Eje hipotalámico-hipofisario-adrenal
El eje hipotálamo-hipofisario-suprarrenal (eje HHS) (en inglés Hypothalamic-Pituitary-Adrenal Axis HPA, HTPA) es un conjunto complejo de influencias directas e interacciones de retroalimentación entre tres componentes: el hipotálamo, la hipófisis (una estructura en forma de guisante ubicada debajo del tálamo) y en las suprarrenales (órganos pequeños y cónicos en la parte superior de los riñones).
.svg.png.webp)
.svg.png.webp)
Estos órganos y sus interacciones constituyen el eje HPA, un importante sistema neuroendocrino[1] que controla las reacciones al estrés y regula muchos procesos corporales, incluida la digestión, el sistema inmunitario, el estado de ánimo y las emociones, la sexualidad y el almacenamiento y gasto de energía. Es el mecanismo común de interacciones entre glándulas, hormonas y partes del mesencéfalo que median el síndrome de adaptación general (GAS).[2] Si bien las hormonas esteroides se producen principalmente en vertebrados, el papel fisiológico del eje HPA y los corticosteroides en la respuesta al estrés es tan fundamental que también se pueden encontrar sistemas análogos en invertebrados y organismos monocelulares.
El eje HPA, el eje hipotálamo-pituitario-gonadal (HPG), el eje hipotalámico-pituitario-tiroideo (HPT) y el sistema hipotalámico-neurohipofisario son los cuatro sistemas neuroendocrinos principales a través de los cuales el hipotálamo y la hipófisis dirigen la función neuroendocrina.
Morfología
| Fisiología del sistema endocrino | ||
|---|---|---|
| Sistemas regulatorios | ||
| Campos | ||
| Metabolismo | ||
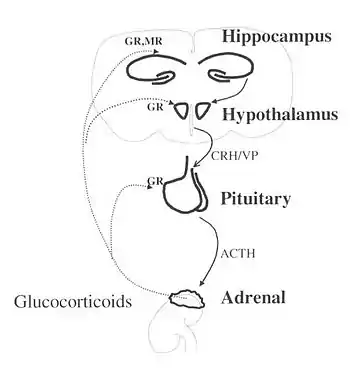
Los elementos clave del eje HPA son:
- El núcleo paraventricular del hipotálamo, que contiene neuronas neuroendocrinas que sintetizan y secretan vasopresina y hormona liberadora de corticotropina (CRH). Estos dos péptidos regulan:
- El lóbulo anterior de la hipófisis. En particular, la CRH y la vasopresina estimulan la secreción de la hormona adrenocorticotrópica (ACTH), antes conocida como corticotropina. ACTH a su vez actúa sobre:
- La corteza suprarrenal, que produce hormonas glucocorticoides (principalmente cortisol en humanos) en respuesta a la estimulación por ACTH. Los glucocorticoides, a su vez, actúan sobre el hipotálamo y la hipófisis (para suprimir la producción de CRH y ACTH) en un ciclo de retroalimentación negativa.
La CRH y la vasopresina se liberan de las terminaciones nerviosas neurosecretoras en la eminencia media. La CRH se transporta a la hipófisis anterior a través del sistema de vasos sanguíneos portales del tallo hipofisario y la vasopresina se transporta por transporte axonal a la hipófisis posterior. Allí, la CRH y la vasopresina actúan sinérgicamente para estimular la secreción de ACTH almacenada de las células corticotropas. La ACTH es transportada por la sangre a la corteza suprarrenal de la glándula suprarrenal, donde estimula rápidamente la biosíntesis de corticosteroides como el cortisol a partir del colesterol. El cortisol es una importante hormona del estrés y tiene efectos en muchos tejidos del cuerpo, incluido el cerebro. En el cerebro, el cortisol actúa sobre dos tipos de receptores: los receptores de mineralocorticoides y los receptores de glucocorticoides, y estos se expresan en muchos tipos diferentes de neuronas. Un objetivo importante de los glucocorticoides es el hipotálamo, que es un importante centro de control del eje HPA.
Se puede considerar que la vasopresina es una "hormona de conservación de agua" y también se la conoce como "hormona antidiurética". Se libera cuando el cuerpo está deshidratado y tiene potentes efectos de conservación de agua en el riñón. También es un potente vasoconstrictor.[3]
Algunos de los bucles de retroalimentación son importantes para la función del eje HPA:
- El cortisol producido en la corteza suprarrenal se retroalimentará negativamente para inhibir tanto el hipotálamo como la hipófisis. Esto reduce la secreción de CRH y vasopresina, y también reduce directamente la división de proopiomelanocortina (POMC) en ACTH y β-endorfinas.
- La epinefrina y la norepinefrina (E/NE) son producidas por la médula suprarrenal a través de la estimulación simpática y los efectos locales del cortisol (enzimas de regulación positiva para producir E/NE). E/NE retroalimentará positivamente a la hipófisis y aumentará la descomposición de las POMC en ACTH y β-endorfinas.
Función
La liberación de la hormona liberadora de corticotropina (CRH) del hipotálamo está influenciada por el estrés, la actividad física, la enfermedad, los niveles de cortisol en la sangre y el ciclo de sueño/vigilia (ritmo circadiano). En individuos sanos, el cortisol aumenta rápidamente después de despertarse, alcanzando un máximo en 30 a 45 minutos. Luego cae gradualmente a lo largo del día y vuelve a subir al final de la tarde. Los niveles de cortisol luego caen al final de la noche, alcanzando un punto mínimo durante la mitad de la noche. Esto corresponde al ciclo de reposo-actividad del organismo.[4] Un ciclo de cortisol circadiano anormalmente aplanado se ha relacionado con el síndrome de fatiga crónica,[5] el insomnio[6] y el agotamiento.[7]
El eje HPA tiene un papel central en la regulación de muchos sistemas homeostáticos en el cuerpo, incluidos el sistema metabólico, el sistema cardiovascular, el sistema inmunitario, el sistema reproductivo y el sistema nervioso central. El eje HPA integra influencias físicas y psicosociales para permitir que un organismo se adapte efectivamente a su entorno, use recursos y optimice la supervivencia.[4]
Las conexiones anatómicas entre áreas del cerebro como la amígdala, el hipocampo, la corteza prefrontal y el hipotálamo facilitan la activación del eje HPA.[8] La información sensorial que llega al aspecto lateral de la amígdala se procesa y se transmite al núcleo central de la amígdala, que luego se proyecta a varias partes del cerebro involucradas en las respuestas al miedo. En el hipotálamo, los impulsos de señales de miedo activan tanto el sistema nervioso simpático como los sistemas de modulación del eje HPA.
El aumento de la producción de cortisol durante el estrés da como resultado una mayor disponibilidad de glucosa para facilitar la lucha o huida. Además de aumentar directamente la disponibilidad de glucosa, el cortisol también suprime los procesos metabólicos altamente exigentes del sistema inmunitario, lo que da como resultado una mayor disponibilidad de glucosa.[4]
Los glucocorticoides tienen muchas funciones importantes, incluida la modulación de las reacciones de estrés, pero en exceso pueden ser dañinos. Se cree que la atrofia del hipocampo en humanos y animales expuestos a estrés severo es causada por una exposición prolongada a altas concentraciones de glucocorticoides. Las deficiencias del hipocampo pueden reducir los recursos de memoria disponibles para ayudar al cuerpo a formular reacciones apropiadas al estrés.
Sistema inmunitario
Hay comunicación bidireccional y retroalimentación entre el eje HPA y el sistema inmunológico. Varias citocinas, como IL-1, IL-6, IL-10 y TNF-alfa, pueden activar el eje HPA, aunque la IL-1 es la más potente. El eje HPA, a su vez, modula la respuesta inmunitaria, con altos niveles de cortisol que dan como resultado una supresión de las reacciones inmunitarias e inflamatorias. Esto ayuda a proteger al organismo de una sobreactivación letal del sistema inmunitario y minimiza el daño tisular por inflamación.[4]
El SNC es, en muchos sentidos, "inmune privilegiado", pero juega un papel importante en el sistema inmunológico y, a su vez, se ve afectado por él. El SNC regula el sistema inmunitario a través de vías neuroendocrinas, como el eje HPA. El eje HPA es responsable de modular las respuestas inflamatorias que ocurren en todo el cuerpo.[9][10]
Durante una respuesta inmunitaria, las citocinas proinflamatorias (por ejemplo, IL-1) se liberan en el sistema de circulación periférica y pueden atravesar la barrera hematoencefálica donde pueden interactuar con el cerebro y activar el eje HPA.[10][11][12] Las interacciones entre las citoquinas proinflamatorias y el cerebro pueden alterar la actividad metabólica de los neurotransmisores y causar síntomas como fatiga, depresión y cambios de humor.[10][11] Las deficiencias en el eje HPA pueden desempeñar un papel en las alergias y las enfermedades inflamatorias/autoinmunes, como la artritis reumatoide y la esclerosis múltiple.[9][10][13]
Cuando el eje HPA es activado por factores estresantes, como una respuesta inmunitaria, se liberan altos niveles de glucocorticoides en el cuerpo y suprimen la respuesta inmunitaria al inhibir la expresión de citocinas proinflamatorias (por ejemplo, IL-1, TNF alfa e IFN gamma) y aumentando los niveles de citocinas antiinflamatorias (por ejemplo, IL-4, IL-10 e IL-13), en células inmunitarias, como monocitos y neutrófilos[10][11][13][14]
La relación entre el estrés crónico y su activación concomitante del eje HPA y la disfunción del sistema inmunitario no está clara; los estudios han encontrado tanto inmunosupresión como hiperactivación de la respuesta inmune.[14]
Estrés
Estrés y enfermedad
El eje HPA está involucrado en la neurobiología de los trastornos del estado de ánimo y las enfermedades funcionales, incluidos el trastorno de ansiedad, el trastorno bipolar, el insomnio, el trastorno de estrés postraumático, el trastorno límite de la personalidad, el TDAH, el trastorno depresivo mayor, el agotamiento, el síndrome de fatiga crónica, la fibromialgia, el síndrome del intestino irritable, y alcoholismo.[15] Los antidepresivos, que se prescriben de forma rutinaria para muchas de estas enfermedades, sirven para regular la función del eje HPA.[16]
Las diferencias de sexo prevalecen en los seres humanos con respecto a los trastornos psiquiátricos relacionados con el estrés, como la ansiedad y la depresión, donde las mujeres experimentan estos trastornos con más frecuencia que los hombres.[17] Particularmente en roedores, se ha demostrado que las hembras pueden carecer de la capacidad de tolerar y procesar el estrés (particularmente para el estrés crónico) debido a la posible regulación a la baja de la expresión de GR, así como a una deficiencia de la proteína de unión a FKBP51 en el citosol. Al activar constantemente el eje HPA, esto podría conducir a mayores casos de estrés y trastornos que solo empeorarían con el estrés crónico.[18] Específicamente en roedores, las hembras muestran una mayor activación del eje HPA después del estrés que los machos. Es probable que estas diferencias también surjan debido a las acciones opuestas que tienen ciertos esteroides sexuales, como la testosterona y el estrógeno. El estrógeno funciona para mejorar la secreción de ACTH y CORT activada por el estrés, mientras que la testosterona funciona para disminuir la activación del eje HPA y funciona para inhibir las respuestas de ACTH y CORT al estrés.[19] Sin embargo, se requieren más estudios para comprender mejor la base subyacente de estas diferencias sexuales.
Los estudios experimentales han investigado muchos tipos diferentes de estrés y sus efectos sobre el eje HPA en muchas circunstancias diferentes.[20] Los factores estresantes pueden ser de muchos tipos diferentes: en estudios experimentales en ratas, a menudo se hace una distinción entre "estrés social" y "estrés físico", pero ambos tipos activan el eje HPA, aunque a través de vías diferentes.[21] Varios neurotransmisores de monoamina son importantes en la regulación del eje HPA, especialmente la dopamina, la serotonina y la norepinefrina (noradrenalina). Existe evidencia de que un aumento en la oxitocina, como resultado, por ejemplo, de interacciones sociales positivas, actúa para suprimir el eje HPA y, por lo tanto, contrarresta el estrés, promoviendo efectos positivos para la salud, como la cicatrización de heridas.[22]
El eje HPA es una característica de los mamíferos y otros vertebrados. Por ejemplo, los biólogos que estudiaron el estrés en los peces demostraron que la subordinación social conduce al estrés crónico, relacionado con la reducción de las interacciones agresivas, la falta de control y la amenaza constante impuesta por los peces dominantes . La serotonina (5HT) parece ser el neurotransmisor activo involucrado en la mediación de las respuestas al estrés, y los aumentos en la serotonina están relacionados con el aumento de los niveles plasmáticos de α-MSH, lo que provoca el oscurecimiento de la piel (una señal social en los peces salmónidos), la activación del eje HPA e inhibición de la agresión. La inclusión del aminoácido L-triptófano, un precursor de 5HT, en el alimento de la trucha arcoíris hizo que la trucha fuera menos agresiva y menos sensible al estrés.[23] Sin embargo, el estudio menciona que el cortisol plasmático no se vio afectado por el L-triptófano de la dieta. Se ha demostrado que el fármaco LY354740 (también conocido como Eglumegad, un agonista de los receptores metabotrópicos de glutamato 2 y 3) interfiere en el eje HPA, y la administración oral crónica de este fármaco conduce a una marcada reducción de los niveles basales de cortisol en los macacos de bonete (Macaca radiata); la infusión aguda de LY354740 resultó en una marcada disminución de la respuesta de estrés inducida por yohimbina en esos animales.[24]
Los estudios en personas muestran que el eje HPA se activa de diferentes maneras durante el estrés crónico según el tipo de factor estresante, la respuesta de la persona al factor estresante y otros factores. Los factores estresantes que son incontrolables, amenazan la integridad física o implican un trauma tienden a tener un perfil diurno alto y plano de liberación de cortisol (con niveles de cortisol más bajos de lo normal en la mañana y niveles más altos de lo normal en la noche) lo que resulta en un alto nivel general de liberación diaria de cortisol. Por otro lado, los factores estresantes controlables tienden a producir cortisol matutino más alto de lo normal. La liberación de la hormona del estrés tiende a disminuir gradualmente después de que ocurre un factor estresante. En el trastorno de estrés postraumático parece haber una liberación de cortisol más baja de lo normal, y se cree que una respuesta hormonal atenuada al estrés puede predisponer a una persona a desarrollar TEPT.[25]
También se sabe que las hormonas del eje HPA están relacionadas con ciertas enfermedades de la piel y la homeostasis de la piel. Existe evidencia que demuestra que las hormonas del eje HPA pueden estar relacionadas con ciertas enfermedades de la piel y tumores de la piel relacionados con el estrés. Esto sucede cuando las hormonas del eje HPA se vuelven hiperactivas en el cerebro.[26]
Estrés prenatal
Existe evidencia de que el estrés prenatal puede influir en la regulación de HPA. En experimentos con animales, se ha demostrado que la exposición al estrés prenatal provoca una respuesta de estrés HPA hiperreactiva. Las ratas que han estado estresadas prenatalmente tienen niveles basales elevados y un ritmo circadiano anormal de corticosterona en la edad adulta.[27] Además, requieren más tiempo para que sus niveles de hormonas del estrés vuelvan a la línea base después de la exposición a factores estresantes tanto agudos como prolongados. Los animales con estrés prenatal también muestran niveles de glucosa en sangre anormalmente altos y tienen menos receptores de glucocorticoides en el hipocampo.[28] En humanos, el estrés materno prolongado durante la gestación se asocia con un deterioro leve de la actividad intelectual y del desarrollo del lenguaje en sus hijos, y con trastornos del comportamiento como déficit de atención, esquizofrenia, ansiedad y depresión; el estrés materno autoinformado se asocia con una mayor irritabilidad, problemas emocionales y de atención.[29]
Cada vez hay más pruebas de que el estrés prenatal puede afectar la regulación de HPA en humanos. Los niños que estuvieron estresados prenatalmente pueden mostrar ritmos de cortisol alterados. Por ejemplo, varios estudios han encontrado una asociación entre la depresión materna durante el embarazo y los niveles de cortisol infantil.[30] El estrés prenatal también se ha implicado en una tendencia hacia la depresión y la falta de atención en la infancia.[31] No hay una indicación clara de que la desregulación del HPA causada por el estrés prenatal pueda alterar el comportamiento de los adultos.
Estrés de la vida temprana
El papel del estrés en la vida temprana en la programación del Eje HPA ha sido bien estudiado en modelos animales. Se ha demostrado que la exposición a factores estresantes leves o moderados en las primeras etapas de la vida mejora la regulación de HPA y promueve una resiliencia al estrés de por vida. Por el contrario, la exposición en la vida temprana al estrés extremo o prolongado puede inducir un eje HPA hiperreactivo y puede contribuir a la vulnerabilidad al estrés de por vida.[32] En un experimento ampliamente replicado, las ratas sometidas al estrés moderado del contacto humano frecuente durante las dos primeras semanas de vida habían reducido las respuestas de estrés hormonales y conductuales mediadas por HPA en la edad adulta, mientras que las ratas sometidas al estrés extremo de períodos prolongados de separación materna mostraron Mayores respuestas fisiológicas y conductuales al estrés en la edad adulta.[33]
Se han propuesto varios mecanismos para explicar estos hallazgos en modelos de ratas de exposición al estrés en la vida temprana. Puede haber un período crítico durante el desarrollo durante el cual el nivel de hormonas del estrés en el torrente sanguíneo contribuya a la calibración permanente del eje HPA. Un experimento ha demostrado que, incluso en ausencia de factores estresantes ambientales, la exposición temprana a niveles moderados de corticosterona se asoció con la resiliencia al estrés en ratas adultas, mientras que la exposición a dosis altas se asoció con la vulnerabilidad al estrés.[34]
Otra posibilidad es que los efectos del estrés de la vida temprana sobre el funcionamiento del HPA estén mediados por el cuidado materno. El manejo frecuente de las crías de rata por parte de los humanos puede hacer que su madre muestre un comportamiento más cariñoso, como lamerse y acicalarse. El cuidado maternal cariñoso, a su vez, puede mejorar el funcionamiento de HPA en al menos dos formas. En primer lugar, el cuidado materno es crucial para mantener el período normal de hiporrespuesta al estrés (SHRP), que en los roedores son las dos primeras semanas de vida durante las cuales el eje HPA generalmente no reacciona al estrés. El mantenimiento del período SHRP puede ser crítico para el desarrollo de HPA, y el estrés extremo de la separación materna, que interrumpe el SHRP, puede conducir a una desregulación permanente de HPA.[35] Otra forma en que el cuidado materno podría influir en la regulación de HPA es provocando cambios epigenéticos en la descendencia. Por ejemplo, se ha demostrado que el aumento de los lamidos y el aseo materno altera la expresión del gen del receptor de glutocorticoides implicado en la respuesta adaptativa al estrés.[32] Al menos un estudio en humanos ha identificado patrones de actividad neuronal materna en respuesta a estímulos de video de separación madre-hijo como asociados con una disminución de la metilación del gen del receptor de glucocorticoides en el contexto del trastorno de estrés postraumático derivado del estrés de la vida temprana.[36] Sin embargo, claramente, se necesita más investigación para determinar si los resultados observados en modelos animales intergeneracionales pueden extenderse a los humanos.
Aunque los modelos animales permiten un mayor control de la manipulación experimental, también se han estudiado los efectos del estrés de la vida temprana en la función del eje HPA en humanos. Una población que a menudo se estudia en este tipo de investigación son los sobrevivientes adultos de abuso infantil. Los sobrevivientes adultos de abuso infantil han exhibido mayores concentraciones de ACTH en respuesta a una tarea de estrés psicosocial en comparación con los controles no afectados y sujetos con depresión pero sin abuso infantil.[37] En un estudio, los sobrevivientes adultos de abuso infantil que no están deprimidos muestran una mayor respuesta de ACTH tanto al CRF exógeno como a la liberación normal de cortisol. Los sobrevivientes adultos de abuso infantil que están deprimidos muestran una respuesta de ACTH atenuada a la CRH exógena.[38] Una respuesta atenuada de ACTH es común en la depresión, por lo que los autores de este trabajo postulan que es probable que este patrón se deba a la depresión del participante y no a su exposición al estrés en la vida temprana.
Heim y sus colegas han propuesto que el estrés de la vida temprana, como el abuso infantil, puede inducir una sensibilización del eje HPA, lo que resulta en una actividad neuronal particularmente elevada en respuesta a la liberación de CRH inducida por el estrés.[38] Con la exposición repetida al estrés, el eje HPA sensibilizado puede continuar hipersecretando CRH del hipotálamo. Con el tiempo, los receptores de CRH en la hipófisis anterior se regularán a la baja, lo que producirá síntomas de depresión y ansiedad.[38] Esta investigación en sujetos humanos es consistente con la literatura animal discutida anteriormente.
El eje HPA estuvo presente en las primeras especies de vertebrados y ha permanecido altamente conservado por una fuerte selección positiva debido a sus roles adaptativos críticos.[39] La programación del eje HPA está fuertemente influenciada por el entorno perinatal y juvenil temprano, o "entorno de vida temprana".[40][41][42] El estrés materno y los diferentes grados de cuidado pueden constituir una adversidad temprana en la vida, que se ha demostrado que influye profundamente, si no altera permanentemente, el estrés de los hijos y los sistemas de regulación emocional.[40][41] Ampliamente estudiado en modelos animales (p. ej., lamer y acicalar/LG en cachorros de rata),[43] se ha demostrado que la consistencia del cuidado materno tiene una poderosa influencia en la neurobiología, la fisiología y el comportamiento de las crías. Mientras que el cuidado materno mejora la respuesta cardíaca, el ritmo de sueño/vigilia y la secreción de la hormona del crecimiento en el recién nacido, también suprime la actividad del eje HPA. De esta manera, el cuidado materno regula negativamente la respuesta al estrés en el recién nacido,[43] moldeando así su susceptibilidad al estrés en la edad adulta. Estos efectos de programación no son deterministas, ya que el entorno en el que se desarrolla el individuo puede coincidir o no coincidir con la reactividad del eje HPA "programada" y genéticamente predispuesta del primero. Aunque se conocen los principales mediadores del eje HPA, queda por dilucidar el mecanismo exacto mediante el cual se puede modular su programación durante los primeros años de vida. Además, los biólogos evolutivos cuestionan el valor adaptativo exacto de dicha programación, es decir, si una mayor reactividad del eje HPA puede conferir una mayor aptitud evolutiva.
Se han propuesto varias hipótesis, en un intento de explicar por qué la adversidad en la vida temprana puede producir resultados que van desde la vulnerabilidad extrema hasta la resiliencia, frente al estrés posterior. Se ha propuesto que los glucocorticoides producidos por el eje HPA confieren un papel protector o dañino, según las predisposiciones genéticas de un individuo, los efectos de programación del entorno de la vida temprana y la coincidencia o falta de coincidencia con el entorno posnatal de uno. La hipótesis de la adaptación predictiva (1), el concepto de vulnerabilidad y resiliencia de tres impactos (2) y la hipótesis de la mediación materna (3) intentan dilucidar cómo la adversidad en la vida temprana puede predecir diferencialmente la vulnerabilidad o la resiliencia frente a un estrés significativo en la edad adulta.[44] Estas hipótesis no son mutuamente excluyentes, sino que están altamente interrelacionadas y son únicas para el individuo.
- La hipótesis de la adaptación predictiva:[44] esta hipótesis contrasta directamente con el modelo de estrés de diátesis, que postula que la acumulación de factores estresantes a lo largo de la vida puede mejorar el desarrollo de la psicopatología una vez que se cruza un umbral. La adaptación predictiva afirma que la experiencia de vida temprana induce un cambio epigenético; estos cambios predicen o “preparan el escenario” para las respuestas adaptativas que se requerirán en su entorno. Por lo tanto, si un niño en desarrollo (es decir, del feto al recién nacido) está expuesto al estrés materno continuo y bajos niveles de atención materna (es decir, adversidad en la vida temprana), esto programará su eje HPA para que sea más reactivo al estrés. Esta programación habrá predicho y potencialmente será adaptable en un ambiente precario y altamente estresante durante la niñez y la vida posterior. Sin embargo, la previsibilidad de estos cambios epigenéticos no es definitiva; depende principalmente del grado en que el fenotipo genético y epigenéticamente modulado del individuo “coincide” o “no coincide” con su entorno.
- Concepto de tres golpes de vulnerabilidad y resiliencia:[44] esta hipótesis establece que dentro de un contexto de vida específico, la vulnerabilidad puede aumentar con la falla crónica para hacer frente a la adversidad en curso. Básicamente, busca explicar por qué, en circunstancias aparentemente indistinguibles, un individuo puede hacer frente al estrés con resiliencia, mientras que otro puede no solo hacer frente mal, sino que, en consecuencia, desarrollar una enfermedad mental relacionada con el estrés. Los tres "golpes" (cronológicos y sinérgicos) son los siguientes: predisposición genética (que predispone a una mayor o menor reactividad del eje HPA), entorno de vida temprana (perinatal, es decir, estrés materno, y postnatal, es decir, cuidado materno) y entorno de vida posterior (que determina coincidencia/desajuste, así como una ventana para cambios neuroplásticos en la programación temprana).[45] El concepto de coincidencia/desigualdad es fundamental para esta hipótesis evolutiva. En este contexto, aclara por qué la programación de la vida temprana en el período perinatal y posnatal puede haber sido seleccionada evolutivamente. Específicamente, al instaurar patrones específicos de activación del eje HPA, el individuo puede estar mejor equipado para hacer frente a la adversidad en un entorno de alto estrés. Por el contrario, si un individuo está expuesto a adversidades significativas en los primeros años de su vida, la reactividad del eje HPA aumentada puede "no encajar" con él/ella en un entorno caracterizado por un bajo nivel de estrés. El último escenario puede representar una mala adaptación debido a la programación temprana, la predisposición genética y la falta de coincidencia. Este desajuste puede entonces predecir resultados de desarrollo negativos, como psicopatologías en la edad adulta.
En última instancia, la conservación del eje HPA ha subrayado sus roles adaptativos críticos en los vertebrados, así como también en varias especies de invertebrados a lo largo del tiempo. El eje HPA juega un papel claro en la producción de corticosteroides, que gobiernan muchas facetas del desarrollo del cerebro y las respuestas al estrés ambiental continuo. Con estos hallazgos, la investigación en modelos animales ha servido para identificar cuáles son estos roles, con respecto al desarrollo animal y la adaptación evolutiva. En épocas más precarias y primitivas, un eje HPA aumentado puede haber servido para proteger a los organismos de los depredadores y las condiciones ambientales extremas, como el clima y los desastres naturales, fomentando la migración (es decir, la huida), la movilización de energía, el aprendizaje (frente a estímulos novedosos y peligrosos), así como un mayor apetito por el almacenamiento de energía bioquímica. En la sociedad contemporánea, la resistencia del eje HPA y la programación de la vida temprana tendrán implicaciones importantes para asesorar a las madres embarazadas y primerizas, así como a las personas que pueden haber experimentado una adversidad significativa en la vida temprana.[45]
Véase también
Referencias
- Malenka RC, Nestler EJ, Hyman SE (2009). «Chapter 10: Neural and Neuroendocrine Control of the Internal Milieu». En Sydor A, Brown RY, ed. Molecular Neuropharmacology: A Foundation for Clinical Neuroscience (2nd edición). New York: McGraw-Hill Medical. pp. 246, 248-259. ISBN 9780071481274. «
•The hypothalamic–neurohypophyseal system secretes two peptide hormones directly into the blood, vasopressin and oxytocin. ...
•The hypothalamic–pituitary–adrenal (HPA) axis. It comprises corticotropin-releasing factor (CRF), released by the hypothalamus; adrenocorticotropic hormone (ACTH), released by the anterior pituitary; and glucocorticoids, released by the adrenal cortex.
•The hypothalamic–pituitary–thyroid axis consists of hypothalamic thyrotropin-releasing hormone (TRH); the anterior pituitary hormone thyroid–stimulating hormone (TSH); and the thyroid hormones T3 and T4.
•The hypothalamic–pituitary–gonadal axis comprises hypothalamic gonadotropin–releasing hormone (GnRH), the anterior pituitary luteinizing hormone (LH) and follicle-stimulating hormone (FSH), and the gonadal steroids. » - Selye, Hans (1974). Stress without distress. Philadelphia: Lippincott. ISBN 978-0-397-01026-4.
- Cuzzo, Brian; Lappin, Sarah L. (2019), «Vasopressin (Antidiuretic Hormone, ADH)», StatPearls (StatPearls Publishing), PMID 30252325, consultado el 19 de octubre de 2019.
- editors, volume; Besedovsky, Hugo; Chrousos, George; Rey, Adriana Del (2008). The hypothalamus-pituitary-adrenal axis (1st edición). Amsterdam: Academic. ISBN 9780444530400.
- «Diurnal variation of adrenocortical activity in chronic fatigue syndrome». Neuropsychobiology 38 (4): 213-217. November 1998. PMID 9813459. doi:10.1159/000026543.
- Backhaus, Jutta; Junghanns, Klaus; Hohagen, Fritz (2004-10). «Sleep disturbances are correlated with decreased morning awakening salivary cortisol». Psychoneuroendocrinology 29 (9): 1184-1191. ISSN 0306-4530. PMID 15219642. doi:10.1016/j.psyneuen.2004.01.010.
- Pruessner, J.C.; Hellhammer, D.H.; Kirschbaum, C. (1999-03). «Burnout, perceived stress, and cortisol responses to awakening». Psychosomatic Medicine 61 (2): 197-204. ISSN 0033-3174. PMID 10204973. doi:10.1097/00006842-199903000-00012.
- Laura, Freberg (1 de enero de 2015). Discovering behavioral neuroscience : an introduction to biological psychology. Freberg, Laura,, Container of (work): Freberg, Laura. (Third edición). Boston, MA. p. 504. ISBN 9781305088702. OCLC 905734838.
- Marques-Deak, A; Cizza, G; Sternberg, E (February 2005). «Brain-immune interactions and disease susceptibility». Molecular Psychiatry 10 (3): 239-250. PMID 15685252. doi:10.1038/sj.mp.4001643.
- Otmishi, Peyman; Gordon, Josiah; El-Oshar, Seraj; Li, Huafeng; Guardiola, Juan; Saad, Mohamed; Proctor, Mary; Yu, Jerry (2008). «Neuroimmune Interaction in Inflammatory Diseases». Clinical Medicine: Circulatory, Respiratory, and Pulmonary Medicine 2: 35-44. PMC 2990232. PMID 21157520. doi:10.4137/ccrpm.s547.
- Tian, Rui; Hou, Gonglin; Li, Dan; Yuan, Ti-Fei (June 2014). «A Possible Change Process of Inflammatory Cytokines in the prolonged Chronic Stress and its Ultimate Implications for Health». The Scientific World Journal 2014: 780616. PMC 4065693. PMID 24995360. doi:10.1155/2014/780616.
- Hall, Jessica; Cruser, desAgnes; Podawiltz, Alan; Mummert, Diana; Jones, Harlan; Mummert, Mark (August 2012). «Psychological Stress and the Cutaneous Immune Response: Roles of the HPA Axis and the Sympathetic Nervous System in Atopic Dermatitis and Psoriasis». Dermatology Research and Practice 2012: 403908. PMC 3437281. PMID 22969795. doi:10.1155/2012/403908.
- Bellavance, Marc-Andre; Rivest, Serge (March 2014). «The HPA-immune axis and the immunomodulatory actions of glucocorticoids in the brain». Frontiers in Immunology 5: 136. PMC 3978367. PMID 24744759. doi:10.3389/fimmu.2014.00136.
- Padgett, David; Glaser, Ronald (August 2003). «How stress influences the immune response». Trends in Immunology 24 (8): 444-448. PMID 12909458. doi:10.1016/S1471-4906(03)00173-X. Archivado desde el original el 27 de marzo de 2016. Consultado el 12 de febrero de 2016.
- Spencer, R. L.; Hutchison, K. E. (1999). «Alcohol, aging, and the stress response». Alcohol Research & Health: The Journal of the National Institute on Alcohol Abuse and Alcoholism 23 (4): 272-283. ISSN 1535-7414. PMC 6760387. PMID 10890824.
- Pariante, Carmine M. (2003-08). «Depression, stress and the adrenal axis». Journal of Neuroendocrinology 15 (8): 811-812. ISSN 0953-8194. PMID 12834443. doi:10.1046/j.1365-2826.2003.01058.x.
- Rosinger, Zachary; Jacobskind, Jason; Park, Shannon; Justice, Nicholas; Zuloaga, Damian (2017). «Distribution of Corticotropin-releasing factor receptor 1 in the developing mouse forebrain: A novel sex difference revealed in the rostral periventricular hypothalamus». Neuroscience 361: 167-178. PMC 7173945. PMID 28823817. doi:10.1016/j.neuroscience.2017.08.016.
- Palumbo, Michelle C.; Dominguez, Sky; Dong, Hongxin (2020). «Sex differences in hypothalamic–pituitary–adrenal axis regulation after chronic unpredictable stress». Brain and Behavior (en inglés) 10 (4): e01586. PMC 7177572. PMID 32154650. doi:10.1002/brb3.1586.
- Handa, R. J.; Weiser, M. J.; Zuloaga, D. G. (2009). «A Role for the Androgen Metabolite, 5α-Androstane-3β,17β-Diol, in Modulating Oestrogen Receptor β-Mediated Regulation of Hormonal Stress Reactivity». Journal of Neuroendocrinology (en inglés) 21 (4): 351-358. PMC 2727750. PMID 19207807. doi:10.1111/j.1365-2826.2009.01840.x.
- Douglas, Alison J. (2005-03). «Central noradrenergic mechanisms underlying acute stress responses of the Hypothalamo-pituitary-adrenal axis: adaptations through pregnancy and lactation». Stress (Amsterdam, Netherlands) 8 (1): 5-18. ISSN 1025-3890. PMID 16019594. doi:10.1080/10253890500044380.
- «The hypothalamic-neurohypophysial system regulates the hypothalamic–pituitary–adrenal axis under stress: an old concept revisited». Frontiers in Neuroendocrinology 25 (3–4): 132-49. 2004. PMID 15589266. doi:10.1016/j.yfrne.2004.09.001.
- Detillion, Courtney E.; Craft, Tara K. S.; Glasper, Erica R.; Prendergast, Brian J.; DeVries, A. Courtney (2004-09). «Social facilitation of wound healing». Psychoneuroendocrinology 29 (8): 1004-1011. ISSN 0306-4530. PMID 15219651. doi:10.1016/j.psyneuen.2003.10.003.
- Winberg, S.; Øverli, Ø; Lepage, O. (2001-11). «Suppression of aggression in rainbow trout (Oncorhynchus mykiss) by dietary L-tryptophan». The Journal of Experimental Biology 204 (Pt 22): 3867-3876. ISSN 0022-0949. PMID 11807104. doi:10.1242/jeb.204.22.3867.
- Coplan, J. D.; Mathew, S. J.; Smith, E. L.; Trost, R. C.; Scharf, B. A.; Martinez, J.; Gorman, J. M.; Monn, J. A. et al. (2001-07). «Effects of LY354740, a novel glutamatergic metabotropic agonist, on nonhuman primate hypothalamic-pituitary-adrenal axis and noradrenergic function». CNS spectrums 6 (7): 607-612, 617. ISSN 1092-8529. PMID 15573025. doi:10.1017/s1092852900002157.
- Miller, Gregory E.; Chen, Edith; Zhou, Eric S. (2007-01). «If it goes up, must it come down? Chronic stress and the hypothalamic-pituitary-adrenocortical axis in humans». Psychological Bulletin 133 (1): 25-45. ISSN 0033-2909. PMID 17201569. doi:10.1037/0033-2909.133.1.25.
- Kim, Jung Eun; Cho, Baik Kee; Cho, Dae Ho; Park, Hyun Jeong (6 de julio de 2013). «Expression of hypothalamic-pituitary-adrenal axis in common skin diseases: evidence of its association with stress-related disease activity». Acta Dermato-Venereologica 93 (4): 387-393. ISSN 1651-2057. PMID 23462974. doi:10.2340/00015555-1557.
- Koehl, M.; Darnaudéry, M.; Dulluc, J.; Van Reeth, O.; Le Moal, M.; Maccari, S. (5 de septiembre de 1999). «Prenatal stress alters circadian activity of hypothalamo-pituitary-adrenal axis and hippocampal corticosteroid receptors in adult rats of both gender». Journal of Neurobiology 40 (3): 302-315. ISSN 0022-3034. PMID 10440731.
- Weinstock, M.; Matlina, E.; Maor, G. I.; Rosen, H.; McEwen, B. S. (13 de noviembre de 1992). «Prenatal stress selectively alters the reactivity of the hypothalamic-pituitary adrenal system in the female rat». Brain Research 595 (2): 195-200. ISSN 0006-8993. PMID 1467966. doi:10.1016/0006-8993(92)91049-k.
- Weinstock, Marta (2008-08). «The long-term behavioural consequences of prenatal stress». Neuroscience and Biobehavioral Reviews 32 (6): 1073-1086. ISSN 0149-7634. PMID 18423592. doi:10.1016/j.neubiorev.2008.03.002.
- Gutteling, Barbara M.; de Weerth, Carolina; Buitelaar, Jan K. (2004-12). «Maternal prenatal stress and 4-6 year old children's salivary cortisol concentrations pre- and post-vaccination». Stress (Amsterdam, Netherlands) 7 (4): 257-260. ISSN 1025-3890. PMID 16019591. doi:10.1080/10253890500044521.
- Buitelaar, Jan K.; Huizink, Anja C.; Mulder, Edu J.; de Medina, Pascalle G. Robles; Visser, Gerard H. A. (2003-05). «Prenatal stress and cognitive development and temperament in infants». Neurobiology of Aging. 24 Suppl 1: S53-60; discussion S67-68. ISSN 0197-4580. PMID 12829109. doi:10.1016/s0197-4580(03)00050-2.
- Flinn, Mark V.; Nepomnaschy, Pablo A.; Muehlenbein, Michael P.; Ponzi, Davide (2011-06). «Evolutionary functions of early social modulation of hypothalamic-pituitary-adrenal axis development in humans». Neuroscience and Biobehavioral Reviews 35 (7): 1611-1629. ISSN 1873-7528. PMID 21251923. doi:10.1016/j.neubiorev.2011.01.005.
- Liu, D.; Diorio, J.; Tannenbaum, B.; Caldji, C.; Francis, D.; Freedman, A.; Sharma, S.; Pearson, D. et al. (12 de septiembre de 1997). «Maternal care, hippocampal glucocorticoid receptors, and hypothalamic-pituitary-adrenal responses to stress». Science (New York, N.Y.) 277 (5332): 1659-1662. ISSN 0036-8075. PMID 9287218. doi:10.1126/science.277.5332.1659.
- «Developmental plasticity of HPA and fear responses in rats: a critical review of the maternal mediation hypothesis». Hormones and Behavior 50 (5): 667-80. December 2006. PMID 16890940. doi:10.1016/j.yhbeh.2006.06.015.
- de Kloet, E. Ronald; Sibug, Rosana M.; Helmerhorst, Frans M.; Schmidt, Mathias V.; Schmidt, Mathias (2005-04). «Stress, genes and the mechanism of programming the brain for later life». Neuroscience and Biobehavioral Reviews 29 (2): 271-281. ISSN 0149-7634. PMID 15811498. doi:10.1016/j.neubiorev.2004.10.008.
- Schechter DS, Moser DA, Paoloni-Giacobino A, Stenz A, Gex-Fabry M, Aue T, Adouan W, Cordero MI, Suardi F, Manini A, Sancho Rossignol A, Merminod G, Ansermet F, Dayer AG, Rusconi Serpa S (epub May 29, 2015). Methylation of NR3C1 is related to maternal PTSD, parenting stress and maternal medial prefrontal cortical activity in response to child separation among mothers with histories of violence exposure. Frontiers in Psychology.
- Heim C.; Newport D. J.; Heit S.; Graham Y. P.; Wilcox M.; Bonsall R.; Nemeroff C. B. (2000). «Pituitary-adrenal and autonomic responses to stress in women after sexual and physical abuse in childhood». JAMA 284 (5): 592-597. PMID 10918705. doi:10.1001/jama.284.5.592.
- Heim C.; Newport D.J.; Bonsall R.; Miller A.H.; Nemeroff C.B. (2001). «Altered Pituitary-Adrenal Axis Responses to Provocative Challenge Tests in Adult Survivors of Childhood Abuse». Am J Psychiatry 158 (4): 575-581. PMID 11282691. doi:10.1176/appi.ajp.158.4.575.
- Denver RJ (Apr 2009). «Structural and functional evolution of vertebrate neuroendocrine stress systems». Ann N Y Acad Sci 1163 (1): 1-16. Bibcode:2009NYASA1163....1D. PMID 19456324. doi:10.1111/j.1749-6632.2009.04433.x.
- «Brain development under stress: hypotheses of glucocorticoid actions revisited». Neurosci Biobehav Rev 34 (6): 853-66. May 2010. PMID 19631685. doi:10.1016/j.neubiorev.2009.07.006.
- Horton TH (Jan 2005). «Fetal origins of developmental plasticity: animal models of induced life history variation». Am. J. Hum. Biol. 17 (1): 34-43. PMID 15611963. doi:10.1002/ajhb.20092.
- Matthews SG (Mar 2000). «Antenatal glucocorticoids and programming of the developing CNS.». Pediatr Res 47 (3): 291-300. PMID 10709726. doi:10.1203/00006450-200003000-00003.
- Champagne, Frances A.; Francis, Darlene D.; Mar, Adam; Meaney, Michael J. (2003-08). «Variations in maternal care in the rat as a mediating influence for the effects of environment on development». Physiology & Behavior 79 (3): 359-371. ISSN 0031-9384. PMID 12954431. doi:10.1016/s0031-9384(03)00149-5.
- Daskalakis, Nikolaos P.; Bagot, Rosemary C.; Parker, Karen J.; Vinkers, Christiaan H.; de Kloet, E. R. (2013-09). «The three-hit concept of vulnerability and resilience: toward understanding adaptation to early-life adversity outcome». Psychoneuroendocrinology 38 (9): 1858-1873. ISSN 1873-3360. PMC 3773020. PMID 23838101. doi:10.1016/j.psyneuen.2013.06.008.
- Roth, Tania L.; Matt, Stephanie; Chen, Kenneth; Blaze, Jennifer (2014-12). «Bdnf DNA methylation modifications in the hippocampus and amygdala of male and female rats exposed to different caregiving environments outside the homecage». Developmental Psychobiology 56 (8): 1755-1763. ISSN 1098-2302. PMC 4205217. PMID 24752649. doi:10.1002/dev.21218.
Enlaces externos
- Mind-Body-Health.net page on HPA axis
- Understanding the stress response. Harvard Health Publishing. Universidad de Harvard 2020. En inglés.
| Control de autoridades |
|
|---|
 Datos: Q1369123
Datos: Q1369123 Multimedia: Hypothalamic-pituitary-adrenal axis / Q1369123
Multimedia: Hypothalamic-pituitary-adrenal axis / Q1369123