Europasaurus holgeri
Europasaurus holgeri ("lagarto de Europa Holger Lüdtke") es la única especie conocida del género extinto Europasaurus de dinosaurio saurópodo braquiosáurido basal, que existieron a finales del Jurásico, hace aproximadamente 154 millones de años, durante el Kimmeridgiense, en lo que hoy es Europa. Fue encontrado en el norte de Alemania y ha sido identificado como un ejemplo de enanismo insular, resultados del aislamiento de la población de saurópodos en una isla dentro de la cuenca de Baja Sajonia.
| Europasaurus holgeri | ||
|---|---|---|
| Rango temporal: 154 Ma Jurásico superior | ||
 | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Sauropsida | |
| Superorden: | Dinosauria | |
| Orden: | Saurischia | |
| Suborden: | Sauropodomorpha | |
| Infraorden: | Sauropoda | |
| (sin rango): | Macronaria | |
| Familia: | Brachiosauridae? | |
| Género: |
Europasaurus Mateus, Laven, & Knötschke vide Sander et al., 2006 | |
| Especie: |
E. holgeri Mateus, Laven, & Knötschke vide Sander et al., 2006 | |
Descripción

Europasaurus es un saurópodo muy pequeño, que mide solo 6,2 metros como adulto. Esta longitud se estimó basándose en un fémur parcial, escalado al tamaño de un espécimen casi completo de Camarasaurus. Se conocen individuos más jóvenes, desde tamaños de 3,7 metros hasta el juvenil más joven a 1.75 metros.[1][2] El titanosaurio Magyarosaurus tiene un tamaño similar, con 900 kilogramos en comparación con el Europasaurus con aproximadamente 800 kilogramos. Otro pequeño titanosauriano, Neuquensaurus, es más grande que cualquiera de los taxones, tiene una longitud de hasta 9 metros y pesa 3200 kilogramos.[3]
Además de ser un neosauropodo muy pequeño, se pensaba que Europasaurus tenía múltiples características morfológicas únicas para distinguirlo de parientes cercanos por sus descriptores originales, Sander et al. en 2006.[1] El proceso nasal del premaxilar se pensaba que era curva en sentido anterior mientras que se proyecta hacia arriba, ahora conocido por ser un artefacto de la preservación,[4] hay una muesca en la superficie superior del cuerpo de las vértebras cervicales, la escápula tiene un proceso prominente en la superficie posterior de su cuerpo, y el astrágalo, un hueso del tobillo, es dos veces más ancho que alto.[1]
Cuando se compara con Camarasaurus, Europasaurus tiene una morfología diferente del postorbital en el que la pestaña posterior no es tan corta, un contacto corto entre los huesos nasales y frontales del cráneo, la forma de su parietal, rectangular en Europasaurus, y las espinas neurales de sus vértebras en frente de la pelvis que no están divididas. También se mencionaron las comparaciones con Brachiosaurus brancai, ahora llamado Giraffatitan, y se identificó que Europasaurus tiene un hocico más corto, un contacto entre el cuadratojugal y el escamoso y un húmero, hueso del miembro anterior superior, que ha aplanado y alineado a las superficies proximales y distales . Finalmente, hubo comparaciones rápidas con el braquisáurido potencial Lusotitan , que tiene una forma diferente de ilion y astrágalo, y Cetiosaurus humerocristatus, llamado ahora Duriatitan, que tiene una cresta deltopectoral que es menos prominente y se extiende a través del húmero.[1]
Diagnosis y morfología comparativa

Sander et al. en 206 notó que Europasaurus holgeri exhibía los siguientes estados inequívocos autapomorficos, "protuberancia nasal de proyección premaxilar anterodorsalmente, la muesca medial es un margen posterior dorsal del centro de vértebras cervicales; acromión escapular con proyección posterior prominente; y anchura transversal del astragálo, el doble de su altura dorsoventral y ancho anteroposterior." Comparando Europasaurus con Camarasaurus, Sander et al. en 2006 declararon que se diferencia "en la protuberancia posterior con forma de ala del postorbital siendo levemente más larga y ancha que la protuberancia anterior, considerando que es mucho más corta en Camarasaurus." Otras diferencias incluyen el contacto nasal-frontal más corto en Europasaurus y un hueso parietal el cual es rectangular en vista posterior. Al contrario de Camarasaurus, las espinas nerviosas de las vértebras presacras no están divididas. Comparando Europasaurus con Giraffatitan (Brachiosaurus) brancai, Sander et al. en 2006, encontró que el género anterior se diferencia del último al tener un hocico más corto, un cuadratoyugal el cual hace contacto con el escamosal, y un húmero allanado anteromedialmente poseyendo epífisis no alineados proximales y distales. Europasaurus fue también comparado con los macronarianos Lusotitan y Duriatitan (Cetiosaurus) humerocristatus y se lo encontró distinto de ambas formas. Finalmente, Europasaurus puede distinguirse de la mayoría de miembros conocidos de Neosauropoda por su tamaño corporal de adulto muy pequeño.[1]
Cráneo
Casi todos los huesos externos del cráneo se han conservado entre los especímenes de Europasaurus , excepto los prefrontales. Algunos huesos adicionales solo están representados por fósiles muy desinformados y fragmentados, como los lacrimales . Se conocen ocho premaxilares, con una forma de hocico generalmente rectangular como la que se encuentra en Camarasaurus. La proyección anterior de la premaxila identificada en Sander et al. en 2006[1] se volvió a identificar como un artefacto de conservación en Marpmann et al. en 2014,[4] similar a la anatomía encontrada en Camarasaurus y Euhelopus en menor grado. La proyección dorsal de la premaxila, la que entra en contacto con el hueso nasal, comienza como una proyección postero-dorsal, antes de volverse verticalmente recta en el punto del foramen subnarial, hasta que alcanza el nasal. Este "paso" débil se ve en Camarasaurus y Euhelopus y está más presente en Abydosaurus, Giraffatitan y un posible cráneo de Brachiosaurus. Estos últimos taxones también tienen un hocico más largo, con más distancia desde el primer diente hasta el proceso nasal de la premaxila. Además, Europasaurus comparte con los camarasauromorfos basales, braquiosáuridos, Camarasaurus, Euhelopus y Malawisaurus una órbita de tamaño similar a la fenestra nasal, mientras que la abertura nasal se reduce significativamente en los titanosaurios derivados, Rapetosaurus, Tapuiasaurus y Nemegtosaurus.[4]

Un solo maxilar está presente en el material bien conservado de Europasaurus, DFMMh/FV 291.17. Este maxilar tiene un cuerpo largo, con dos procesos alargados, uno nasal y uno posterior. Solo hay un proceso lagrimal débil, como en la mayoría de los saurópodos, excepto el Rapetosaurus. El proceso nasal es alargado y cubre el borde anterior y ventral de la fenestra antorbital . Este proceso se extiende alrededor de 120º desde la hilera de dientes horizontal. La base del proceso nasal también forma el cuerpo del proceso lagrimal, y en su divergencia está la fenestra antorbital, similar en forma a las de Camarasaurus , Euhelopus , Abydosaurus y Giraffatitan, pero alrededor de 1/2 más alto proporcionalmente. La fenestra pre-antorbital, una pequeña abertura delante o debajo de la abertura antorbital, está bien desarrollada en taxones como Diplodocus y Tapuiasaurus y está casi ausente, como en Camarasaurus y Euhelopus. Hubo alrededor de 12-13 dientes en total en el maxilar superior del Europasaurus , menos que en más taxones basales, 16 dientes en Jobaria y 14-25 en Atlasaurus, pero cayendo dentro del rango de variación en Brachiosauridae, 15 en Brachiosaurus a 10 en Abydosaurus. Todos los dientes sin usar conservados muestran hasta cuatro pequeños dentículos en sus bordes mesiales. Una pequeña cantidad de las coronas dentales posteriores están ligeramente torcidas cerca de 15º, pero mucho menos que en los braquiosáuridos que es de entre 30°-45°.[4]
Los huesos nasales de Europasaurus, que se conocen varios, pero pocos están completos o sin distorsión. Las nasales se superponen posteriormente por los huesos frontales y hacia el lado se articulan sin rodeos con los prefrontales. A diferencia de las nasales de Giraffatitan, los de Europasaurus se proyectan horizontalmente hacia adelante, formando una pequeña porción del techo del cráneo sobre la fenestra antorbital. Se conocen cuatro frontales de Europasaurus, tres de la izquierda y uno de la derecha. Debido a su desarticulación, es probable que los frontales nunca se fusionen durante el crecimiento, a diferencia de Camarasaurus. Los frontales forman una porción del techo del cráneo, que se articula con otros huesos como los nasales, parietales, prefrontales y posorbitales y son más largos que anchos, un carácter único entre los eusauropodos . Al igual que en los diplodocoides, Amargasaurus , Dicraeosaurus y Diplodocus así como en Camarasaurus, los frontales se excluyen de la fenestra frontoparietal o fenestra parietal cuando se excluyen los frontales. Los frontales también están excluidos del margen de la fenestra supratemporal, algo generalizado en saurópodos más derivado de Shunosaurus y solo tienen una participación pequeña, no decorada en la órbita. Se conocen varios huesos parietales en Europasaurus , que muestran una forma rectangular mucho más ancha que larga. los parietales también son anchos cuando se ven desde la parte posterior del cráneo, siendo ligeramente más altos que el foramen magnum, la abertura de la médula espinal. Los parietales contribuyen a aproximadamente la mitad del borde de la fenestra post-temporal (que se abre por encima de la parte posterior del cráneo), con la otra región encerrada por los huesos escamosos y algunos huesos del cerebro. Los parietales también forman parte del borde de la fenestra supratemporal, que es más ancha que larga en Europasaurus, como en Giraffatitan, Camarasaurus y Spinophorosaurus. Además de la fenestra antes mencionada, los parietales también tienen una "fenestra postparietal", algo que rara vez se ve fuera de Dicraeosauridae. Un hueso postorbital trirradiado está presente en el Europasaurus, que evolucionó como la fusión del hueso posfrontal y postosbital de los taxones más basales. Entre los procesos de proyección anterior y ventral, el posorbital forma el margen de la órbita, y entre los procesos posterior y ventral limita con la fenestra infratemporal .[4]
Se conocen múltiples yugales de Europasaurus , que son más similares en morfología a los sauropodomorfos basales que otros macronarios. Forma parte del borde de la órbita, fenestra infratemporal y el borde inferior del cráneo, pero no alcanza la fenestra antorbital. El proceso posterior del yugal es muy frágil y estrecho, mostrando una cicatriz ósea de la articulación con el cuadratojugal. Hay dos prominencias que se proyectan desde la parte posterior del cuerpo yugal, que divergen a 75° y forman los bordes inferior y frontal de la fenestra infratemporal. Como en Riojasaurus y Massospondylus, dos sauropodomorfos no saurópodos, el yugal forma una gran parte del borde de la órbita, desde la parte posterior hasta la esquina inferior delantera. Esta característica se ha visto en embriones de titanosaurios, pero no en individuos adultos. El hueso cuadratojugal es un elemento alargado que tiene dos brazos salientes, uno anterior y otro dorsal. Como en otros saurópodos, el proceso anterior es más largo que el dorsal, pero en el Europasaurus los brazos tienen longitudes más similares. El proceso horizontal es paralelo a la fila de dientes de Europasaurus , similar a Camarasaurus pero a diferencia de Giraffatitan y Abydosaurus. Hay una pestaña ventral prominente en el brazo anterior del hueso, que es posiblemente una sinapomorfia de Brachiosauridae, aunque también se encuentra en algunos individuos con Camarasaurus. Los dos procesos cuadráticos divergen en un ángulo casi recto, aunque el proceso dorsal se curva a medida que sigue la forma del cuadrante . Los escamosos encontrados en Europasaurus muestran la misma forma aproximada en vista lateral que Camarasaurus, la de un signo de interrogación. Los escamosos se articulan con muchos huesos del cráneo, incluidos los del techo del cráneo, los del cráneo ventral y los del cerebro. Al igual que los postorbitales, los escamosos son triradiados, con un ventral, anterior y proceso medial .[4]
Hay trece elementos conservados en el paladar de Europasaurus , incluyendo el cuadrado, pterigoideo y ectopterigoideo . Los cuadrandos se articulan con los huesos del paladar y el cerebro, así como con los huesos externos del cráneo. Son similares en forma a los de Giraffatitan y Camarasaurus y tienen superficies articulares bien desarrolladas. Un solo eje está presente para la mayoría de la longitud de los cuadrados, con un ala pterigoideo a lo largo del lado medial. Los pterigoideos son los huesos más grandes del paladar saurópodo, y tienen una forma de triradiado, como los posorbitales. Una proyección anterior entra en contacto con el pterigoideo opuesto, mientras que un ala lateral hace contacto con el ectopterigoide, y un ala posterior sostiene el cuadrado y el basiptergoide,un hueso que proporciona la conexión entre el paladar y el cerebro. El ectopterigoide es un hueso del paladar pequeño, que articula los huesos del paladar central (pterigoideo y palatino ) con el maxilar superior. Los ectoptergoides tienen forma de L, con un proceso anterior que se adhiere al maxilar y un proceso dorsal que se encuentra con el pterigoideo.[4]

Vértebras
Las vértebras cervicales de Europasaurus son las mejor conservadas y más representadas de la columna vertebral. Sin embargo, no se conoce todo el cuello, por lo que el número cervical podría estar entre Camarasaurus, 12 vértebras y Rapetosaurus, 17 vértebras. Además, las vértebras cervicales múltiples provienen de individuos de diferentes edades, y se sabe que la longitud central y la estructura interna cambian a lo largo del desarrollo. Los centros cervicales adultos son alargados y opistocelélicos, el extremo anterior tiene forma de bola, con una muesca en la parte superior del extremo posterior del centro. Esta característica se describió como característica de Europasaurus, pero también se conoce en Euhelopus y Giraffatitan. En el lado de los centros del Europasaurus hay una excavación que se abre a la neumaticidad interna de las vértebras. A diferencia de Giraffatitan y braquiosáuridos, Europasaurus no tiene crestas delgadas o láminas que dividen esta abertura. Europasaurus comparte características de las láminas en las vértebras superiores con macronarios basales y braquiosauridos. A diferencia de las cervicales anteriores y medias, las vértebras cervicales posteriores son menos alargadas y proporcionalmente más altas, como en otros macronarios, con cambios significativos en las posiciones de las superficies articulares.[5]
Piel
Entre los macronarios, las impresiones de piel fosilizadas solo se conocen de Haestasaurus , Tehuelchesaurus y Saltasaurus. Tanto Tehuelchesaurus como Haestasaurus pueden estar estrechamente relacionados con Europasaurus como macronarios basales, y las características de todas las impresiones de piel de saurópodos son similares. Haestasaurus, el primer dinosaurio conocido por las impresiones de la piel, conservó el tegumento en una parte del brazo alrededor de la articulación del codo de aproximadamente 19,5 por 21,5 centímetro cuadrados de área.[6] Las impresiones dérmicas están más difundidas en el material de Tehuelchesaurus, donde se conocen de las zonas de la extremidad anterior, escápula y torso. No hay placas óseas ni nódulos, lo que indica armadura, pero hay varios tipos de escamas. Los tipos de piel del Tehuelchesaurus son en general más similares a los encontrados en diplodócidos y Haestasaurus que en los embriones de titanosáurido de Auca Mahuevo.[7] Como la forma y la articulación de los tubérculos preservados en estos macronarios basales son similares en otros taxones donde se conserva la piel, incluidos los especímenes de Brontosaurus excelsus y diplodocoides intermedios, tales estructuras dérmicas probablemente se encuentran generalizadas en toda Neosauropoda.[6]
Descubrimiento e investigación
Alrededor de 1995, se descubrieron múltiples huesos pequeños de saurópodos en una cantera abierta en la montaña Langenberg, cerca de la ciudad de Oker, Alemania. La cantera de 203 metros expone rocas bien datadas de la cuenca de la Baja Sajonia que datan de los principios de Oxfordiense al Kimmeridgiense. A lo largo de los próximos 10 años, cerca de 650 huesos individuales fueron recuperados y almacenados en el Dinosaur Park Münchehagen ubicado cerca de Hanover. La variedad de huesos de saurópodos se encuentra dentro de un área de 60–80 metros cuadrados, e incluye individuos articulados de diversas maneras.[2] Dentro de la cantera, Europasaurus los fósiles están restringidos a una cama individual, la cama 83, que pertenece a la parte superior del Kimmeridgiense inferior.[8] De estos especímenes, se seleccionó el holotipo , un individuo desarticulado pero asociado.[2]
El holotipo incluye múltiples huesos craneales, premaxilar, maxilar y cuadratojugal, una base de cerebro parcial, múltiples huesos de la mandíbula, dentario, surangular y angular, grandes cantidades de dientes, vértebras cervicales, vértebras sacras y costillas del cuello y el torso. Este espécimen está catalogado bajo el número de ejemplar DFMMh / FV 291. Al menos otros 10 individuos fueron remitidos al mismo taxón basándose en la superposición del material. En la descripción de 2006 , estos individuos fueron nombrados como el nuevo género relacionado con Brachiosauridae , Europasaurus holgeri. La descripción y el documento de nombramiento fueron escritos por Martin P. Sander, Octávio Mateus, Thomas Laven y Nils Knötschke. La etimología dada para el nombre del género es "reptil de Europa", y el nombre específico honra a Holger Lüdtke, el descubridor de los fósiles de saurópodos.[1]
Parte del material fósil de Europasaurus se dañó o destruyó por incendio provocado en la noche del 4 al 5 de octubre de 2003. El incendio destruyó el laboratorio y la sala de exposiciones del Dinosaur Park Münchehagen, lo que provocó la pérdida de 106 huesos, lo que Representa el 15% de los huesos preparados al momento. Además, el fuego afectó a la mayoría de los bloques que aún no estaban preparados, y el agua de extinción de incendios golpeó la piedra caliente y causó un desmoronamiento adicional.[9] Los especímenes destruidos incluyen DFMMh / FV 100, que incluye la serie vertebral mejor conservada y la única pelvis completa.[5][9]
Clasificación
Cuando se nombró por primera vez, se consideraba que Europasaurus era un taxón dentro de Macronaria que no pertenecía a la familia Brachiosauridae o al clado Titanosauromorpha. Esto indicó que el enanismo del taxón era un resultado de la evolución, en lugar de ser una característica de un grupo.[1] Se analizaron tres matrices con la inclusión de Europasaurus, la de Wilson de 2002[10] y Upchurch de 1998[11] y Upchurch et al. de 2004.[12] Todos los análisis dieron como resultado una filogenética similar, donde Europasaurus colocó más derivados que Camarasaurus pero fuera de un clado de Brachiosauridae y Titanosauromorpha, ahora llamado Titanosauriformes.

Durante una descripción de las vértebras del Europasaurus por Carballido & Sander en 2013, se realizó otro análisis filogenético. La matriz cladística se expandió para incluir más taxones de saurópodos, como Bellusaurus, Cedarosaurus y Tapuiasaurus. El taxón Brachiosaurus también se dividió en Brachiosaurus altithorax ) y Giraffatitan brancai, basado en Taylor de 2009.[13] Basado en este análisis más nuevo y más expansivo, Europasaurus se encontró que estaba en una posición similar, como un camarasauromorfo basal más cercano a los titanosaurios que el Camarasaurus. Sin embargo, Euhelopus, Tehuelchesaurus, Tastavinsaurus y Galvesaurus se colocaron entre Europasaurus y Brachiosauridae.[5]
En un análisis de 2012 de la filogenia de Titanosauriformes, D'Emic consideró que Europasaurus pertenecía a Brachiosauridae, en lugar de ser basal a los braquiosáuridos más tempranos. La filogenia resolvió los braquiosáuridos más verdaderos hasta la fecha, aunque se determinó que varios braquiosáuridos potenciales pertenecían a Somphospondyli como Paluxysaurus, Sauroposeidon y Qiaowanlong. Sin embargo, D'Emic fue tentativo al considerar a Europasaurus como un braquiosurido confirmado. Si bien hubo un fuerte apoyo en la filogenia para su colocación, Europasaurus, uno de los pocos macronarios basales con un cráneo, carece de múltiples huesos que muestran rasgos característicos del grupo, como vértebras caudales.[14]
Un análisis posterior sobre titanosauriformes coincidió con D'Emic en la colocación de Europasaurus. Formó una politomía con Brachiosaurus y el "Bothriospondylus francés " ahora llamado Vouivria, como los braquiosáuridos más basales. El más derivado en el clado fue Lusotitan, con Giraffatitan, Abydosaurus , Cedarosaurus y Venenosaurus formando un clado de braquiosáuridos más derivado. Se encontró que los dientes "torcidos" de Europasaurus eran una de las características únicas de Brachiosauridae, lo que podría significar una referencia segura de dientes de saurópodos aislados al clado.[15]
Se realizó un análisis filogenético adicional en Brachiosauridae, basado en el de D'Emic de 2012. Esta filogenia, llevada a cabo por D'Emic et al. en 2016, resolvieron una colocación muy similar de Europasaurus dentro de Brachiosauridae, aunque Sonorasaurus se colocó en un clado con Giraffatitan y Lusotitan se colocó en una politomía con Abydosaurus y Cedarosaurus. El árbol restante fue el mismo que en D'Emic en 2012, aunque Brachiosaurus se colapsó en una politomía con más braquiosáuridos derivados.[16] Otra filogenia de Mannion et al. en 2017 encontraron resultados similares a D'Emic de 2012 y D'Emic et al. de 2016. Europasaurus fue el braquiosáuridos basal, con el "Bothriospondylus francés", o Vouivria , como el siguiente braquiosáurido más basal. Brachiosaurus se colocó fuera de un poltomia de todos los otros braquiosauridos, Giraffatitan , Abydosaurus , Sonorasaurus , Cedarosaurus y Venenosaurus.[17] Una filogenia de 2017, la de Royo-Torres et al., resolvió relaciones más complejas dentro de Brachiosauridae. Además de Europasaurus como el braquiosáurido basal, había dos subgrupos dentro del clado, uno que contenía Giraffatitan, Sonorasaurus y Lusotitan y otro que incluía casi todos los demás braquiosáuridos, así como Tastavinsaurus . Este segundo clado se denominaría Laurasiformes según la definición del grupo. Brachiosaurus estaba en una politomía con los dos clados de Brachiosauridae. La filogenia de Royo-Torres et al. (2017) se puede ver arriba, en la columna derecha.[18]
Filogenia
Cladograma basado en el análisis filogenético de Sanders et al. de 2006.[2]
| Sauropoda |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

El cladograma a continuación a la izquierda ilustra los resultados filogenéticos de D'Emic de 2012, con Euhelopodidae y Titanosauria colapsados.[14]
| Macronaria |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Cladograma basado en el análisis filogenético de Carballido et al. de 2013.[5]
| Macronaria |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Filogenia de Royo-Torres et al. de 2017.[18]

| Macronaria |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Paleobiología
Crecimiento
Se identificó que Europasaurus era una especie enana única, y no un juvenil de un taxón existente como Camarasaurus, por un análisis histológico de múltiples especímenes de Europasaurus. En este análisis se demostró que la muestra más joven, DFMMh/FV 009, carece de signos de envejecimiento, como marcas de crecimiento o tejido óseo laminar, y también es el espécimen más pequeño a 1,75 metros de longitud. Dicho tejido óseo es un indicador de rápido crecimiento, por lo que la muestra es probablemente una cría joven. Una muestra más grande, DFMMh/FV 291.9 a 2 metros muestra grandes cantidades de tejido laminar, sin marcas de crecimiento presentes, por lo que es probable que también sea un juvenil. El siguiente espécimen más pequeño, DFMMh/FV 001 muestra la presencia de marcas de crecimiento, específicamente anillos de crecimiento, y en la longitud de 3,5 metros es posiblemente un subadulto. Más grande, el DFMMh/FV 495 muestra las osteonas maduras y anillo , y mide 3,7 metros. La segunda muestra analizada más grande, DFMMh/FV 153 también muestra marcas de crecimiento, pero son más frecuentes. Este espécimen mide 4,2 metros. Un solo fémur parcial representa el mayor tamaño conocido de Europasaurus, con una longitud corporal de 6,2 metros. A diferencia de todos los demás especímenes, este, DFMMh/FV 415 muestra la presencia de líneas de crecimiento detenido , lo que indica que murió después de alcanzar el tamaño corporal completo. El hueso interno también es parcialmente laminar, lo que demuestra que había dejado de crecer recientemente.[1][2]
Estas combinaciones de factores de crecimiento muestran que Europasaurus desarrolló su pequeño tamaño debido a una tasa de crecimiento muy reducida, ganando tamaño más lentamente que los taxones más grandes como el Camarasaurus. Esta desaceleración en la tasa de crecimiento es lo opuesto a la tendencia general vista en saurópodos y terópodos, que alcanzaron mayores tamaños con mayores tasas de crecimiento.[1] Algunos de los parientes cercanos de Europasaurus representan los dinosaurios más grandes conocidos, incluyendo Brachiosaurus y Sauroposeidon . Marpmann et al.en 2014 propusieron que el pequeño tamaño y la reducida tasa de crecimiento de Europasaurus era un efecto de pedomorfismo, donde los adultos de los taxones conservan características juveniles, como el tamaño.[4]
Enanismo

Se ha sugerido que un antepasado de Europasaurus habría disminuido rápidamente en tamaño corporal después de emigrar a una isla que existía en ese momento, ya que la mayor de las islas en la región del norte de Alemania era más pequeña que 200,000 kilómetros cuadrados, que puede no haber sido capaz de apoyar a una comunidad de grandes saurópodos. Alternativamente, un macronario puede haberse encogido al mismo tiempo que una masa de tierra más grande se hacia más pequeña, hasta alcanzar el tamaño de Europasaurus.[1] Los estudios previos sobre el enanismo insular se limitan en gran medida al Maastrichtiano de la isla Haţeg en Rumania, hogar del enano titanosáurido Magyarosaurus y el también enano hadrosáurido Telmatosaurus.[1][19][20] Se sabe que Telmatosaurus es de un adulto pequeño, y aunque es muy pequeño, se sabe que los ejemplares de Magyarosaurus de tamaños pequeños son de individuos adultos a viejos.[20][3] Los adultos de Magyarosaurus dacus tenían un tamaño corporal similar al Europasaurus, pero el mayor de estos últimos tenía fémures más largos que los más grandes, mientras que el Magyarosaurus hungaricus era significativamente más grande que cualquiera de los taxones.[3] El enanismo en Europasaurus representa el único cambio rápido y significativo de masa corporal en un sauropodomorfo derivado, con la tendencia general de los taxones siendo un crecimiento en el tamaño total en otros grupos.[21]
Paleoecología
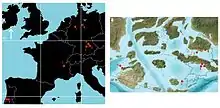
La localidad de Langenberg en Alemania, desde el principio de Oxfordiense hasta finales del Kimmeridgiense, muestra la variedad de vida animal y vegetal de un ecosistema de isla del Jurásico tardío.[22] Durante el Kimmeridgiense la localidad habría sido marino, que se encuentra entre los macizos Rhenisch, Bohemia, y Londres-Brabant. Esto no indica que los taxones presentes fueran marinos, ya que los animales y las plantas pueden haber sido depositados de forma alóctona, aunque solo por una corta distancia, de las islas circundantes.[23] Los sedimentos para mostrar que hubo una afluencia ocasional de agua dulce o salobre y los fósiles conservados muestran eso. Hay un gran número de bivalvos marinos fósiles, así como los equinodermos y microfósiles presentes en la piedra caliza de la cantera, aunque muchos de los animales y plantas eran terrestres.[24]
Muchos taxones marinos se conservan en Langenberg, aunque no habrían coexistido a menudo con el Europasaurus. Hay al menos tres géneros de tortugas, Plesiochelys , Thalassemys y hasta dos taxones sin nombre.[22][25][26] Los peces actinopterygios son abundantes, representados por Lepidotes, Macromesodon, Proscinetes, Coelodus, Macrosemius, Notagogus, Histionotus, Ionoscopus, Callopterus, Caturus, Sauropsis, Belonostomus y Thrissops.[27] También están presentes al menos cinco morfologías distintas de los tiburones Hybodontiformes, los neoselachii Palaeoscyllium , Synechodus y Asterodermus.[28] Se conocen dos cocodrilos marinos de Langenberg, Machimosaurus y Steneosaurus , que probablemente se alimentaron de tortugas y peces, y también se ha encontrado el cocodrilo anfibio Goniopholis.[29]
Los conos y ramitas de coníferas pueden identificarse como la araucaria Brachyphyllum, de los fósiles terrestres del sitio. En la localidad están depositados muchos taxones, incluida una gran acumulación de huesos e individuos de Europasaurus. Se recuperaron al menos 450 huesos de Europasaurus de la cantera de Langenberg, con aproximadamente 1/3 de marcas de dientes.[22] De estas marcas de dientes, los tamaños y formas coinciden bien con los de los peces, cocodrilos u otros carroñeros, pero no hay marcas de terópodos confirmadas.[22][30] El alto número de individuos presentes sugiere que una manada de Europasaurus cruzaba una zona de marea y se ahogó.[22] Si bien el animal dominante de gran cuerpo presente es Europasaurus, también hay material de un saurópodo diplodócido, un estegosáurido y múltiples terópodos. Se conservan tres cervicales del diplodócido y debido a su tamaño, es posible que el taxón también fuera un enano. El estegosáurido y la variedad de terópodos solo conservan los dientes, con la excepción de unos pocos huesos posiblemente de un taxón Ceratosauridae.[22] Los dientes aislados muestran que había al menos cuatro tipos diferentes de terópodos presentes en la localidad, incluido el megalosáurido Torvosaurus sp. así como un megalosáurido adicional y miembros indeterminados de la Allosauridae y Ceratosauria y también hay los dientes más antiguos conocidos de Velociraptorinae .[31][32]
Además de los dinosaurios, muchos vertebrados terrestres de cuerpo pequeño también se conservan en la cantera de Langenberg. Tales animales incluyen un esqueleto de pterosaurio tridimensional bien conservado de Dsungaripteridae y restos aislados de Ornithocheiroidea y Ctenochasmatidae, un lagarto Paramacellodidae y esqueletos parciales y cráneos de un pariente de Theriosuchus ahora llamado como el género Knoetschkesuchus.[22][23][25] Los dientes de mamíferos dryolestid también se conservan, así como un docodont , un taxón en Eobaataridae , unmultituberculado con similitudes con Proalbionbataar, ahora llamado Teutonodon.[22][33]
Extinción
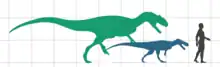
Las huellas de dinosaurios conservadas en la Cantera Langenberg muestran una posible razón para la extinción del Europasaurus y potencialmente, otros enanos insulares presentes en las islas de la región. Las huellas se ubican a 5 metros sobre el depósito de individuos de Europasaurus lo que muestra que al menos 35000 años después de ese depósito hubo una caída en el nivel del mar que permitió un vuelco de la fauna. Los terópodos que habitan en la isla, que coexistían con Europasaurus, habrían medido de aproximadamente 4 metros, pero los terópodos que llegaron sobre el puente de tierra conservaron las huellas de hasta 54 centímetros, lo que indica un tamaño de cuerpo entre 7 y 8 metros si se reconstruyen como un Allosaurus. Fue sugerido por los descriptores de estas huellas, Jens Lallensack y colegas, que estos taxones terópodos probablemente extinguieron la fauna enana especializada, y la cama a partir de la cual se originaron las huellas, Langenberg cama 92, es probablemente la más joven en la que está presente Europasaurus.[8]
Referencias
- Sander, P.M.; Mateus, O.V.; Laven, T.; Knötschke, N. (2006). «Bone histology indicates insular dwarfism in a new Late Jurassic sauropod dinosaur». Nature 441 (7094): 739-741. Bibcode:2006Natur.441..739M. PMID 16760975. doi:10.1038/nature04633.
- Sander, P.M.; Mateus, O.V.; Laven, T.; Knötschke, N. (2006). «Supplementary Information for: Bone histology indicates insular dwarfism in a new Late Jurassic sauropod dinosaur». Nature 441: 739-741. Bibcode:2006Natur.441..739M. PMID 16760975. doi:10.1038/nature04633.
- Stein, K.; Csiki, Z.; Curry-Rogers, K.; Weishampel, D.B.; Redelstorff, R.; Carballido, J.L.; Sander, P.M. (2010). «Small body size and extreme cortical bone remodeling indicate phyletic dwarfism in Magyarosaurus dacus (Sauropoda: Titanosauria)». Proceedings of the National Academy of Sciences 107 (20): 9258-9263. Bibcode:2010PNAS..107.9258S. ISSN 0027-8424. PMC 2889090. PMID 20435913. doi:10.1073/pnas.1000781107.
- Marpmann, J.S.; Carballido, J.L.; Sander, P.M.; Knötschke, N. (2014). «Cranial anatomy of the Late Jurassic dwarf sauropod Europasaurus holgeri (Dinosauria, Camarasauromorpha): Ontogenetic changes and size dimorphism». Journal of Systematic Palaeontology 13 (3): 1-43. doi:10.1080/14772019.2013.875074.
- Carballido, J.L.; Sander, M.P. (2013). «Postcranial axial skeleton of Europasaurus holgeri (Dinosauria, Sauropoda) from the Upper Jurassic of Germany: implications for sauropod ontogeny and phylogenetic relationships of basal Macronaria». Journal of Systematic Palaeontology 12 (3): 335-387. ISSN 1477-2019. doi:10.1080/14772019.2013.764935.
- Upchurch, P.; Mannion, P.D.; Taylor, M.P. (2015). «The Anatomy and Phylogenetic Relationships of "Pelorosaurus" becklesii (Neosauropoda, Macronaria) from the Early Cretaceous of England». PLOS One 10 (6): e0125819. ISSN 1932-6203. PMC 4454574. PMID 26039587. doi:10.1371/journal.pone.0125819.
- Giménez, O. del V. (2007). «Skin impressions of Tehuelchesaurus (Sauropoda) from the Upper Jurassic of Patagonia». Revista del Museo Argentino de Ciencias Naturales 9 (2): 119-124. doi:10.22179/revmacn.9.303. Archivado desde el original el 7 de octubre de 2012. Consultado el 12 de noviembre de 2018.
- Lallensack, J.N.; Sander, M.P.; Knötschke, N.; Wings, O. (2015). «Dinosaur tracks from the Langenberg Quarry (Late Jurassic, Germany) reconstructed with historical photogrammetry: Evidence for large theropods soon after insular dwarfism». Palaeontologia Electronica 18 (2): 1-34.
- Wings, O.; Knötschke, N. «A song of blasting and fire: Europasaurus holgeri». 74th Annual Meeting of the Society of Vertebrate Paleontology: Program and Abstracts.
- Wilson, J.A. (2002). «Sauropod dinosaur phylogeny: critique and cladistic analysis». Zoological Journal of the Linnean Society 136 (2): 217-276. doi:10.1046/j.1096-3642.2002.00029.x.
- Upchurch, P.M. (1998). «The phylogenetic relationships of sauropod dinosaurs». Zoological Journal of the Linnean Society 124: 43-103. doi:10.1111/j.1096-3642.1998.tb00569.x.
- Upchurch, P.M.; Barrett, P.M.; Dodson, P. (2004). «Sauropoda». En Weishampel, D.B.; Dodson, P.; Osmólska, H., eds. The Dinosauria (2 edición). Berkeley: University of California Press. pp. 259–322. ISBN 978-0-520-94143-4.
- Taylor, M.P. (2009). «A re-evaluation of Brachiosaurus altithorax Riggs 1903 (Dinosauria, Sauropoda) and its generic separation from Giraffatitan brancai (Janensch 1914).». Journal of Vertebrate Paleontology 29: 787-806. doi:10.1671/039.029.0309.
- D'Emic, M.D. (2012). «The early evolution of titanosauriform sauropod dinosaurs». Zoological Journal of the Linnean Society 166 (3): 624-671. doi:10.1111/j.1096-3642.2012.00853.x.
- Mannion, P.D.; Upchurch, P.; Barnes, R.N.; Mateus, O. (2013). «Osteology of the Late Jurassic Portuguese sauropod dinosaur Lusotitan atalaiensis (Macronaria) and the evolutionary history of basal titanosauriforms». Zoological Journal of the Linnean Society 168 (1): 98-206. ISSN 0024-4082. doi:10.1111/zoj.12029.
- D’Emic, M.D.; Foreman, B.Z.; Jud, N.A. (2016). «Anatomy, systematics, paleoenvironment, growth, and age of the sauropod dinosaur Sonorasaurus thompsoni from the Cretaceous of Arizona, USA». Journal of Paleontology 90 (1): 102-132. ISSN 0022-3360. doi:10.1017/jpa.2015.67.
- Mannion, P.D.; Allain, R.; Moine, O. (2017). «The earliest known titanosauriform sauropod dinosaur and the evolution of Brachiosauridae». PeerJ 5: e3217. ISSN 2167-8359. PMC 5417094. PMID 28480136. doi:10.7717/peerj.3217.
- Royo-Torres, R.; Fuentes, C.; Meijide, M.; Meijide-Fuentes, F.; Meijide-Fuentes, M. (2017). «A new Brachiosauridae Sauropod dinosaur from the lower Cretaceous of Europe (Soria Province, Spain)». Cretaceous Research 80: 38-55. doi:10.1016/j.cretres.2017.08.012.
- Nopcsa, F. (1914). «Über das Vorkommen der Dinosaurier in Siebenbürgen». Ver. Zool. Bot. Ges. Wien. 54: 12-14.
- Weishampel, D.B.; Norman, D.B.; Grigorescu, D. (1993). «Telmatosaurus transsylvanicus from the Late Cretaceous of Romania: the most basal hadrosaurid dinosaur». Palaeontology 36: 361-385.
- Benson, R.B.J.; Campione, N.E.; Carrano, M.T.; Mannion, P.D.; Sullivan, C.; Upchurch, P.; Evans, D.C. (2014). «Rates of Dinosaur Body Mass Evolution Indicate 170 Million Years of Sustained Ecological Innovation on the Avian Stem Lineage». PLoS Biology 12 (5): e1001853. PMC 4011683. PMID 24802911. doi:10.1371/journal.pbio.1001853.
- Wings, O. (2015). «The Langenberg Quarry near Goslar: Unique window into a terrestrial Late Jurassic ecosystem in Northern Germany». Abstracts of the 12th Symposium on Mesozoic Terrestrial Ecosystems: 99-100.
- Schwarz, D.; Raddatz, M.; Wings, O. (2017). «Knoetschkesuchus langenbergensis gen. nov. sp. nov., a new atoposaurid crocodyliform from the Upper Jurassic Langenberg Quarry (Lower Saxony, northwestern Germany), and its relationships to Theriosuchus». PLOS One 12 (2): e0160617. Bibcode:2017PLoSO..1260617S. doi:10.1371/journal.pone.0160617.
- Wings, O. (2013). «Unique taphonomy of the dwarfed sauropod Europasaurus from Late Jurassic marine strata of the Langenberg Quarry (Lower Saxony, Northern Germany)». Journal of Vertebrate Paleontology, Program and Abstracts. 73rd Annual Meeting of the Society of Vertebrate Paleontology 2013: 239.
- Wings, O.; Sander, M.P. (2012). «The Late Jurassic Vertebrate Assemblage of the Langenberg Quarry, Oker, Northern Germany». 10th Annual Meeting of the European Association of Vertebrate Paleontologists 20: 281-284.
- Marinheiro, J.; Mateus, O. (2011). «Occurrence of the marine turtle Thalassemys in the Kimmeridgian of Oker, Germany». 71st Annual Meeting of the Society of Vertebrate Paleontology. Abstracts of the 71st Annual Meeting of the Society of Vertebrate Paleontology: 151.
- Mudroch, A.; Thies, D. (1996). «Knochenfischzähne (Osteichthyeso Actinopterygii) aus dem Oberjura (Kimmeridgium) des Langenbergs bei Oker (Norddeutschland)». Geologica et Palaeontologica (en alemán) 30: 239-265.
- Thies, D. (1995). «Placoid scales (Chondrichthyes: Elasmobranchii) from the Late Jurassic (Kimmeridgian) of northern Germany». Journal of Vertebrate Paleontology 15 (3): 463-481. doi:10.1080/02724634.1995.10011242.
- Karl, H.-V.; Gröning, E.; Brauckmann, C.; Schwarz, D.; Knötschke, N. (2006). «The Late Jurassic crocodiles of the Langenberg near Oker, Lower Saxony (Germany), and description of related materials (with remarks on the history of quarrying the "Langenberg Limestone" and "Obernkirchen Sandstone")». Clausthaler Geowissenschaften 5: 59-77.
- Slodownik, M.; Wings, O. (2015). «Bite marks on Europasaurus bones from the Langenberg Quarry near Goslar (Lower Saxony, Germany)». 13th Annual Meeting of the European Association of Vertebrate Palaeontologists Opole, Poland, Abstracts: 141.
- Gerke, O.; Wings, O. (2016). «Multivariate and Cladistic Analyses of Isolated Teeth Reveal Sympatry of Theropod Dinosaurs in the Late Jurassic of Northern Germany». PLOS One 11 (7): e0158334. Bibcode:2016PLoSO..1158334G. doi:10.1371/journal.pone.0158334.
- Lubbe, T. van Der; Richter, U.; Knötschke, N. (2009). «Velociraptorine Dromaeosaurid Teeth from the Kimmeridgian (Late Jurassic) of Germany». Acta Palaeontologica Polonica 54 (3): 401-408. doi:10.4202/app.2008.0007.
- Martin, T.; Schultz, J.A.; Schwermann, A.H.; Wings, O. (2016). «First Jurassic mammals of Germany: Multituberculate teeth from Langenberg Quarry (Lower Saxony)». Palaeontologia Polonica 67: 171-179. doi:10.4202/pp.2016.67_171. Archivado desde el original el 10 de agosto de 2016. Consultado el 12 de noviembre de 2018.
Véase también
Enlaces externos
 Portal:Dinosaurios. Contenido relacionado con Dinosaurios.
Portal:Dinosaurios. Contenido relacionado con Dinosaurios. Wikimedia Commons alberga una categoría multimedia sobre Europasaurus holgeri.
Wikimedia Commons alberga una categoría multimedia sobre Europasaurus holgeri. Wikispecies tiene un artículo sobre Europasaurus holgeri.
Wikispecies tiene un artículo sobre Europasaurus holgeri.- El PaleoFreak - Europasaurus: más enanismo insular
- Palaeoblog - Europasaurus holgeri: Germany's Pygmy Dinosaur (en inglés)
| Control de autoridades |
|
|---|
 Datos: Q120970599
Datos: Q120970599