Spermatophyta
Las espermatofitas o fanerógamas (Spermatophyta) son un grupo monofilético del reino de las plantas (Plantae) que comprende a todos los linajes de plantas vasculares que producen semillas.
| Plantas con semilla (espermatofitas) | ||
|---|---|---|
| Rango temporal: Devónico Superior-reciente | ||
 | ||
| Taxonomía | ||
| Reino: | Plantae | |
| (sin rango) |
Embryophyta | |
| División: |
Spermatophyta Willkomm 1854 | |
| Subdivisiones | ||
El nombre científico proviene del griego σπέρμα ("sperma", que significa "semilla"), y φυτόν ("fiton", que significa "planta"), que se traduce como «plantas con semilla». La circunscripción del grupo (es decir, los taxones de los que está compuesto) coincide exactamente con la del antiguo taxón Phanerogamae, que por lo tanto es sinónimo de esta división. Debido a que en las espermatofitas el grano de polen produce un tubo (haustorial o polínico) para llegar al óvulo y que ocurra la fecundación, este grupo también es llamado de las embriofitas sifonógamas (del griego: embrios: embrión; fiton: planta; xifos: tubo; gamos: unión sexual. Literalmente, "plantas con embrión cuya unión sexual ocurre con tubo"). A veces la jerga científica se refiere a este grupo como "embriofitas", dejando fuera a las embriofitas asifonógamas o de los briófitos y los helechos y afines.
Hace mucho tiempo que los científicos consensúan la monofilia de las espermatofitas. Entre las evidencias morfológicas de la monofilia de las espermatofitas está, por supuesto, la semilla misma, y también la producción de madera (o "xilema secundario" generado en el meristema secundario llamado "cámbium"), al menos en forma ancestral. Otra característica notable es la ramificación axilar, en comparación con la ramificación dicotómica anisotónica de sus ancestros eufilofitos.
Las espermatofitas se originaron a fines del Devónico, a partir de lignofitas, que ya tenían producción de madera y ramificación axilar, como puede observarse en el registro fósil.
Hoy en día las espermatofitas son, por mucho, el linaje más extenso de plantas vasculares, con unas 270 000 especies vivientes. [1] Un solo subclado es el mayor responsable de esa diversidad: las angiospermas, o plantas con flores periantadas. Otros subclados, normalmente agrupados como gimnospermas, son las cícadas, los ginkgos, las coníferas y los gnetales. Estos cuatro grupos comparten un ancestro común. También se llaman "gimnospermas" a algunos fósiles de espermatofitas no productores de flores periantadas, que no comparten el mismo ancestro que las gimnospermas vivientes, por lo que algunos autores diferenciaban "Gymnospermae sensu lato" (pteridospermas + gimnospermas vivientes), que sería parafilético con respecto a las angiospermas y a "Gymnospermae sensu stricto", monofiléticos, comprendidos por los linajes vivientes.
Sinonimia
Este grupo ha sido denominado Phanerogama (Bartling 1830), Phanerogamae (Brongniart 1843, Eichler 1883), Spermatophyta (Willkomm 1854, Goebel 1882, Britton & A. Brown 1896), Anthophyta (Braun in Ascherson 1864, Wettstein 1924-1935), Siphonogamae (Engler 1886), Embryophyta siphonogama, (Engler 1892-1924), división Magnoliophyta (Takhtajan 1964) y Spermatophytina (Cavalier-Smith 1998, Ruggiero et al. 2015).
Ciclo de vida y estructuras reproductivas de espermatofitas
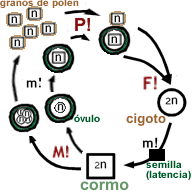
Referencias:
n: generación haploide,
2n: generación diploide,
m!: mitosis,
M!: meiosis,
F!: fecundación
Las espermatofitas pueden definirse como traqueofitas con las siguientes características:
- Como en todas las traqueofitas, hay dos generaciones multicelulares alternadas, denominados gametófito y esporófito, siendo el gametófito taloide, y el esporófito organizado en tejidos y órganos. El esporófito es un "cormo" (con sistema vascular, raíz y vástago). Como en todas las eufilofitas, el vástago del esporófito crece gracias a su meristema apical, y se ramifica como un tallo principal con ramas laterales, y además tiene hojas ("eufilos").
- En las espermatofitas la alternancia de generaciones gametofítica y esporofítica se da en forma enmascarada, ya que el gametófito se desarrolla por completo dentro de las estructuras del esporófito (incluso en los grupos más primitivos todavía pueden observarse anteridios y arquegonios).
- Poseen un ciclo de vida heterospórico, con gametófito de desarrollo endospórico (el esporófito da esporas femeninas dentro de las cuales se desarrolla el gametófito femenino, y esporas masculinas dentro de las cuales se desarrolla el gametófito masculino).
- Los esporangios nacen siempre en las hojas (las hojas fértiles son llamadas "esporofilos"). En las espermatofitas, las hojas fértiles, portadoras de esporangios, siempre están sobre una rama de crecimiento limitado ("braquiblasto"). Por lo tanto en el cormo aparece una nueva estructura, característica de las espermatofitas: la flor (una rama de crecimiento definido portadora de hojas fértiles). Morfológicamente en la flor puede diferenciarse un eje que se origina en el vástago y que presenta una o dos brácteas, que remata en un receptáculo donde se ubican los megasporofilos que darán las megasporas, y los microsporofilos o estambres que darán las microsporas. Los megasporofilos y los microsporofilos, en conjunto son los antofilos, por eso a este taxón también se lo conoce como Antophyta (pero no es recomendable utilizar ese nombre, porque algunos autores consideran como "antofitas" solo a las angiospermas y los gnetales, que antes se creía que tenían un antecesor común). En la flor veremos el desarrollo "enmascarado" del gametófito:
- en la flor las hojas fértiles portadoras de megaesporangios (los "megaesporofilos") contendrán el megaesporangio dentro del cual se produce la meiosis. 3 de los productos de la meiosis degeneran y el sobreviviente se desarrolla en la única espora femenina o "megaspora" del megasporangio, que a diferencia de lo que pasa en las demás plantas vasculares, nunca abandona al megasporangio, es una característica distintiva de las espermatofitas. La megaspora produce en su interior el gametofito femenino, que en su estado adulto puede ser de pocas (6) o muchas células, que tampoco abandona nunca a la megaspora ("gametofito endospórico", fenómeno común en plantas heterospóricas). El gametofito femenino a su vez produce en su interior una o varias gametas femeninas o "huevos" (como en todas las embriofitas, la gameta femenina nunca abandona al gametofito femenino). A esta nueva estructura (megasporangio + megaspora + gametofito + gameta) se la llama óvulo.
- La cubierta del megasporangio por mitosis va formando las cubiertas protectoras del futuro embrión (llamadas tegumentos, 1 en gimnospermas y 2 en angiospermas), puede haber una parte del megasporangio que se transforme en una cubierta de tejido de reserva (nucela).
- Como en espermatofitas la megaspora nunca abandona al megasporangio, el megasporangio de espermatofitas posee una abertura relativamente especializada por la selección natural para recibir el grano de polen y lograr la fecundación. Esta abertura del megaesporangio se llama micrópila. En gimnospermas el óvulo y su micrópila están expuestos, en angiospermas están cubiertos por el carpelo (hoja fértil portadora de esporangios). El gametofito femenino produce las gametas femeninas cerca de la abertura de la micrópila.
- en la flor las hojas fértiles portadoras de microesporangios (los "microesporofilos") se llamarán estambres y desarrollarán el microsporangio (que en los estambres de las espermatofitas se llama "saco polínico") que dará las microsporas, cada una contendrá un solo gametofito masculino (el gametofito masculino maduro cubierto por la pared de la microspora es el "grano de polen"). Los granos de polen serán liberados al exterior por "polinación" (apertura del saco polínico o microsporangio).
- en la flor las hojas fértiles portadoras de megaesporangios (los "megaesporofilos") contendrán el megaesporangio dentro del cual se produce la meiosis. 3 de los productos de la meiosis degeneran y el sobreviviente se desarrolla en la única espora femenina o "megaspora" del megasporangio, que a diferencia de lo que pasa en las demás plantas vasculares, nunca abandona al megasporangio, es una característica distintiva de las espermatofitas. La megaspora produce en su interior el gametofito femenino, que en su estado adulto puede ser de pocas (6) o muchas células, que tampoco abandona nunca a la megaspora ("gametofito endospórico", fenómeno común en plantas heterospóricas). El gametofito femenino a su vez produce en su interior una o varias gametas femeninas o "huevos" (como en todas las embriofitas, la gameta femenina nunca abandona al gametofito femenino). A esta nueva estructura (megasporangio + megaspora + gametofito + gameta) se la llama óvulo.
- Se da el nuevo fenómeno de polinización que es el transporte del polen hasta las estructuras de la flor femenina preparadas para recibirlo, mediante agentes externos. En gimnospermas el polen se transporta directamente a la micropila, en angiospermas se transporta al estigma del carpelo. Los agentes externos son principalmente el viento (fenómeno de anemofilia, común en gimnospermas) o los animales (fenómeno de zoofilia, común en angiospermas).
- Cuando la polinización es exitosa, el gametofito masculino crece, atraviesa la pared del grano de polen y emite un tubo polínico o haustorial, que emitirá los anterozoides poliflagelados —carácter primitivo— o los núcleos espermáticos —carácter avanzado— en el óvulo produciéndose la fecundación.
- Luego de la fecundación el gametofito femenino por mitosis va formando un tejido nutricio de reserva (haploide en gimnospermas, llamado protalo o endosperma primario, triploide en angiospermas por fecundación con un núcleo espermático del polen, llamado endosperma a secas).
- Como en todas las embriofitas, el cigoto por mitosis se transforma en un embrión, que creció nutrido por el gametofito femenino a través de una placenta. En las espermatofitas, el embrión al madurar entra en estado de latencia rodeado de las estructuras del gametofito —protalo o endosperma— y esporangio femeninos —nucela o no, 1 o 2 tegumentos— formando la semilla.
- el embrión es bipolar con meristema apical y meristema radical.
- la semilla madura es liberada convirtiéndose en la unidad de dispersión (a diferencia de la espora de pteridophytas).
- en condiciones adecuadas la semilla germina (despierta de la latencia), la primera fase de la germinación es la absorción de agua y luego se sigue desarrollando el embrión por mitosis, nutrido al principio de los tejidos de reserva de la semilla y luego de sus propios productos de la fotosíntesis, dando el nuevo esporófito adulto.
Evolución de espermatofitas

Como se puede observar de sus hermanos vivientes los helechos, las espermatofitas descienden de un ancestro caracterizado por la homosporía (un solo tipo de espora, gametofitos siempre bisexuales). Un paso crítico en el desarrollo de la semilla fue la evolución de la heterosporía: la producción de dos tipos de esporas, las megasporas y las microsporas, que darán los gametofitos femeninos y masculinos respectivamente.
La heterosporía se originó muchas veces en forma independiente en linajes no emparentados de plantas vasculares (hay ejemplos en lycophytas, en equisetopsidas y en polypodiopsidas), en muchos de esos casos el desarrollo de la heterosporía fue seguido de una reducción del número de megasporas funcionales. En la línea que condujo a las plantas con semilla, en el esporófito adulto que desarrolla sus megasporangios, se produce la meiosis de solo una célula por megasporangio, y por aborto de 3 de los productos de la meiosis, el número de megasporas funcionales es reducido a solo un megaspora funcional por megasporangio. Esa única megaspora funcional, en el linaje de las espermatofitas fue retenida dentro del megasporangio, desarrollando su gametofito femenino y gameta femenina por completo dentro del megasporangio de la generación esporofítica anterior. Finalmente el megasporangio desarrolló los tegumentos dejando abierto el pequeño orificio llamado micrópila.
Nuestro conocimiento de los orígenes de la semilla se basa principalmente en fósiles bien preservados del Devónico tardío y Carbonífero temprano, que fueron llamados "progimnospermas" o "helechos con semilla". Recordemos que la diferenciación entre un tallo principal y ramas laterales ya había evolucionado en el linaje de las eufilofitas. Lo primero que aparece en el linaje que derivó en las espermatofitas, fue encontrado en el Devónico tardío, y es la aparición de troncos muy grandes, con madera bastante similar en su estructura a la de las modernas coníferas. Estos troncos estaban conectados a enormes sistemas de ramas que portaban muchas hojas pequeñas. Archaeopteris, como ahora es llamada, fue descubierta como heterospórica, pero aún sin formar semillas.
La reconstrucción más exacta y la ubicación en el árbol filogenético de Archaeopteris y otras "progimnospermas" como Aneurophyton (Beck 1981, 1988, Beck y Wight 1988), fue fundamental para que los científicos puedan establecer tanto el origen de la heterosporía como el de la producción de madera, y llegar a la conclusión de que los dos eran anteriores a la evolución de la semilla. Por lo tanto es incorrecto afirmar que esos dos fósiles pertenecen a un clado que se llame espermatofitas ("plantas con semilla"), si bien son ancestros de las plantas con semilla actuales. Por lo tanto las espermatofitas y esos dos fósiles pertenecen a un grupo monofilético mayor, que fue llamado "Lignophyta" (Doyle y Donogue 1990), en referencia a que ya producían madera.
Análisis cuidadosos (por ejemplo el de Serbet y Rothwell 1992) han revelado que las primeras semillas estaban situadas en "cúpulas" ("cupules" en inglés) y cada semilla era cubierta por una excrecencia de la pared del esporangio que formaba una cámara especializada para recibir el polen, o "cámara polínica". Esta estructura probablemente se ayudaba de una secreción de una gota pegajosa ("gota de polinización") para capturar los granos de polen.
Los tejidos del tegumento externo probablemente derivan de una serie de esporangios estériles, que inicialmente tuvieron la forma de una serie de lóbulos en el ápice de la semilla, más que la forma de una micrópila diferenciada (ver gráfico).

Filogenia
Conocimientos actuales
Se ha demostrado —en contra de lo esperado— que el grupo de las gimnospermas vivientes es un grupo monofilético, hermano de las angiospermas.
Los análisis multigenéticos moleculares han demostrado que Spermatophyta se divide en dos grandes grupos: Gymnospermae y Angiospermae, los cuales fueron definidos por John Ray en 1703, y taxonómicamente esta clasificación se ha mantenido estable desde 1854.[2] Así pues, la filogenia más actualizada (2014) de las plantas vivientes con semilla presenta los siguientes subgrupos:[3]
| Spermatophyta |
| ||||||||||||||||||||||||||||||||||||||||||
Las diferencias entre los dos grupos están resumidas en el siguiente cuadro.
| Estructura | Gymnospermae | Angiospermae |
|---|---|---|
| Gametofito | Con anteridio, o arquegonio, o reducido, pero no mucho. | Muy reducido, a solo 3 células el masculino y a 7 células con 8 núcleos el femenino. |
| Gametas masculinas | Flageladas móviles | No hay gametas, hay solo núcleos espermáticos. |
| Tipo de fecundación | Fecundación simple. Por sinfonogamia o zoidiogamia | Doble fecundación (se fecunda la gameta femenina formando un cigoto 2n y una célula del gametofito femenino que dará el endosperma 3n). Siempre por sinfonogamia. |
| Tejidos del óvulo | Gameta femenina + el protalo (gametofito femenino haploide convertido en tejido de reserva) + 1 tegumento proveniente del megasporangio | Gameta femenina + el endosperma 3n + la nucela y los 2 tegumentos provenientes del megasporangio |
| Aparato vascular | Tipo traqueida | Tráqueas |
| Flor | Unisexual | Flores fundamentalmente hermafroditas |
| Brácteas adicionales de la flor | Flor bracteada, no tienen perianto, son aclamideas | Flor bracteada (con 1 o 2 profilos) y con perianto (compuesto por pétalos y sépalos) con función atractora de animales |
| Tipo de polinización | Por anemofilia (agente de transporte es el viento) | Típicamente por zoofilia (agente de transporte son los animales) |
| Unidad de dispersión | La semilla: desnuda o a lo sumo agrupada en pseudofrutos, el óvulo está expuesto al aire. | El fruto (semilla rodeada de la pared del carpelo) |
| Biotipo | Árboles o arbustos con lignina | Cualquier biotipo |
| Tipo de ramificación | Monopodial: todos los ejes están abiertos al crecimiento | Simpódica: los ejes se van cerrando al crecimiento. |
Grupos extintos
Una visión general de los grupos extintos relacionados con Spermatophyta sería la siguiente:
| Lignophyta |
| ||||||||||||||||||||||||
Visión histórica
Durante una gran parte del último siglo, los linajes de plantas con semilla, tanto vivientes como extintos, fueron comúnmente divididos en dos grandes grupos: las cycadofitas y las coniferofitas. Las cycadofitas, entre las que se incluyen las modernas cycadáceas, se distinguían por una producción de madera (crecimiento secundario del tallo) más bien limitada, con radios más bien anchos (lo que se llama "leño manoxílico"), y por hojas grandes, de tipo fronde de helecho, y semillas con simetría radial. En contraste, en las coniferofitas, entre las que se incluyen el Ginkgo y las coníferas, la madera está bien desarrollada y es densa ("leño picnoxílico"), las hojas son simples y muchas veces en forma de aguja, y las semillas tienen simetría dorsiventral (están "aplastadas"). Esta distinción sugirió a algunos investigadores que las plantas con semilla en realidad se originaron dos veces. Desde este punto de vista, la línea de las cycadofitas se derivó de un ancestro de tipo Progymnosperma, en el cual el sistema de ramificación lateral aplanado derivó en grandes hojas de tipo fronde de helecho. En cambio la línea de las espermatofitas habría derivado de un ancestro del tipo Archaeopteris, en el que las hojas individuales pueden haber sido modificadas en hojas de tipo aguja. Este escenario implica que la semilla misma fue originada dos veces, cada una correspondiéndose a un tipo de simetría diferente.
Sin embargo, en los análisis de filogenia que incluyeron los linajes vivientes junto con los representantes fósiles, en general aprueban el árbol filogenético que se muestra en la figura siguiente (ver por ejemplo Crane 1985, Doyle y Donoghue 1986, Nixon et al. 1994, Rothwell y Serbet 1994).

Estos estudios asumen que la semilla apareció una sola vez, y que las primeras plantas con semilla eran más bien parecidas a las cícadas, al menos en lo que respecta a las hojas grandes y pinadas, y las semillas con simetría radial. Específicamente, parece que una serie de "helechos con semilla" del Devónico-Carbonífero (Lygniopteris y medulosas) están situados en la base de la filogenia de las plantas con semilla, y que las coniferofitas están anidadas varios niveles hacia adentro del árbol, en un clado "platyspérmico" (de semillas con simetría dorsiventral). Este árbol hipotetiza que el cambio a hojas de tipo aguja y el cambio a semillas de simetría dorsiventral fueron posteriores a la aparición de las hojas y las semillas, y probablemente fueron una adaptación a ambientes de tipo árido.
A pesar de los enormes esfuerzos hechos hasta ahora para dilucidar las relaciones filogenéticas de los 5 grupos vivientes de espermatofitas (cícadas, ginkgos, coníferas, gnetofitas y angiospermas), utilizando tanto información morfológica como molecular, las relaciones aún no reciben consenso, y son motivo de variadas discusiones en el ambiente científico. En ese sentido cabe aclarar que está definitivamente descartado que las angiospermas deriven de una gnetofita ancestral. Los estudios morfológicos detallados muestran un origen diferente de los vasos xilemáticos en gnetofitas y en angiospermas (esto quiere decir que se originaron dos veces), y las estructuras de la flor de las gnetáceas (aquí definimos "flor" como "rama de crecimiento definido portadora de hojas fértiles") no son homólogas a las de la flor de angiosperma. Además, en el año 2004 se han encontrado fósiles de Gnetofitas que confirman (de forma morfológica) su pertenencia al grupo de las gimnospermas, y descarta nuevamente su relación con las angiospermas.
Hay que tener en cuenta que también se agrupaba dentro de las "gimnospermas" a muchos fósiles de plantas con semilla extintas (las pteridospermas) que en conjunto no forman un grupo monofilético: de hecho el grupo llamado Gymnosperma sensu lato se vuelve parafilético cuando uno toma en consideración los linajes de espermatofitas basales, así como otros "helechos con semilla" del Pérmico tardío y el Mesozoico, algunos de los cuales están probablemente en el linaje que derivó en las angiospermas.
Sistemática de espermatofitas
Clasificación
Los diferentes sistemas de clasificación le dan diferente importancia relativa a las estructuras de las espermatofitas:
Según el Sistema de clasificación de Engler se subdividen en:
- División Embryophyta siphonogama (=Spermatophyta)
- Subdivisión Gymnospermae (Cycadofilicales, Cycadales, Bennettitales, Ginkgoales, Coniferae, Cordaitales, Gnetales)
- Subdivisión Angiospermae
- Clase Monocotyledoneae
- Clase Dicotyledoneae
Según el sistema de Cronquist se dividen en:
- Progimnospermas (subdivisión Progimnospermophytina).
- Cicadofitinos (subdivisión Cycadice, Cycadophytina es un sinónimo) o gimnospermas de hoja pinnada.
- Pteridospermas o liginoptéridas (clase Pteridospermopsida).
- Benetitatas (clase Cycadeoidopsida).
- Cycadatas (clase Cycadatae, Cycadopsida es un sinónimo).
- Coniferofitinos (subdivisión Pinicae, Coniferophytina es un sinónimo) o gimnospermas de hoja dicótoma.
- Ginkgoatas (clase Gincoatae).
- Pinatas (clase Pinatae, Coniferopsida es un sinónimo).
- Gnetofitinos (subdivisión Gneticae, Gnetophytina es un sinónimo).
- Gnetatas o clamidospermas (clase Gnetoatae, Gnetopsida es un sinónimo).
- Angiospermas (subdivisión Magnoliophytina).
- Dicotiledóneas (clase Magnoliopsida).
- Monocotiledóneas (clase Liliopsida).
Diversidad de espermatofitas: grupos vivientes
Hoy en día hay dos linajes vivientes de espermatofitas: las gimnospermas y las angiospermas. Gymnospermae hace referencia a que poseen las semillas "desnudas" o no totalmente cubiertas por el carpelo, en oposición a las angiospermas o plantas con flores, cuyo carpelo cubre completamente a la semilla.

Cícadas
Fueron las más abundantes y diversas durante el Mesozoico. Hoy quedan alrededor de 130 especies. Las cícadas generalmente poseen un tronco bajo y ancho, con xilema secundario limitado, y hojas compuestas grandes parecidas a las de los helechos o las palmeras. Son dioicos, eso quiere decir que algunos esporófitos solo portan óvulos y luego semillas, y otros esporófitos solo portan estróbilos masculinos productores de polen. Los dos tipos de estróbilo son típicamente muy grandes, y en algunos casos de coloración brillante. Asimismo las semillas suelen ser grandes y usualmente tienen un tegumento externo carnoso y coloreado, presumiblemente una adaptación para atraer a los agentes de dispersión vertebrados. Muchas características de las cícadas pueden ser ancestrales, como el polen con tubo haustorial (en lugar de tubo polínico), y el esperma gigante multiflagelado (en lugar de los núcleos espermáticos). Sin embargo, las cícadas poseen características únicas que las alejan de las plantas con semilla ancestrales, que presumiblemente son caracteres derivados, entre los cuales se incluye la pérdida de la ramificación axilar, la presencia de trazas foliares "girdling" ("¿con fajas?"), y la producción de raíces coralloides que albergan cianobacterias fijadoras de nitrógeno. Dentro de las cícadas, los análisis filogenéticos indican que la primera división del grupo fue la que dividió al linaje de Cycas del resto. Por lo tanto Cycas estaría reteniendo algunos caracteres presumiblemente ancestrales, como los que han sido encontrados en parientes fósiles como Taeniopteris, a saber: el tener muchos óvulos nacidos en los márgenes de carpelos (carpelos definidos como hojas fértiles portadoras de óvulos) con mofología de tipo foliar, en lugar de tener dos óvulos por carpelo peltado que los sostiene apuntando hacia el eje del esporofilo (en su "cara adaxial"), que es el carácter que se encuentra en la otra línea. También en Cycas las hojas fértiles portadoras de óvulos no se encuentran agrupadas en estróbilos, como sí lo están en la otra línea.

Ginkgos
Solo hay una especie sobreviviente de ginkgos: Ginkgo biloba. Esta especie es muy raramente encontrada en forma silvestre, pero los árboles presentes en los templos de China fueron mantenidos por siglos por los monjes que los habitan, y en los tiempos modernos fue cultivado por el hombre en las veredas de las ciudades. Quizás la característica más distintiva del Ginkgo moderno es la producción de hojas deciduas, con forma de abanico ("flabeladas"), con venación dicotómica. Los ginkgos son bien conocidos en el registro fósil, donde se observa una gran diversidad en la morfología de las hojas. Como las cícadas, los ginkgos son dioicos (esporófitos diferentes portan o bien carpelos o bien estambres —estambres definidos como "hojas fértiles portadoras de sacos polínicos que contienen a los granos de polen"—). Los óvulos nacen en pares sobre ramas axilares que se piensa que son estróbilos reducidos. El tegumento del óvulo se diferencia en una capa externa carnosa (y olorosa) y una capa interna pétrea (dura) que encierra al gametofito femenino. También al igual que en las cícadas, los ginkgos retienen varios caracteres de las espermatofitas ancestrales, como el polen que emite un tubo haustorial (no polínico), y el esperma flagelado capaz de nadar.

Coníferas
Hay aproximadamente unas 600 especies de coníferas vivientes. Son árboles o arbustos con madera bien desarrollada y usualmente hojas de tipo aguja. Normalmente las hojas son solitarias, naciendo a lo largo del tallo, pero en los pinos (Pinus) están agrupadas en ramitas pequeñas. Las hojas usualmente tienen adaptaciones adicionales a la sequedad, por ejemplo estomas hundidos. Sin embargo, algunas coníferas del Hemisferio Sur (por ejemplo Podocarpus, Agathis) presentan hojas aplanadas y grandes, y en Phyllocladus se observan ramas aplanadas que parecen hojas. Muchas coníferas son monoicas (poseen carpelos y estambres en el mismo esporófito), pero algunos grupos son dioicos: Juniperus, Taxus, y Podocarpus. En los estróbilos masculinos (que en las coníferas se llaman conos masculinos o "pollen cones" en inglés), los estambres (o "microesporofilos") sostienen en su cara abaxial a los esporangios ("microsporangios") que darán gametofitos masculinos ("microgametofitos"). Los granos de polen son los gametofitos masculinos protegidos por una pared originada en el esporófito, y a veces tienen un par de apéndices llenos de aire parecidos a sacos, presumiblemente adaptaciones a la dispersión por el viento, pero estos "sacos aéreos" parecen haberse perdido en muchas líneas. Los óvulos receptivos, a diferencia de los microsporangios, están situados en la cara adaxial de cada carpelo o "escama ovulífera", mirando hacia el eje del cono femenino. La meiosis que dará las gametas, ocurre dentro de cada óvulo, y uno solo de los 4 productos de la meiosis se desarrollará hasta dar el gametofito femenino, siempre dentro del óvulo. El gametofito femenino o "protalo", produce uno o más gametos femeninos o "huevos" en el sector cercano a la micropila. Cuando el grano de polen llega finalmente a la micropila, el gametofito masculino encerrado en él desarrolla un tubo ("tubo polínico") que atraviesa la pared del gametófito femenino. Cuando el tubo polínico termina de crecer, el gametofito masculino emite a través de él dos espermas, que pueden ser células o núcleos celulares ("núcleos espermáticos") según el linaje. Es muy común en coníferas el fenómeno de "poliembronía", con muchos embriones desarrollándose en el mismo gametófito femenino, que puede ser o bien debido a que ocurrieron eventos de fertilización independientes en los que varios huevos fueron fertilizados por varios tubos polínicos distintos, o bien debido a que el embrión único se dividió en una etapa temprana en varios embriones genéticamente idénticos, siendo la última posibilidad más comúnmente encontrada que la primera. En las modernas coníferas, se dice que el estróbilo portador de polen es "simple", mientras que el portador de óvulos es "compuesto". Esto es debido a cómo es interpretada la morfología de los estróbilos: El cono masculino es interpretado como una rama modificada portadora de hojas fértiles o estambres, por lo tanto sería una "flor" única portadora de muchos estambres. En cambio el cono femenino es interpretado como derivado de una rama con hojas, que a su vez porta ramas laterales de crecimiento definido nacidas en la axila de las hojas, cada una de las ramas laterales portadoras de hojas fértiles o carpelos. Esta interpretación es sostenida por el registro fósil, que muestra una serie de pasos en la reducción de las ramas laterales portadoras de carpelos, hasta la aparición de la "escama ovulífera" altamente modificada que vemos en los grupos modernos (Florin 1951, 1954). También se observa que cada escama ovulífera está sostenida por una bráctea ("bráctea tectriz"), que representaría la hoja portadora de la rama lateral, también altamente modificada. En unas pocas coníferas, la bráctea es conspicua, emergiendo de entre las escamas ovulíferas (por ejemplo en Pseudotsuga mensiezii). Sin embargo, en muchas coníferas la bráctea es sumamente reducida. En las cupresáceas Taxodium y Cryptomeria, la bráctea está fusionada a la escama ovulífera, y la escama ovulífera aún muestra signos de presentar "hojas" (visibles como pequeños dientes). Los estudios filogenéticos han revelado algunas cuestiones interesantes acerca de la evolución de las coníferas. (por ejemplo Stefanovic et al. 1998). Los datos moleculares muestran una división basal entre las pináceas y un clado que albergaría a todas las demás coníferas. Las pináceas poseen varias características singulares, como óvulos invertidos (con la micropila mirando hacia el eje del cono) y las semillas aladas, alas que se originan en la escama ovulífera durante el desarrollo de la semilla. Dentro del otro clado de las coníferas, los dos grupos más grandes del Hemisferio Sur (Podocarpaceae y Araucariaceae) forman un clado, presumiblemente con la sinapomorfía de poseer un solo óvulo por escama ovulífera. Las cupresáceas están marcadas por muchas características singulares, como la fusión de la escama ovulífera con la báctea tectriz. A su vez, este grupo puede estar emparentado con las taxáceas, que tienen conos femeninos altamente reducidos con una sola semilla terminal, rodeada de un tercer tegumento ("arilo") carnoso y colorido.
Gnetales
Este grupo contiene solo unas 80 especies vivientes, que pertenecen a tres linajes bastante diferenciados (Doyle 1996, Friedman 1996, Price 1996). Uno es Ephedra, con alrededor de 40 especies distribuidas en los desiertos de todo el mundo, con hojas muy reducidas escamosas. Otro es Gnetum, con unas 35 especies en bosques tropicales del Viejo y Nuevo Mundo, con hojas con lámina entera muy parecidas a las vistas en la mayoría de las angiospermas. Finalmente, Welwitschia con una sola especie, Welwitschia mirabilis encontrada en el sudoeste de África, produce solo dos o raramente cuatro hojas funcionales a lo largo de toda su vida, hojas que crecen indefinidamente por meristemas presentes en la base, y se necrosan gradualmente en las puntas.
 Hábito de
Ephedra. |
Hojas y semillas de Gnetum gnemon
. |
.jpg.webp) Hábito de
Welwitschia mirabilis. |
Si bien estos tres clados se ven muy diferentes uno del otro, comparten muchas características inusuales, como las hojas opuestas, múltiples yemas por axila, vasos xilemáticos con aberturas circulares entre células adjuntas, polen compuesto, semillas en estróbilo, y un polen elipsoide ancestral con unas estrías características que corren de punta a punta. Las semillas también tienen dos tegumentos, el interno formando el tubo micropilar que exuda la gota de polinización, el externo derivado de un par de brácteas fusionadas. Los estudios moleculares también consensúan altamente la monofilia de este grupo. Dentro de las gnetofitas, Gnetum y Welwitschia forman un clado bien consensuado. Algunas de las sinapomorfías morfológicas son: hojas con venación reticulada, reducción aún mayor del gametófito masculino, y algunos aspectos de la estructura del gametófito femenino, como el desarrollo tetraspórico, la pérdida de los arquegonios, los núcleos libres funcionando como huevos en lugar de las células. El característico polen estriado encontrado en Ephedra y Welwitschia fue aparentemente perdido en la línea de la que derivó Gnetum, que tiene un polen con granitos con forma de pico, no aperturado. En lo que respecta al registro fósil, es más bien pobre salvo en los granos de polen. Solo algunos macrofósiles han sido descriptos (Crane 1996). Si bien los granos de polen de las gnetofitas son encontrados desde el Triásico, parece ser que el clado que contiene a los grupos modernos se ha diversificado más significativamente durante el Cretácico medio, al mismo tiempo que las angiospermas. Al igual que las angiospermas, las gnetofitas acortaron su ciclo de vida (y probablemente se volvieron herbáceas) y evolucionaron junto con los insectos para ser polinizadas por ellos, característica aún encontrada en algunos grupos vivientes. En marcado contraste con las angiospermas en cambio, las gnetofitas nunca se volvieron un componente significativo de la flora en paleolatitudes altas y medias, y han sufrido una disminución dramática de su representatividad durante el Cretácico tardío (Crane et al. 1995, Crane 1996).

Angiospermas
Con unas 257 000 especies vivientes, las angiospermas son las responsables de la mayor parte de la diversidad en espermatofitas, en embriofitas y en viridofitas. La fuerte evidencia de la monofilia de las angiospermas proviene de los estudios moleculares y de los muchos caracteres morfológicos compartidos por los miembros de este clado. De estos, algunos de los más obvios, que también son características reproductivas importantes, son: (1) las semillas son producidas dentro de un carpelo con una superficie estigmática que permite la germinación del polen, (2) el gametófito femenino es muy reducido, en la mayoría de las especies son solo 8 núcleos en 7 células, y (3) la doble fertilización, que llevó a la formación de un tejido nutritivo característico, triploide, llamado endosperma. Otras características son: (4) muchas angiospermas poseen vasos xilemáticos en lugar de traqueidas, carácter derivado dentro del grupo, en los vasos el agua puede fluir sin necesidad de atravesar una membrana, lo que los vuelve muy eficientes en el transporte de fluidos dentro del esporófito, pero probablemente también más propensos a recibir daño (en especial por embolias de aire) cuando están sujetos a estrés hídrico. (5) El floema de las angiospermas difiere del de todas las demás plantas en que los elementos del tubo criboso (que son células vivas, pero sin núcleo, encargadas del transporte de azúcares) están acompañadas por una o más "células acompañantes", que nacen de la misma célula madre que el elemento criboso.
Véase también
Referencias
- Judd, W. S. Campbell, C. S. Kellogg, E. A. Stevens, P.F. Donoghue, M. J. 2002. Plant systematics: a phylogenetic approach, Second Edition. Sinauer Axxoc, USA.
- Moritz Willkomm 1854, Anleitung zum Studium der wissenschaftlichen Botanik
- Cole & Hilger 2014, Tracheophyte Phylogeny, Vascular Plants – Systematics and Characteristic Archivado el 4 de marzo de 2016 en Wayback Machine. Freie Universität Berlin
Enlaces externos
 Wikcionario tiene definiciones y otra información sobre espermatófita.
Wikcionario tiene definiciones y otra información sobre espermatófita.
| Control de autoridades |
|
|---|
 Datos: Q25814
Datos: Q25814 Multimedia: Spermatophyta / Q25814
Multimedia: Spermatophyta / Q25814 Especies: Spermatophyta
Especies: Spermatophyta