Electroforesis en gel
La electroforesis en gel es un grupo de técnicas empleadas por los científicos para separar moléculas basándose en propiedades como el tamaño, la forma o el punto isoeléctrico. La electroforesis en gel se utiliza generalmente con propósitos analíticos, pero puede ser una técnica preparativa para purificar moléculas parcialmente antes de aplicar espectrometría de masas, PCR, clonación o secuenciación de ADN.
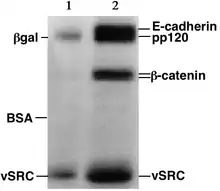
Significado

La segunda parte del núcleo, gel se refiere a la matriz usada para separar las moléculas. En muchos casos un gel es un polímero entrelazado de porosidad controlable. Cuando la separación es de proteínas, ADN o ácidos nucleicos pequeños (ADN, ARN o ribonucleótidos), el gel está compuesto por diferentes concentraciones de acrilamida/bis-acrilamida y un iniciador de la polimerización, para producir redes de poliacrilamida de diferentes tamaños. Para separar ácidos nucleicos grandes (más de unos centenares de bases), la matriz empleada es agarosa purificada. En ambos casos, el gel forma una matriz sólida pero porosa. La acrilamida, en contraste con la poliacrilamida, es una neurotoxina y debe ser manipulada con precaución.
La primera parte del nombre, electroforesis, se refiere a la fuerza electromotriz que es empleada para desplazar las moléculas a través del gel. Al situar las moléculas en el gel y aplicar una diferencia de potencial eléctrico, las moléculas se mueven a diferentes velocidades, hacia el cátodo, si están cargadas positivamente, y hacia el ánodo, si están cargadas negativamente (en una electroforesis, los electrodos se comportan igual que en una cubeta electrolítica).
Aplicaciones
Ácidos nucleicos
En los ácidos nucleicos la dirección de migración es del electrodo negativo al positivo, y esto es debido a la carga negativa presente en el esqueleto azúcar-fosfato. En los fragmentos de ADN dobles (con estructura de doble hélice) la velocidad de migración es inversamente proporcional a su tamaño. En fragmentos simples de ADN y ARN (una sola cadena), dichas moléculas tienden a plegarse de forma compleja y a migrar de forma más complicada, según la estructura terciaria formada tras el plegamiento. Sin embargo, compuestos que puedan romper los enlaces de puente de hidrógeno, como el hidróxido de sodio o la formamida, son empleados para 'desplegar' las moléculas plegadas y permitir que la velocidad de migración dependa únicamente y exclusivamente del tamaño y no de la estructura formada tras el plegamiento.[1]
Proteínas
Las proteínas no tienen una estructura predecible como los ácidos nucleicos, y por tanto sus velocidades de migración no son similares entre las proteínas. Incluso puede que no migren ni al aplicar una fuerza electromotriz (al encontrarse en su punto isoeléctrico). En estos casos, las proteínas se desnaturalizan mediante la adición de un detergente como el dodecilsulfato sódico/dodecilfosfato sódico (SDS/SDP) y un agente reductor como el 2-mercaptoetanol. Los detergentes otorgan una carga neta negativa a la proteína que les permite migrar a través del gel de poliacrilamida en relación directa a su masa, ya que la cantidad de cargas negativas que se unen a la proteína depende del tamaño de ésta, existiendo una relación carga/masa similar. Por otro lado, el agente reductor rompe los enlaces disulfuros, separando a la proteína en sus subunidades. Además, la desnaturalización hace que pierdan su estructura terciaria y cuaternaria por tanto su velocidad de migración es proporcional al tamaño y no a su estructura terciaria ni a su interacción con otras macromoléculas. Así, los más grandes se desplazan más lentamente.[1]
Concentración del gel
Concentración de agarosa
La concentración de agarosa dependerá siempre del tamaño del ácido nucleico que se desea analizar. Esto es importante, debido a que permitirá una mejor resolución de las muestras si el porcentaje de concentración es el adecuado. Usualmente los porcentajes de concentración van desde el 0.2% hasta un 6%, dependiendo del número de pares de bases (pb), como se observa en la Tabla 1. A mayor concentración del gel, los poros serán más pequeños por lo que se podrán apreciar aquellos fragmentos con un número pb menor y viceversa.[1]
Tabla 1. Recomendaciones de concentraciones de geles de agarosa para diferentes tamaños de ADN.[1]
| % de Agarosa | Tamaño del fragmento de ADN (pb) |
| 0.3 | > 700 pb |
| 0.5 | 700 pb a 25000 pb |
| 0.8 | 500 pb a 15000 pb |
| 1.0 | 250 pb a 12000 pb |
| 1.2 | 150 pb a 6000 pb |
| 1.5 | 80 pb a 4000 pb |
| 2.0 | 100 pb a 3000 pb |
| 3.0 | 500 pb a 1000 pb |
| 4.0 | 100 pb a 500 pb |
| 6.0 | 10 pb a 100 pb |
Concentración de poliacrilamida
Así como en el caso de la agarosa, determinar la concentración apropiada para las moléculas que se van a evaluar por medio de un gel de poliacrilamida resulta crucial. El tamaño de los poros de estos geles está determinada por la concentración final de acrilamida y biscacrilamida, de los cuales usualmente hay una proporción 19:1 para la observación de DNA o RNA desnaturalizado y una proporción de 29:1 para la mayoría de las proteínas y geles nativos para DNA y RNA (Tabla 2, Tabla 3). Si se regula adecuadamente la relación y concentración de acrilamida con poliacrilamida se podrán obtener diferentes porosidades, que serán menores que la de geles de agarosa.[2]
Tabla 2. Concentración de gel de poliacrilamida y relación de acrilamida y biscacrilamida para ADN/ARN nativo y desnaturalizado según su número de pb.[2]
| Proporción
Acrilamida/bisacrilamida |
% del gel | ADN/ARN
Nativo (bp) |
ADN/ARN
desnaturalizado (bp) |
|---|---|---|---|
| 19:1 | 4 | 100-1500 | 70-500 |
| 6 | 60-600 | 40-400 | |
| 8 | 40-500 | 20-200 | |
| 10 | 30-300 | 15-150 | |
| 12 | 20-150 | 10-100 | |
| 29:1 | 5 | 200-2000 | 70-800 |
| 6 | 80-800 | 50-500 | |
| 8 | 60-400 | 30-300 | |
| 10 | 50-300 | 20-200 | |
| 12 | 40-200 | 15-150 | |
| 20 | <40 | <40 |
Tabla 3. Concentración del gel de poliacrilamida para la visualización de proteínas según su peso molecular.[3]
| Proporción
Acrilamida/bisacrilamida |
% del gel | Peso molecular de la proteína (kDa) |
|---|---|---|
| 29:1 | 7.5 | 25-500 |
| 10 | 15-300 | |
| 12 | 10-200 | |
| 15 | 10-45 | |
| 20 | 5-40 |
Corriente eléctrica
Para generar la movilidad de las muestras a lo largo de sus carriles se requiere una fuente de poder que genere una corriente eléctrica. El voltaje que debe de ser aplicado depende del gel y de las muestras que se desean analizar. Usualmente la acrilamida es capaz de soportar una mayor corriente que la agarosa, sin embargo si el voltaje resulta ser demasiado, las bandas pueden verse difusas. Asimismo, si la corriente aplicada resulta ser excesiva para el sistema, puede provocar el derretimiento del gel, una curvatura en las bandas y un resolución. En caso de ser un voltaje insuficiente, la movilidad de las bandas más pequeñas se verá perjudicada y por lo tanto no se apreciará una adecuada visualización de las muestras .[2] Tomando todo esto bajo consideración, se recomienda voltajes de 25 a 100 V para geles de agarosa, mientras que para geles de poliacrilamida es importante conocer el largo del gel para determinar el voltaje apropiado. La distancia medida desde la parte superior hasta la parte inferior del gel se multiplica por un valor dentro del rango de 8-15 V, lo que permitirá determinar el voltaje de corrimiento apropiado .[1]
Revelado y visualización

Cuando se ha completado la electroforesis, las moléculas más pequeñas han llegado al ánodo. Entonces se pueden 'revelar' mediante la adición de un colorante específico para hacerlas visibles. Se emplean compuestos como el bromuro de etidio, para los ácidos nucleicos, o tinción de plata, azul de coomassie o tinción fluorescente, para las proteínas. Asimismo se emplean otros métodos para visualizar la separación de la mezcla en el gel. Si el reactivo es fluorescente bajo la luz UV, se puede simplemente hacer una fotografía de la placa bajo dicha luz. También, si las moléculas contienen átomos radiactivos se puede efectuar una autorradiografía.
Si se han inyectado varias mezclas una junto a otra en la placa, se producirán separaciones paralelas (carriles). Cada carril mostrará distintas bandas correspondientes a cada componente de la mezcla. Si dos componentes poseen la misma masa las separaciones serán incompletas, por lo cual ambas bandas se verán solapadas.
Las bandas en diferentes carriles que se encuentran a la misma altura contienen moléculas que han atravesado el gel a la misma velocidad. Existen marcadores especiales que contienen una mezcla de moléculas de tamaño conocido. Si se hace una electroforésis de una mezcla desconocida y se agrega un carril con un determinado marcador, las bandas observadas en el marcador pueden ser comparadas con las obtenidas en la mezcla desconocida para determinar su tamaño o punto isoeléctrico. Sabiendo que el desplazamiento de las moléculas es proporcional al logaritmo de la masa, se puede estimar el peso molecular de una molécula de interés comparándola con el patrón de migración de los marcadores. Esto se realiza construyendo una curva con los valores del marcador, dónde el eje (y) representa el logaritmo del peso molecular y el eje (x) representa el Rf de cada banda. Luego, se calcula el Rf de la molécula incógnita y se extrapola en la curva el logartimo del peso molecular. Aplicando el antilogarítmo se obtiene el peso molecular de la proteína incógnita.
Tipos
La electroforesis en gel se utiliza en biología molecular, genética y bioquímica:
- La electroforesis en gel de muestras grandes de ADN y ARN se efectúa en geles de agarosa.
- La electroforesis de proteínas se lleva a cabo en geles de poliacrilamida-SDS (SDS-PAGE), isoelectroenfoque, geles nativos o electroforesis bidimensional.
- Electroforesis capilar.
- Electroforesis de ADN.
- Zimografía o zimogramas
- Extracción en gel.
- Electroforesis en gel de campo pulsado.[4]
Referencias
- Salazar Montes, Adriana (2013). «Electroforesis». Biología Molecular. Mc Graw Hill. ISBN 9786071509123.
- Magdeldin, Sameh (2012). Gel electrophoresis -Principles and Basics (en inglés). InTech. ISBN 9789535104582.
- Análisis de proteínas. Electroforesis, transferencia e inmunodetección.. Advansta Corportation. 2011.
- Centers for Disease Control and Prevention (18 de julio de 2013). «Pulsed-field Gel Electrophoresis (PFGE)» (en inglés). Consultado el 7 de septiembre de 2013.
Bibliografía
- Bandow J, Baker JD, Berth M, Painter C, et al.: Improved image analysis workflow for 2-D gels enables large-scale 2-D gel-based proteomics studies - COPD biomarker discovery study. Proteomics 2008 (enlace roto disponible en Internet Archive; véase el historial, la primera versión y la última).
- Berth M, Moser FM, Kolbe M, et al: The state of the art in the analysis of two-dimensional gel eletcrophoresis images. Appl Microbiol Biotechnol. 2007;76(6):1223–43. (licencia Springer Open Access.)
Enlaces externos
- The state of the art in the analysis of two-dimensional gel electrophoresis images (enlace roto disponible en Internet Archive; véase el historial, la primera versión y la última).
- BIOtk - Análisis de proteinograma electroforético: BIOtk (enlace roto disponible en Internet Archive; véase el historial, la primera versión y la última).
| Control de autoridades |
|
|---|
 Datos: Q48255
Datos: Q48255 Multimedia: Gel electrophoresis / Q48255
Multimedia: Gel electrophoresis / Q48255