. Soc. Lond. B
- Candeiro, C. R. d. A.; Martinelli, A. G. (2005) "Abelisauroidea and carchardontosauridae (theropoda, dinosauria) in the cretaceous of south america. Paleogeographical and geocronological implications". Uberlândia (Sociedade de Naturaleza) 17 (33): 5–19.
- Juárez Valieri, R. D.; Porfiri, J. D.; Calvo, J. O. (2010). "New information on Ekrixinatosaurus novasi Calvo et al 2004, a giant and massively-constructed Abelisauroid from the Middle Cretaceousof Patagonia".
- Bonaparte, J. F.; Novas, F. E.; Coria, R. A. (1990). Carnotaurus sastrei Bonaparte, the horned, lightly built carnotaur from the middle Cretaceous of Patagonia. Contributions Science Natural History Museum of Los Angeles County, 416, 1-42.
- Mazzetta, G. V.; Fariña, R. Q.; Vizcaíno, S. F. (1998).On the palaeobiology of the south american horned theropod Carnotaurus sastrei Bonaparte.Gaia. Aspects of Theropod Paleobiology. Ed. B. P. Pérez Moreno, T. Holtz, Jr., J. L. Sanz & J. Moratalla. 15,185-192.
- Mazzetta, G. V.; Christiansen, P.; Farin, R. (2004).Giants and Bizarres:Body Size of Some Southern South American Cretaceous Dinosaurs; Historical Biology, 1–13
- Paul, Gregory S. (2010). The Princeton Field Guide to Dinosaurs (1st edición). Princeton University Press. ISBN 9780691137209. Parámetro desconocido
|title-link=ignorado (ayuda) - Cerroni, Mauricio A.; Paulina-Carabajal, Ariana (2019). «Novel information on the endocranial morphology of the abelisaurid theropod Carnotaurus sastrei». Comptes Rendus Palevol 18 (8): 985-995. doi:10.1016/j.crpv.2019.09.005. Parámetro desconocido
|doi-access=ignorado (ayuda) - Calvo, J. O.; Rubilar-Rogers, D.; Moreno, K. (2004). "A new Abelisauridae (Dinosauria: Theropoda) from northwest Patagonia". Archivado el 9 de marzo de 2012 en Wayback Machine. Ameghiniana 41 (4): 555–563.
- Carrano, M. T.; Sampson, S. D. (January 2008). "The Phylogeny of Ceratosauria (Dinosauria: Theropoda)". Journal of Systematic Palaeontology 6 (2): 183–236.
- Novas, F. E. (2009). The age of dinosaurs in South America. Bloomington: Indiana University Press. ISBN 978-0-253-35289-7.
- Sampson, S. D.; Witmer, L. M. (2007). "Craniofacial Anatomy of Majungasaurus crenatissimus(Theropoda: Abelisauridae) From the Late Cretaceous of Madagascar". Journal of Vertebrate Paleontology 27 (sp8): 95–96. doi:10.1671/0272-
- Carabajal, P. A. (2011). "The braincase anatomy of Carnotaurus sastrei (Theropoda: Abelisauridae) from the Upper Cretaceous of Patagonia". Journal of Vertebrate Paleontology 31 (2): 379. doi:10.1080/02724634.2011.550354.
- Cerroni, M. A.; Canale, J. I.; Novas, F. E. (2020). «The skull of Carnotaurus sastrei Bonaparte 1985 revisited: insights from craniofacial bones, palate and lower jaw». Historical Biology: 1-42. doi:10.1080/08912963.2020.1802445.
- Paul, G. S. (1988). Predatory Dinosaurs of the World. pp. 284–285. ISBN 0-671-61946-2.
- Novas, F. E. (1989); Los dinosaurios carnívoros de la Argentina. Tesis Univ. Nac. La Plata, Fac. Ciencias Naturales, 510 pp.
- Bonaparte, J. F. (1996). Cretaceous tetrapods of Argentina. Münchner Geowissenchaftliche Abhandlungen, (A), 30, 73-130.
- Hartman, S. (2012)."Carnotaurus – delving into self-parody?". Retrieved 7.12.2012.
- Méndez, A. (2014). "The cervical vertebrae of the Late Cretaceous abelisaurid dinosaur Carnotaurus sastrei". Acta Palaeontologica Polonica. doi:10.4202/app.2011.0129.
- Persons, W. S.; Currie, P. J. (2011). Farke, Andrew Allen, ed. "Dinosaur Speed Demon: The caudal musculature of Carnotaurus sastrei and implications for the evolution of South American abelisaurids". PLoS ONE 6 (10): e25763. doi:10.1371/journal.pone.0025763. PMC 3197156. PMID 22043292. edit
- Czerkas, S. A.; Czerkas, S. J. (1997). "The Integument and Life Restoration of Carnotaurus". In Wolberg, D. I.; Stump, E.; Rosenberg, G. D. Dinofest International. Academy of Natural Sciences, Philadelphia. pp. 155–158.
- Glut, D. F. (2003). "Carnotaurus". Dinosaurs: The Encyclopedia. 3rd Supplement. Jefferson, North Carolina: McFarland & Company, Inc. pp. 274–276.ISBN 0-7864-1166-X.
- Ruiz, J.; Torices, A.; Serrano, H.; Lopez, V. (2011). The hand structure of Carnotaurus sastrei (Theropoda, Abelisauridae): implications for hand diversity and evolution in abelisaurids: Palaeontology, v. 54, part 6, p. 1271-1277.
- Agnolin, F. L.; Chiarelli, P. (6/2010). "The position of the claws in Noasauridae (Dinosauria: Abelisauroidea) and its implications for abelisauroid manus evolution". Paläontologische Zeitschrift 84 (2): 293–300.
- Middleton, K.; Gatesy, S. (2000). Theropod forelimb design and evolution; Zoological Journal of the Linnean Society, 128: 149–187.
- http://dml.cmnh.org/2001Apr/msg00486.html Archivado el 22 de julio de 2020 en Wayback Machine. Post de Archives of the DINOSAUR Mailing List sobre la posición de los brazos del Carnotaurus
- Senter, P. (2010). "Vestigial skeletal structures in dinosaurs". Journal of Zoology 280: 60–71. doi:10.1111/j.1469-7998.2009.00640.x.
- Salgado, Leonardo; Bonaparte, José F. (1991). "Un nuevo sauropodo Dicraeosauridae, Amargasaurus cazaui gen. et sp. nov., de la Formacion La Amarga, Neocomiano de la Provincia del Neuquén, Argentina". Ameghiniana (in Spanish). 28 (3–4): 334.
- Creisler, Ben (7 de julio de 2003). «Dinosauria Translation and Pronunciation Guide». Dinosauria On-Line. Archivado desde el original el 10 de julio de 2011. Consultado el 27 de diciembre de 2012.
- Headden, Jaime A. (19 de septiembre de 2006). «Re: Carnotaurus sastrei etymology». Dinosaur Mailing List. Archivado desde el original el 22 de febrero de 2012. Consultado el 10 de octubre de 2017.
- Bonaparte J. F. (1991). "The Gondwanian theropod families Abelisauridae and Noasauridae." Historical Biology. An International Journal of Paleobiology, 5: 1-25.
- Glut, Donald F. (1997). "Carnotaurus". Dinosaurs, the encyclopedia. Jefferson, North Carolina: McFarland & Company, Inc. Publishers. pp. 256–259. ISBN 978-0-375-82419-7.
- Glut, Donald F. (2000). "Carnotaurus". Dinosaurs: The Encyclopedia. 1st Supplement. Jefferson, North Carolina: McFarland & Company, Inc. pp. 165–167. ISBN 0-7864-0591-0.
- Lamanna, M. C.; Martínez, R. B.; Smith, J. B. (2002). A definitive abelisaurid theropod dinosaur from the early Late Cretaceous of Patagonia. J. Vert. Paleontol. 22, 58–69.
- Tortosa, T.; Buffetaut, E.; Vialle, N.; Dutour, Y.; Turini, E.; Cheylan, G. (2013). «A new abelisaurid dinosaur from the Late Cretaceous of southern France: Palaeobiogeographical implications». Annales de Paléontologie (In press). doi:10.1016/j.annpal.2013.10.003. Consultado el 13 de diciembre de 2013.
- Bonaparte, J. F.; Novas, F. E. (1985). Abelisaurus comahuensis, n. g., n. sp., Carnosauria del Cretácico tardío de Patagonia. Ameghiniana, 21 (2-4), 259-261.
- Bonaparte, J. F. (1986). History of the Terrestrial Cretaceous Vertebrates of Gondwana. In: Fourth Congress, Argentina Palaeo. Biostrat. Mendoza, 2, 63-95.
- Sereno, Paul (2005). «Carnotaurinae». Taxon Search. Archivado desde el original el 16 de mayo de 2012. Consultado el 29 de diciembre de 2012.
- Coria, R. A.; Chiappe, L. M.; Dingus, L. (2002). A new close relative of Carnotaurus sastrei Bonaparte 1985 (Theropoda: Abelisauridae) from the Late Cretaceous of Patagonia. Archivado el 7 de octubre de 2011 en Wayback Machine. Journal of Vertebrate Paleontology, 22, 2, 460-465.
- Sereno, Paul (2005). «Carnotaurini». Taxon Search. Archivado desde el original el 16 de mayo de 2012. Consultado el 29 de diciembre de 2012.
- Canale, J.I.; Scanferla, C.A.; Agnolin, F.; Novas, F.E. (2008). "New carnivorous dinosaur from the Late Cretaceous of NW Patagonia and the evolution of abelisaurid theropods." Naturwissenschaften. doi: 10.1007/s00114-008-0487-4.
- Ezcurra, Martín D.; Agnolin, Federico L.; Novas, Fernando E. (2010). «An abelisauroid dinosaur with a non-atrophied manus from the Late Cretaceous Pari Aike Formation of southern Patagonia». Zootaxa 2450: 14.
- Sereno, Paul C.; Wilson, Jeffrey A.; Conrad, Jack L. (7 de julio de 2004). «New dinosaurs link southern landmasses in the Mid-Cretaceous». Proceedings of the Royal Society B: Biological Sciences 271 (1546): 1325-1330. PMC 1691741. PMID 15306329. doi:10.1098/rspb.2004.2692.
- Tykoski, Ronald B.; Rowe, Timothy (2004). «Ceratosauria». En Weishampel, David B.; Dodson, Peter; Osmólska, Halszka, eds. The Dinosauria (Second edición). Berkeley: University of California Press. pp. 65. ISBN 0-520-24209-2.
- Wilson, Jeffrey A.; Sereno, Paul C.; Srivastava, Suresh; Bhatt, Devendra K.; Khosla, Ashu; Sahni, Ashok (2003). «A new abelisaurid (Dinosauria, Theropoda) from the Lameta Formation (Cretaceous, Maastrichtian) of India». Contributions from the Museum of Paleontology (Museum of Paleontology, The University of Michigan) 31 (1): 25.
- Leonardo S. Filippi, Ariel H. Méndez, Rubén D. Juárez Valieri and Alberto C. Garrido (2016). «A new brachyrostran with hypertrophied axial structures reveals an unexpected radiation of latest Cretaceous abelisaurids». Cretaceous Research 61: 209-219. doi:10.1016/j.cretres.2015.12.018.
- Mazzetta, G. V. y Farina, R. A. (1999). Estimación de la capacidad atlética de Amargasaurus cazaui (Salgado y Bonaparte, 1991) y Carnotaurus sastrei (Bonaparte, 1985) (Saurischia, Sauropoda-Theropoda). In: XIV Jornadas Argentinas de paleontología de vertebrados, Ameghiniana, 36, 1, 105-106.
- Bakker, R. T. (1998). Brontosaur killers: Late Jurassic Allosaurids are sabre-tooht cat analogues; Gaia Nº15; Lisboa; pp145-158 (ISSN: 0871 5424)
- Mazzetta, Gerardo V.; Cisilino, Adrián P.; Blanco, R. Ernesto; Calvo, Néstor (2009). "Cranial mechanics and functional interpretation of the horned carnivorous dinosaur Carnotaurus sastrei". Journal of Vertebrate Paleontology 29 (3): 822–830. doi:10.1671/039.029.0313.
- Mazzetta, G. V.; Cisilino, A. P.; Blanco, E. (2004). Distribución de tensiones durante la mordida en la mandíbula de Carnotaurus sastrei Bonaparte, 1985 (Theropoda: Abelisauridae). Ameghiniana, 41, 4, 605-617.
- Rayfield, E. J.; Norman, D. B.; Horner, C. C.; Horner, J. R.; Smith, P. May; Thomason, J. J.; Upchurch, P. (2001). «Cranial design and function in a large theropod dinosaur». Nature 409: 1033-1037. doi:10.1038/35059070.
- Bakker, Robert T. (1999). «Brontosaur killers: Late Jurassic allosaurids as sabre-tooth cat analogues» (pdf). Gaia 15: 145-158. ISSN 0871-5424. Archivado desde el original el 31 de octubre de 2007.
- Therrien, François; Henderson, Donald; Ruff, Christopher (2005). "Bite Me – Biomechanical Models of Theropod Mandibles and Implications for Feeding Behavior". In Carpenter, Kenneth. The carnivorous dinosaurs. Indiana University Press. pp. 179–198, 228. ISBN 0-253-34539-1.
- Paul, Gregory S. (1988). Predatory Dinosaurs of the World. pp. 284–285. ISBN 0-671-61946-2.
- Chure, Daniel J. (1998). "On the orbit of theropod dinosaurs". Gaia 15: 233–240.
- «Predatory dinosaur was fearsomely fast». CBC News. 21 de octubre de 2011. Consultado el 22 de abril de 2017.
- Pascual R.; Goin F. J.; González P.; Ardolino A.; Puerta P. F. (2000). A highly derived docodont from the Patagonian Late Cretaceous: evolutionary implications for Gondwanan mammals. Archivado el 14 de abril de 2012 en Wayback Machine.; Geodiversitas 22 (3): 395-414.
- Gasparini, Z.; Sterlib, J.; Parrasb, A.; O'Gormana, J. P.; Salgado, L.; Varelad, J.; Polb, D. (2014) Cretaceous reptilian biota of the La Colonia Formation, central Patagonia, Argentina: Occurrences, preservation and paleoenvironments; Cretaceous Research; Volume 54, May 2015, Pages 154–168.doi:10.1016/j.cretres.2014.11.010
- Albino A. M. (2000). New record of snakes from the Cretaceous of Patagonia (Argentina). Geodiversitas 22 (2): 247-253.
- Gasparini, Z.; de la Fuente, M. (2000.) Tortugas y plesiosauros de la Formación La Colonia (Cretácico Superior) de Patagonia, Argentina.(res.); Revista Española de Paleontología, 15, 23-35.
- Gasparini, Z.; y Spalletti. L. (1990). Un nuevo cocodrilo en depósitos mareales maastrichtianos de la Patagonia noroccidental. Ameghiniana 27: 141-150.
- Gasparini Z.; Fernández M.; de la Fuente M.; Salgado L. (2007) Reptiles marinos jurásicos y cretácicos de la Patagonia argentina: su aporte al conocimiento de la herpetofauna mesozoica. Asociación Paleontológica Argentina.; Publicación Especial Ameghiniana 50º aniversario: 125-136. Buenos Aires, 11 ISSN 0328-347X
- Kielan−Jaworowska, Z.; Ortiz−Jaureguizar, E.; Vieytes, C.; Pascual, R.; Goín, F.J. (2007). First?cimolodontan multi−tuberculate mammal from South America. Acta Palaeontologica Polonica 52 (2): 257–262.
- http://dml.cmnh.org/2003Sep/msg00120.html Archivado el 5 de diciembre de 2008 en Wayback Machine. Entrada de Archives of the DINOSAUR Mailing List sobre el posible cambio de edad de Chubutisaurus
- Archangelsky, A.;, Phipps, C. J.; Taylor, T. N.; Edith, L. (1999). Paleoazolla, a new heterosporous fern from the Upper Cretaceous of Argentina (enlace roto disponible en Internet Archive; véase el historial, la primera versión y la última).; American Journal of Botany.; 86:1200-1206.
- Gandolfo M. A.,Ruben; N. (2003). Flora of the La Colonia Formation (Maastrichtian, Late Cretaceus), Chubut, Patagonia, Argentina
- Prámparo, M. B.; Quattrocchio, M.; Gandolfo, M. A.; Zamaloa M. d. C.; Romero, E. (2007). Historia evolutiva de las angiospermas (Cretácico-Paleógeno) en Argentina a través de los registros paleoflorísticos; Asociación Paleontológica Argentina. Publicación Especial 11; Ameghiniana 50º aniversario: 157-172. Buenos Aires; ISSN 0328-347X
- Gandolfo, M. A.; Cuneo R. N. (2005). Fossil Nelumbonaceae from the La Colonia Formation (Campanian–Maastrichtian, Upper Cretaceous), Chubut, Patagonia, Argentina; Review of Palaeobotany and Palynology, Volumen 133, Issues 3-4, Páginas 169-178.
- Crichton, Michael (9/1995). The Lost World. Ballantine Books. p. 416 pp. ISBN 0-345-40288-X. (requiere registro).
- Eric Leighton & Ralph Zondag (Director). (19-5-2000) Dinosaur (película). USA: Walt Disney Pictures.
Enlaces externos
 Wikimedia Commons alberga una categoría multimedia sobre Carnotaurus sastrei.
Wikimedia Commons alberga una categoría multimedia sobre Carnotaurus sastrei. Wikispecies tiene un artículo sobre Carnotaurus sastrei.
Wikispecies tiene un artículo sobre Carnotaurus sastrei.- MEF - Carnotaurus sastrei
- Dinosauricon
- Ficha e imagen de Carnotaurus en Dinosaurier-web Archivado el 4 de febrero de 2009 en Wayback Machine. (en alemán)
- Significado y pronunciación del nombre (en inglés)
- Carnotaurus en The Theropod Database (en inglés)
| Control de autoridades |
|
|---|
 Datos: Q18510948
Datos: Q18510948- Multimedia: Carnotaurus / Q18510948
- Especies: Carnotaurus