Brachiosaurus altithorax
Brachiosaurus altithorax ("lagarto brazo de tórax alto"), castellanizado como braquiosaurio, es la única especie conocida del género extinto Brachiosaurus de dinosaurio saurópodo braquiosáurido, que vivió durante el Jurásico Superior, hace aproximadamente 154 a 153 millones de años, en lo que es hoy Norteamérica. Fue descrita por primera vez por Elmer S. Riggs en 1903 a partir de fósiles encontrados en el río del Gran Cañón, ahora Río Colorado del oeste de Colorado, en sedimentos de la famosa Formación Morrison, Estados Unidos. Brachiosaurus tiene el cuello extremadamente largo, un cráneo pequeño y gran tamaño general, todos los cuales son rasgos típicos de los saurópodos. Sin embargo, Brachiosaurus tiene proporciones que son diferentes de la mayoría de los saurópodos, siendo las patas delanteras más largas que las patas traseras, característica a la que hace referencia su nombre, y su cola era más corta en proporción a su cuello que otros saurópodos del Jurásico.
| Brachiosaurus altithorax | ||
|---|---|---|
| Rango temporal: 154 Ma - 153 Ma Jurásico Superior (Kimmeridgiense) | ||
 | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Sauropsida | |
| Superorden: | Dinosauria | |
| Orden: | Saurischia | |
| Suborden: | Sauropodomorpha | |
| Infraorden: | Sauropoda | |
| Superfamilia: | Titanosauriformes | |
| Familia: | Brachiosauridae | |
| Género: |
Brachiosaurus Riggs, 1903 | |
| Especie: |
B. altithorax Riggs, 1903 | |
| Sinonimia | ||
| ||
Brachiosaurus es el género que da nombre a la familia Brachiosauridae, que incluye a otros saurópodos similares. Gran parte de lo que se conoce sobre Brachiosaurus se basa de hecho en Giraffatitan brancai, una especie de dinosaurio braquiosáurido de la Formación Tendaguru de Tanzania que fue descrito originalmente por el paleontólogo alemán Werner Janensch como una especie de Brachiosaurus en 1914. Investigaciones posteriores muestran que las diferencias entre la especie tipo de Brachiosaurus y el material Tendaguru son lo suficientemente significativas para que el material de África deba ser colocado en un género propio. Varias otras especies potenciales de Brachiosaurus se han descrito desde África y Europa, pero se cree que ninguno de ellos pertenecía a Brachiosaurus.
Brachiosaurus es uno de los saurópodos más raros de la Formación Morrison. El espécimen tipo de B. altithorax sigue siendo el ejemplar más completo, y se piensa que solo debe haber un puñado de otros especímenes que pertenezcan al género. Brachiosaurus es considerado como un alto ramoneador, que probablemente hubiese mordisqueado o pellizcado la vegetación de la copa de los árboles posiblemente tan alto como 9 metros por encima del suelo. A diferencia de otros saurópodos, la representación de pararse sobre sus patas traseras en la película Parque Jurásico, es inadecuada. Brachiosaurus se ha utilizado como un ejemplo de un dinosaurio que era probablemente ectotérmico debido a su gran tamaño y su necesidad de alimentación, pero investigaciones más recientes han demostrado que habría sido de sangre caliente[cita requerida].

Descripción
Al igual que todos los dinosaurios saurópodos, Brachiosaurus era un animal cuadrúpedo con un cráneo pequeño, cuello largo, una cola relativamente corta, musculosa y delgada, extremidades columnares.[1] El cráneo tenía un hocico ancho y robusto y con una gran mandíbula, con dientes en forma de espátula. Al igual que Giraffatitan, Brachiosaurus tenía un arco de hueso sobre el hocico y en frente de los ojos que redondeaban la fenestra nasal, aunque este arco no era tan grande como en su pariente.[2] Tenía grandes sacos de aire conectados al sistema pulmonar que estaban presentes en el cuello y el tronco, incluyendo las vértebras y costillas, reduciendo en gran medida el peso.[3][4] Inusualmente para un saurópodo, las extremidades anteriores eran más largas que las extremidades traseras. El húmero,hueso del brazo, de Brachiosaurus es relativamente ligero por su tamaño,[5] que mide 2,04 metros de longitud en el espécimen tipo.[6] El fémur, hueso del muslo, del espécimen tipo mide solamente 2,03 metros de largo.[6] A diferencia de otros saurópodos, Brachiosaurus parece haber tenido ligeramente tumbada la articulación del hombro,[5] y la caja torácica era inusualmente profunda.[6] Esto hacía que el tronco estuviese inclinado, con la parte delantera mucho más alta que las caderas, además el cuello sale del tronco en un ángulo pronunciado. En general, esta forma se asemeja a una jirafa más que cualquier otro animal vivo.[7]
Tamaño y masa
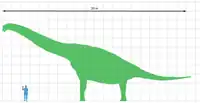
Debido a que se conoce material mucho más completo de Giraffatitan "Brachiosaurus" brancai que de B. altithorax, la mayoría de las estimaciones del tamaño de Brachiosaurus son en realidad de la forma africana. Hay un elemento adicional de incertidumbre para el Brachiosaurus de Norteamérica debido a que el esqueleto más completo que se conoce puede ser de un ejemplar subadulto.[5] Con los años, la masa de B. altithorax se ha estimado en 35,0 toneladas por Paul en 1988,[7] 43,9 toneladas por Foster en 2003,[8] 28,7 toneladas por Taylor en 2009[5] y, más recientemente, en 56,3 toneladas métricas por Benson en 2014.[9] Están los casos en que también los autores proporcionaron estimaciones para Giraffatitan, y encontraron que el género es un poco más ligero 31,5 toneladas por Paul en 1988,[7] 39,5 toneladas por Mazzetta en 2004,[10] 23.3 toneladas por Taylor en 2009,[5] y 34 toneladas métricas por Benson en 2014.[9] La longitud del Brachiosaurus se ha estimado en 26 metros.[11]
Postura del cuello
En contraste con la mayoría de otros saurópodos, los braquiosáuridos tenían la espalda inclinada hacia la zona caudal, debido a sus largas patas delanteras. Por lo tanto, si el cuello salía del cuerpo en línea recta, este apuntaría hacia arriba.[5][12][13][14] El ángulo exacto depende de cómo se reconstruya la cintura escapular, y debido a esto los omóplatos son colocados en la caja torácica.[12][13][14] La movilidad del cuello de Brachiosaurus ha sido interpretada como considerablemente reducida por Stevens y Parrish,[12][13][14] mientras que otros investigadores como Paul, Christian y Dzemski han abogado por la idea de un cuello más flexible.[7][15]
Descubrimiento e investigación

El género Brachiosaurus, y la especie tipo B. altithorax, se basan en un esqueleto parcial postcraneal de Fruita, en el valle del río Colorado en el oeste de Colorado.[16] Este espécimen fue recogido de las rocas de la cuenca Brushy de la Formación Morrison[17] en 1900 por Elmer S. Riggs y su equipo por una expedición del Museo Field Columbian, ahora Museo Field de Historia Natural, de Chicago.[6] El espécimen está catalogado actualmente como FMNH P25107.[5] Riggs y su equipo estaban trabajando en la zona como consecuencia de la correspondencia favorable entre Riggs y S.M. Bradbury, un dentista en las inmediaciones de Grand Junction, que también era coleccionista aficionado de fósiles. En 1899, Riggs había preguntado a las localidades rurales en el oeste de los Estados Unidos en relación con hallazgos fósiles, y Bradbury, informó que se habían recogido huesos de dinosaurios en la zona desde 1885.[16] Riggs y su asistente de campo H. W. Menke encontraron el espécimen FMNH P 25107,[6] el 4 de julio de 1900[18] en la Cantera 13, en una pequeña colina más tarde conocida como la colina Riggs, la cual está marcada con una placa. Se han reportado descubrimientos de fósiles adicionales de Brachiosaurus en la colina Riggs, pero otros fósiles encontrados en la colina han sido objeto de vandalismo.[18][19] Riggs publicó un informe breve en 1901, indicando la longitud inusual del húmero en comparación con el fémur, el tamaño y las proporciones similares a una jirafa, así como el pequeño tamaño de la cola, pero no publicó un nombre para el nuevo dinosaurio.[20] Las publicaciones de Riggs de 1901 y 1903 sugirieron erróneamente que el espécimen fue el más grande dinosaurio conocido.[6][20] Riggs en su publicación de 1903 nombró a Brachiosaurus altithorax[6] e hizo una descripción más detallada en una monografía en 1904.[21] El nombre del género se deriva de los términos griegos brachion/βραχιων que significa "brazo" y sauros/σαυρος que significa "lagarto", porque Riggs se dio cuenta de que la longitud de los brazos era inusual para un saurópodo.[6] El epíteto "altithorax" fue elegido debido a que la cavidad torácica es inusualmente ancha y profunda, y esta proviene de las palabras en latín altus que significa "profundo", y la griega thorax/θώραξ, que significa "peto, coraza, tórax".[22]

El esqueleto encontrado en Fruita no fue el primer descubrimiento de Brachiosaurus, a pesar de que fue el primero en ser reconocido como un nuevo género de saurópodo, el primer verdadero descubrimiento fue en 1883, cuando el cráneo de un saurópodo fue encontrado cerca de Garden Park, Colorado, en la Cantera Felch 1, y fue enviado a Othniel Charles Marsh durante la famosa "guerra de los huesos".[5] Después Marsh incorporó el cráneo a su restauración del esqueleto de Brontosaurus.[5][23] Con el tiempo "ese cráneo" el espécimen USNM 5730 pasó a formar parte de las colecciones del Museo Nacional de Historia Natural.[5] En la década de 1970 cuando Jack McIntosh y David Berman estaban trabajando en el tema del verdadero cráneo del Apatosaurus, revaluaron el cráneo de Garden Park y determinaron que este era más similar al de un Camarasaurus.[24] Y fue descrito y reconocido como un espécimen de Brachiosaurus en 1998 por Kenneth Carpenter y Virginia Tidwell, como una forma intermedia entre Camarasaurus y Giraffatitan, cuando todavía se considera como B. brancai.[25] Debido a que no hay fósiles que coincidan entre este cráneo y el espécimen FMNH P 25107, no se puede determinar con confianza la asignación a la especie,[5][25] por lo que se clasifica como Brachiosaurus sp.[25]
La preparación de FMNH P25107, el holotipo de Brachiosaurus comenzó en el otoño de 1900 poco después de que fue recogido por Elmer Riggs para el Museo Field de Historia Natural, Chicago. Cuando la preparación de cada hueso fue terminada, fue puesta en exhibición en una vitrina en el pabellón 35 del Palacio de Bellas Artes de la Exposición Mundial Colombina de Chicago, primera casa del Museo Field. Todos los huesos estaban en la exhibición antes de 1908 cuando el Apatosaurus recientemente montado del Museo Field fue revelado. Sin embargo, no se intentó ningún montaje debido a que solo se había sido recuperado el 20% del esqueleto. En 1993, los huesos del holotipo fueron moldeados y copiados, y los huesos que faltaban fueron esculpidos basados en el material del Giraffatitan en Berlín. Este esqueleto de plástico fue montado y, en 1994, se puso en exhibición en el extremo norte del Stanley Field Hall, la principal sala de exposiciones del actual edificio del Museo Field. Los huesos reales del holotipo se expusieron en dos grandes cajas de vidrio en cada extremo del montaje. El montaje se mantuvo hasta 1999, cuando fue trasladado al B Concourse de la Terminal One de United Airlines en el Aeropuerto Internacional O'Hare para dejar espacio para el recién adquirido Tyrannosaurus rex del museo, "Sue".[26] Al mismo tiempo, el Museo Field montó un segundo molde plástico del esqueleto, diseñado para el uso exterior y ha estado en la exhibición fuera del museo en la terraza del nanovatio desde entonces. Los únicos huesos verdaderos actualmente en exhibición son el húmero y dos dorsales en la sala del Mesozoico de la Exhibición el Planeta que Evoluciona del Museo Field.[27]
Descubrimientos adicionales de material de Brachiosaurus en Norteamérica han sido infrecuentes y consisten en un puñado de huesos. Se ha descrito material de Colorado,[5][28][29][30] Oklahoma,[5][31] Utah,[5][28] y Wyoming,[5][8] y se ha mencionado material no descrito de varios otros sitios.[5][17] Uno de estos ejemplares, un escapulo-coracoides de la cantera de Dry Mesa, Colorado, es uno de los ejemplares en el centro de la cuestión Supersaurus/Ultrasauros de los años 80 y 90 del siglo XX. En 1985, James A. Jensen describió los restos de un saurópodo desarticulado de la cantera como pertenecientes a varios taxones, incluyendo los nuevos géneros Supersaurus y Ultrasaurus,[32] este último renombrado Ultrasauros poco después porque otro saurópodo ya tenía el nombre.[33] Estudio posterior mostró que el material de "Ultrasauros" pertenecía en su mayoría a Supersaurus, aunque el omóplato no. Debido a que el holotipo de Ultrasauros, una vértebra de la espalda, fue uno de los especímenes que en realidad eran de Supersaurus, el nombre Ultrasauros es un sinónimo de Supersaurus. El omóplato es ahora asignado a Brachiosaurus, pero la especie es incierta.[5][29] Además, el Ultrasauros de Dry Mesa no era tan grande como se había pensado, las dimensiones del hueso coracoides del hombro indican que el animal era más pequeño que el espécimen original de Riggs de Brachiosaurus.[5][29]
Material referido
Taylor en 2009 enumera una serie de especímenes referidos a Brachiosaurus. Estos incluyen distintos materiales, por ejemplo, un húmero de Potter Creek y un poco de material de Dry Mesa, este último parcialmente descrito como Ultrasauros por Jensen, que claramente no son braquiosáuridos en origen, o al menos no claramente referible a Brachiosaurus.[5] Por el contrario, una vértebra cervical y el cráneo mencionado anteriormente pueden pertenecer a B. altithorax o a un braquiosáurido aún desconocido de América del Norte.[5] La cervical fue encontrada cerca Jensen, Utah, por Jensen[28] y si pertenece a Brachiosaurus, es una de un puñado de vértebras del cuello conocidas para braquiosáuridos norteamericanos. No hay ningún material inequívoco en el cráneo, el cuello, la región dorsal anterior, tronco delantero, la porción distal de los miembros o pies.[5] Más recientemente, Carballido et al. en 2012 informaron sobre un esqueleto postcraneal casi completo de un saurópodo juvenil de aproximadamente 2 metros de largo de la Formación Morrison en la Cuenca de Bighorn, al norte-central de Wyoming. Este espécimen se consideró originalmente que pertenecía a un diplodócido, pero los autores lo reinterpretaron como un braquiosáurido, probablemente B. altithorax.
Había un amplio material referido a "B." brancai en las colecciones del Museo de Historia Natural de Berlín, algunas de las cuales fueron destruidas durante la Segunda Guerra Mundial. Otros materiales fueron transferidos a otra institución en toda Alemania, algunos de los cuales también fueron destruidos. Es probable que aparezcan más especímenes entre el material recogido por la expedición Tendaguru del Museo Británico de Historia Natural.[34] Mucho o todo este material pertenece probablemente a Giraffatitan, aunque algunos pueden representar un nuevo braquiosáurido.[35]
Separación de Giraffatitan
Al describir el material del braquiosáurido de Tendaguru en 1914, Janensch enumeró una serie de diferencias y similitudes entre ellos y B. altithorax.[36] En otras tres publicaciones en 1929,[37] 1950[38] y 1961,[39] Janensch comparó las dos especies con más detalle, enumerando 13 características compartidas.[5] De éstas, sin embargo, solo cuatro parecen ser válidas, mientras que seis pertenecen a grupos más inclusivos que Brachiosauridae, y el resto son difíciles de evaluar o se refieren a material que no es Brachiosaurus. En 1988, Gregory Paul publicó una nueva reconstrucción del esqueleto de "B." brancai, destacando una serie de diferencias en proporción entre este y B. altithorax. La principal es una diferencia en la forma en que varían las vértebras del tronco, son bastante uniformes en B. altithorax, pero varían ampliamente en el material africano. Paul creía que los elementos de las extremidades y cinturas de ambas especies eran muy similares, y por lo tanto sugirió separarlos no en el género, sino solo en el nivel del subgénero.[7] Giraffatitan fue elevado a nivel de género por Olshevsky sin comentarios.[33] Un estudio detallado de todo el material, incluyendo los huesos de la extremidad y de la cintura, por Michael Taylor en 2009 encontró que hay diferencias significativas entre B. altithorax y el material de Tendaguru en todos los elementos conocidos de ambas especies. Taylor encontró 26 caracteres osteológicos distintos, una diferencia mayor que la existente entre, por ejemplo, Diplodocus y Barosaurus, y por lo tanto argumentó que el material africano debería ser colocado en su propio género, Giraffatitan, con la especie G. brancai. Una diferencia importante entre los dos géneros es la forma general del cuerpo, con Brachiosaurus teniendo un tronco 23% más largo y una cola 20 a 25% más larga y también más alta.[5]
Clasificación
Brachiosaurus es el género epónimo de Brachiosauridae, familia a la que le da nombre.[21] A través de los años, una serie de saurópodos han sido asignados a Brachiosauridae, como Astrodon, Bothriospondylus, Dinodocus, Pelorosaurus, Pleurocoelus, y Ultrasaurus,[40] pero la mayoría de ellos se consideran actualmente como dudosos o de colocación incierta.[1] Un análisis filogenético de los saurópodos publicados en la descripción de Abydosaurus encontró que el género formaría un clado con Brachiosaurus y Giraffatitan (incluido en Brachiosaurus).[41] Un análisis más reciente que se centró en un posible braquiosáurido asiático encontró que este formaría un clado que incluye Abydosaurus, Brachiosaurus, Cedarosaurus, Giraffatitan, y Paluxysaurus, pero no Qiaowanlong, el saurópodo que se estudiaba.[42] Géneros relacionados incluyen Lusotitan y Sauroposeidon.[1] Brachiosauridae está situada en la base de Titanosauriformes, un grupo de saurópodos, que también incluye a los titanosaurios.[42]

De acuerdo con el diagnóstico revisado por Taylor, Brachiosaurus altithorax se diagnostica como una plétora de rasgos distintivos muchos de los cuales se encuentran en la parte posterior de las vértebras dorsales.[5] Entre los rasgos distintivos que lo colocan en la familia Brachiosauridae son una relación entre la longitud del húmero al fémur longitud de al menos 0,9 (es decir, el hueso del brazo superior es al menos casi tan largo como el hueso del muslo), y un eje del fémur es muy aplanada (relación ≥1.85).[5]
Brachiosaurus altithorax
FMNH 25107, es el holotipo tanto del género Brachiosaurus como de la especie B. altithorax, consiste de un húmero derecho, un fémur derecho, un iliaco derecho, un coracoides derecho, un hueso sacro, las últimas 7 vértebras dorsales, 2 vértebras caudales, y varias costillas.[5][6][20] Riggs describe al coracoides a la izquierda del cuerpo,[6][21][20] pero posteriores estudios lo colocan a la derecha.[5]
En 1969, durante un estudio de Kingham, Brachiosaurus altithorax, junto con especies asignadas ahora a otros géneros, se movieron del género. Kingham encontró que "B." atalaiensis, "B." brancai y B. altithorax son referibles a Astrodon creando muchas nuevas especies de Astrodon aparte del tipo. La visión de Kingham de la taxonomía de los braquiosáuridos no ha sido aceptada por muchos otros autores.[43]
Especies reasignadas
- "B." atalaiensis: Originalmente descrita por de Lapparent y Zbyszewski,[44] la referencia de este material a Brachiosaurus fue puesta en duda por Upchurch, Barret y Dodson,[1] que lo enumeró como un braquiosáurido sin nombre, y colocado en su propio género Lusotitan por Antunes y Mateus.[45] De Lapparent y Zbyszewski describieron una serie de restos pero no designaron un espécimen tipo. Antunes y Mateus seleccionaron un esqueleto postcraneal parcial, MIGM 4978, 4798, 4801-4810, 4938, 4944, 4950, 4952, 4958, 4964-4966, 4981-4982, 4985, 8807, 8793-87934 como lectotipo. Este espécimen incluye 28 vértebras, cheurones, costillas, posible un omóplato, humero, huesos del antebrazo, la pelvis parcial izquierda, huesos de la pierna y parte del tobillo derecho. Las espinas neurales bajas, la cresta deltopectoral prominente del húmero, un sitio de inserción muscular en el hueso del brazo, el húmero alargado muy largo y esbelto y el eje largo del ilion inclinado hacia arriba indican que Lusotitan es un braquiosáurido.[45]
- "B." brancai: Janensch baso su descripción en el "Esqueleto S" de laformación Tendaguru,[36] pero más tarde se dio cuenta de que comprendía dos individuos parciales: S I y S II.[37] Él al principio no los designó como una serie sintipo, pero en 1935 hizo S I, MB.R.2180, el lectotipo. Taylor en 2009, sin darse cuenta de esta acción, propuso el más grande y más completo S II, MB.R.2181, como el lectotipo.[5] Incluye, entre otros huesos, varias vértebras dorsales, la escápula izquierda, ambos coracoides, ambos esternales, ambos húmeros, ambos cúbitos y radios, una mano derecha, una mano izquierda parcial, ambos pubis y el fémur derecho, la tibia y el peroné. Más tarde, Taylor se dio cuenta de que Janensch había designado en 1935 el esqueleto más pequeño S I como el lectotipo.[46][47] Una revaluación de la relación entre el material de braquiosáurido africano y americano indica que un nombre genérico separado está justificado para el material de Tendaguru, por lo que "B. brancai" se ha trasladado a su propio género Giraffatitan.[5][7]
Especies inválidas
- "B." fraasi: erigido por Janensch en 1914, pero luego sinonimizado con "B." brancai;[37] Este material pertenece ahora a Giraffatitan.[5]
- "B." nougaredi: Esta especie se conoce a partir de restos fragmentarios descubiertos en el este de Argelia, en el desierto del Sahara. El material conocido del tipo consiste de un sacro y algo del metacarpo izquierdo y falanges. Se encontraron en el sitio de descubrimiento pero no se recogieron huesos parciales del antebrazo izquierdo, huesos de la muñeca, hueso de la espinilla derecha y fragmentos que pudieron provenir de metatarsianos.[48] De Lapparent, quien describió y nombró el material en 1960, reportó que la localidad de descubrimiento estaba en la serie de Taouratine del Jurásico Superior, asignando las rocas a esta edad en parte debido a la presunta presencia de Brachiosaurus.[48] Este material se encontró desunido en un área de varios cientos de metros,[48] y probablemente no represente una sola especie.[49]
Paleobiología
Alimentación
A Brachiosaurus se lo suele ver como un folívoro alto, alimentándose en el follaje en las copas de los árboles más que sobre la tierra. Incluso si no tenía el cuello cerca de la vertical, y en su lugar tenía un cuello recto, su altura de la cabeza puede haber estado a más de 9 metros sobre el suelo.[2][8] Se alimentó probablemente sobre todo en el follaje sobre los 5 metros. Esto no excluye la posibilidad de que también se alimente más bajo a veces, entre 3 a 5 metros de altura.[2] Su dieta consistía probablemente en ginkgos, coníferas, helechos arborescentes y cicadáceas grandes, con un consumo estimado en 200 a 400 kilogramos de materia vegetal diariamente.[2] Sin embargo, Brachiosaurus pudo haber requerido solamente cerca de 120 kilogramos de forraje por día. La alimentación del Brachiosaurus implicaba un simple movimiento de la mandíbula hacia arriba y hacia abajo. Los dientes se dispusieron para cortar el material a medida que se cerraban, y probablemente se usaron para rastrillar y/o cortar la vegetación.[50]
Se ha sugerido repetidamente, por ejemplo, en la película Jurassic Park, que Brachiosaurus podría adoptar una posición bípeda o trípode, con soporte de cola, para alimentarse.[7] Sin embargo, un análisis basado en el modelado físico detallado de las capacidades de los saurópodos de Heinrich Mallison mostró que mientras muchos saurópodos podrían levantarse sobre sus patas traseras, la relación de la forma del cuerpo del Brachiosaurus y la longitud del miembro anterior excepcionalmente larga habría hecho poco probable el poder hacerlo. La posición delantera del centro de masa habría conducido a problemas con la estabilidad y haber requerido fuerzas excesivamente grandes en las caderas para obtener una postura erguida. Brachiosaurus también habría ganado relativamente poco con levantarse, sólo el 33% más de altura de alimentación, en comparación con otros saurópodos, para los cuales una postura bípeda puede haber triplicado la altura de alimentación.[51]
Metabolismo
Al igual que todos los saurópodos, Brachiosaurus fue homotermo, manteniendo una temperatura interna estable y endotérmico controlando la temperatura del cuerpo a través de los medios internos, lo que significa que era capaz de controlar activamente su temperatura corporal, de "de sangre caliente", produciendo el calor necesario a partir de una alta tasa metabólica basal de sus células.[52] En el pasado, Brachiosaurus se ha utilizado un ejemplo de un dinosaurio para el cual la endotermia era improbable, debido a la combinación del gran tamaño, que lleva al sobrecalentamiento y de las grandes necesidades calóricas para mantener alto el metabolismo.[53] Sin embargo, estos cálculos se basaron en suposiciones incorrectas sobre las superficies de enfriamiento disponibles, no se conocían los grandes sacos de aire y una masa corporal inflada en exceso. Estas imprecisiones resultaron en la sobreestimación de la producción de calor y la subestimación de la pérdida de calor.[52] Se ha postulado que el gran arco nasal es una adaptación para enfriar el cerebro, como una superficie para el enfriamiento evaporativo de la sangre.[53] Si los Brachiosaurus hubiesen sido endotérmicos (de sangre caliente), debieron haber tardado 10 años para alcanzar su madurez, pero si hubiese sido poiquilotermo, de sangre fría, hubiesen requerido más de 100 años. Como animal de sangre caliente, las demandas energéticas diarias de un braquiosaurio habrían sido enormes, necesitando probablemente comer más que 182 kilogramos de alimento por día. En cambio si el braquiosaurio era completamente de sangre fría o una pasiva gigantotermia, habría necesitado mucho menos alimento para cubrir sus necesidades energéticas diarias. Algunos científicos han propuesto que los dinosaurios grandes como 'Brachiosaurus se mantenían calientes por su enorme tamaño.[54]
Paleoecología
Brachiosaurus es conocido solo en la Formación Morrison del oeste de América del Norte,[5] la cual se interpreta como formada en un entorno semiárido con estaciones secas y húmedas,[55][56] y de llanuras de inundación.[55] la vegetación variaba de bosques en galería, bosque vinculado a la ribera de un río, de coníferas, helechos arborescentes y helechos, a las sabanas de helechos con raros árboles similares a una araucaria.[57] Varios otros géneros de saurópodos estaban presentes en la Formación Morrison, con diferentes proporciones del cuerpo y adaptaciones alimenticias.[8] Entre estos se encontraban Apatosaurus, Brontosaurus, Barosaurus, Camarasaurus, Diplodocus, Haplocanthosaurus y Supersaurus.[8][58] Brachiosaurus era uno de los saurópodos menos abundantes de la Formación de Morrison. En un estudio de más de 200 localidades fósiles, John Foster informó que solo hay 12 ejemplares pertenecientes al género, comparable a Barosaurus (13) y Haplocanthosaurus (12), pero mucho menos que Apatosaurus (112), Camarasaurus (179), y Diplodocus (98).[8] Los fósiles de Brachiosaurus sólo se encuentran en la parte media-baja de la Formación Morrison (zonas estratigráficas 2-4), que datan de hace unos 154-153 millones de años,[59] a diferencia de muchos otros tipos de saurópodos que se han encontrado a lo largo de la formación.[8]
La Formación Morrison registra un ambiente y tiempo dominado por dinosaurios saurópodos gigantescos.[60] Otros dinosaurios conocidos de Morrison incluyen terópodos como Koparion, Stokesosaurus, Ornitholestes, Ceratosaurus, Allosaurus y Torvosaurus, los saurópodos Apatosaurus, Camarasaurus y Diplodocus, y los ornitisquios Camptosaurus, Dryosaurus, Othnielia, Gargoyleosaurus y Stegosaurus.[61] Diplodocus se encuentra comúnmente en los mismos sitios que Apatosaurus, Allosaurus, Camarasaurus y Stegosaurus.[62] Allosaurus representan del 70 al 75% de los especímenes de terópodos y era el depredador dominante de la cadena trófica de Morrison.[63] Muchos de los dinosaurios de la Formación Morrison son los mismos géneros que los que se ven en las rocas portuguesas de la Formación Lourinhã, principalmente Allosaurus, Ceratosaurus, Torvosaurus y Stegosaurus o tienen una contraparte cercana, Brachiosaurus y Lusotitan, Camptosaurus y Draconyx.[64] Otros vertebrados que compartió esta paleoambiente incluyen peces con aletas radiadas, ranas, salamandras, tortugas como Dorsetochelys, esfenodontes, lagartos, crocodilomorfos terrestres y acuáticos como Hoplosuchus y varias especies de pterosaurios como Harpactognathus y Mesadactylus. Conchas de bivalvos y caracoles acuáticos también son comunes. La flora de la época ha sido revelada por fósiles de algas verdes, hongos, musgos, colas de caballo, cícadas, ginkgos y varias familias de coníferas. La vegetación variaba de los bosques de helechos arborescentes y helechos en bosques de galería, a las sabanas del helecho con árboles ocasionales tales como la conífera parecida a la araucaria Brachyphyllum.[65]
En la cultura popular

Brachiosaurus es uno de los dinosaurios más conocidos entre los paleontólogos y el público en general. Un asteroide, en el cinturón de asteroides, 1991 GX7, fue llamado (9954) Brachiosaurus en honor al género.[66][67] Como tal, el género ha aparecido en muchas películas y programas de televisión, especialmente Jurassic Park, Jurassic Park 3, Jurassic World: El reino caído y Walking with Dinosaurs. El modelo digital de Brachiosaurus utilizado en Parque Jurásico se convirtió en el punto de partida para el modelo del ronto en la edición especial de 1997 de la película de ciencia ficción Star Wars: Episode IV - A New Hope.[68][69]
En los museos
Una réplica del esqueleto de Brachiosaurus está montado desde enero del 2000 en la sala B de la Terminal Uno de United Airlines en el Aeropuerto Internacional O'Hare de Chicago (Estados Unidos), una cortesía del Museo Field de Historia Natural de Chicago. Otra idéntica, pero en bronce se encuentra en el exterior del propio museo.[70]
Referencias
- Upchurch, P., Barrett, P.M. & Dodson, P. (2004): "Sauropoda." Pp. 259-322 in Weishampel, D.B., Dodson, P. and Osmolska, H. (eds.): The Dinosauria, Second Edition. University of California Press, Berkeley. ISBN 978-0-520-24209-8
- Foster, J. (2007). "Brachiosaurus altithorax." Jurassic West: The Dinosaurs of the Morrison Formation and Their World. Indiana University Press. pp. 205–208.
- Wedel, M.J. (2003). «Vertebral pneumaticity, air sacs, and the physiology of sauropod dinosaurs». Paleobiology 29: 243-255. doi:10.1666/0094-8373(2003)029<0243:vpasat>2.0.co;2.
- Wedel, M.J. (2003). «The evolution of vertebral pneumaticity in sauropod dinosaurs». Journal of Vertebrate Paleontology 23: 344-357. doi:10.1671/0272-4634(2003)023[0344:teovpi]2.0.co;2.
- Taylor, M.P. (2009). «A re-evaluation of Brachiosaurus altithorax Riggs 1903 (Dinosauria, Sauropoda) and its generic separation from Giraffatitan brancai (Janensh 1914)» (PDF). Journal of Vertebrate Paleontology 29 (3): 787-806. doi:10.1671/039.029.0309.
- Riggs, E.S. (1903). «Brachiosaurus altithorax, the largest known dinosaur». American Journal of Science. 4 15 (88): 299-306.
- Paul, G.S. (1988). «The brachiosaur giants of the Morrison and Tendaguru with a description of a new subgenus, Giraffatitan, and a comparison of the world's largest dinosaurs» (pdf). Hunteria 2 (3).
- Foster, J.R. (2003). Paleoecological analysis of the vertebrate fauna of the Morrison Formation (Upper Jurassic), Rocky Mountain region, U.S.A. New Mexico Museum of Natural History and Science Bulletin, 23. Albuquerque, New Mexico: New Mexico Museum of Natural History and Science.
- Benson, R. B. J.; Campione, N. S. E.; Carrano, M. T.; Mannion, P. D.; Sullivan, C.; Upchurch, P.; Evans, D. C. (2014). «Rates of Dinosaur Body Mass Evolution Indicate 170 Million Years of Sustained Ecological Innovation on the Avian Stem Lineage». PLoS Biology 12 (5): e1001853. doi:10.1371/journal.pbio.1001853.
- Mazzetta, G.V.; Christiansen, P.; Farina, R.A. (2004). «Giants and bizarres: body size of some southern South American Cretaceous dinosaurs». Historical Biology 2004: 1-13. doi:10.1080/08912960410001715132.
- Holtz, T.R. Jr. (2008) Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages Supplementary Information
- Stevens, K. A.; Parrish, M. J. (1999). «Neck posture and feeding habits of two Jurassic sauropod dinosaurs». Science 284: 798-800. PMID 10221910. doi:10.1126/science.284.5415.798.
- Stevens, K. A. and Parrish, M. J. (2005). "Digital reconstructions of sauropod dinosaurs and implications for feeding." In The sauropods: evolution and paleobiology (eds. J. A.Wilson & K. Curry-Rogers), pp. 178–200. Berkeley, CA: University of California Press.
- Stevens, K. A. and Parrish, M. J. (2005). "Neck posture, dentition and feeding strategies in Jurassic sauropod dinosaurs." In Thunder Lizards: The Sauropodomorph dinosaurs" (eds. V. Tidwell & K. Carpenter). Bloomington, IN: Indiana University Press.
- Dzemski, G.; Christian, A. (2007). «Flexibility along the neck of the ostrich (Struthio camelus) and consequences for the reconstruction of dinosaurs with extreme neck length». Journal of Morphology 268: 701-714. doi:10.1002/jmor.10542.
- Glut, D.F. (1997). «Brachiosaurus». Dinosaurs: The Encyclopedia. McFarland & Company. pp. 213–221. ISBN 978-0-89950-917-4.
- Turner, C.E.; Peterson, F. (1999). «Biostratigraphy of dinosaurs in the Upper Jurassic Morrison Formation of the Western Interior, USA». En Gillete, David D. (ed.), ed. Vertebrate Paleontology in Utah. Miscellaneous Publication 99-1. Salt Lake City, Utah: Utah Geological Survey. pp. 77–114. ISBN 978-1-55791-634-1.
- Chenoweth, W.L. (1987). «The Riggs Hill and Dinosaur Hill sites, Mesa County, Colorado». En Averett, W. R. (ed.), ed. Paleontology and Geology of the Dinosaur Triangle. Grand Junction, Colorado: Museum of Western Colorado. pp. 97-100. LCCN 93247073. OCLC 680488874.
- Lohman, S.W. (1965). Geology and artesian water supply of the Grand Junction area, Colorado. Professional Paper 451. Reston, Virginia: U.S. Geological Survey.
- Riggs, E.S. (1901). «The largest known dinosaur». Science 13 (327): 549-550. PMID 17801098. doi:10.1126/science.13.327.549-a.
- Riggs, E.S. (1904). «Structure and relationships of opisthocoelian dinosaurs. Part II. The Brachiosauridae». Geological Series (Field Columbian Museum) 2 (6): 229-247.
- θώραξ, Henry George Liddell, Robert Scott, A Greek-English Lexicon, on Perseus Digital Library
- Marsh, O.C. (1891). «Restoration of Triceratops» (pdf). American Journal of Science 41 (244): 339-342. doi:10.2475/ajs.s3-41.244.339.
- McIntosh, J.S.; Berman, D.S. (1975). «Description of the palate and lower jaw of the sauropod dinosaur Diplodocus (Reptilia: Saurischia) with remarks on the nature of the skull of Apatosaurus». Journal of Paleontology 49 (1): 187-199.
- Carpenter, K. and Tidwell, V. (1998). "Preliminary description of a Brachiosaurus skull from Felch Quarry 1, Garden Park, Colorado." Pp. 69–84 in: Carpenter, K., Chure, D. and Kirkland, J. (eds.), The Upper Jurassic Morrison Formation: An Interdisciplinary Study. Modern Geology, 23:1-4.
- «Expect Awe-Struck Travelers». The Field Museum. 26 de noviembre de 1999. Archivado desde el original el 19 de julio de 2008. Consultado el 27 de agosto de 2009.
- The Field Museum. "Captions from Selected Historical Photographs (caption number GN89396_52c)." The Field Museum Photo Archives. pdf link Archivado el 18 de marzo de 2009 en Wayback Machine.. Accessed 2009-Aug-27.
- Jensen, J.A. (1987). «New brachiosaur material from the Late Jurassic of Utah and Colorado». The Great Basin Naturalist 47 (4): 592-608.
- Curtice, B., Stadtman, K., and Curtice, L. (1996) "A re-assessment of Ultrasauros macintoshi (Jensen, 1985)." Pp. 87-95 in M. Morales (ed.), The Continental Jurassic: Transactions of the Continental Jurassic Symposium, Museum of Northern Arizona Bulletin number 60.
- Curtice, B.; Stadtman, K. (2001). «The demise of Dystylosaurus edwini and a revision of Supersaurus vivianae». En McCord, R.D.; Boaz, D., eds. Western Association of Vertebrate Paleontologists and Southwest Paleontological Symposium - Proceedings 2001. Mesa Southwest Museum Bulletin 8. pp. 33-40.
- Bonnan, M.F.; Wedel, M.J. (2004). «First occurrence of Brachiosaurus (Dinosauria, Sauropoda) from the Upper Jurassic Morrison Formation of Oklahoma». PaleoBios 24 (2): 12-21.
- Jensen, J.A. (1985). «Three new sauropod dinosaurs from the Upper Jurassic of Colorado». The Great Basin Naturalist 45 (4): 697-709.
- Olshevsky, G (1991). «A revision of the parainfraclass Archosauria Cope, 1869, excluding the advanced Crocodylia». Mesozoic Meanderings 2: 1-196.
- Maier, G. (2003). African dinosaurs unearthed. The Tendaguru Expeditions." Bloomington, IN: Indiana University Press.
- Taylor, M. (18 Nov. 2009): CT-scanning the Archbishop. Sauropod Vertebrate Picture of the Week (Blog) post at http://svpow.wordpress.com/2009/11/18/ct-scanning-the-archbishop/.
- Janensch, W (1914). «Übersicht über der Wirbeltierfauna der Tendaguru-Schichten nebst einer kurzen Charakterisierung der neu aufgefuhrten Arten von Sauropoden». Archiv für Biontologie 3: 81-110.
- Janensch, W. (1929). "Material und Formengehalt der Sauropoden in der Ausbeute der Tendaguru-Expedition." Palaeontographica (Suppl. 7) 2:1–34.
- Janensch, W. (1950). "Die Wirbelsäule von Brachiosaurus brancai." Palaeontographica (Suppl. 7) 3:27–93.
- Janensch, W. (1961). "Die Gliedmaßen und Gliedmaßengürtel der Sauropoden der Tendaguru-Schichten." Palaeontographica (Suppl. 7) 3:177–235.
- Lambert, David; the Diagram Group (1990). «Brachiosaurids». The Dinosaur Data Book. New York: Avon Books. p. 142. ISBN 978-0-380-75896-8.
- Chure, D.; Britt, B.; Whitlock, J. A.; Wilson, J. A. (2010). «First complete sauropod dinosaur skull from the Cretaceous of the Americas and the evolution of sauropod dentition». Naturwissenschaften 97 (4): 379-391. PMC 2841758. PMID 20179896. doi:10.1007/s00114-010-0650-6.
- Ksepka, D. T.; Norell, M. A. (2010). «The illusory evidence for Asian Brachiosauridae: new material of Erketu ellisoni and a phylogenetic appraisal of basal Titanosauriformes» (pdf). American Museum Novitates 3700: 1-27. doi:10.1206/3700.2.
- D'Emic, M. D. (2012). «The early evolution of titanosauriform sauropod dinosaurs». Zoological Journal of the Linnean Society 166 (3): 624-671. doi:10.1111/j.1096-3642.2012.00853.x.
- de Lapparent, A.F.; Zbyszewski, G. (1957). «Les dinosauriens du Portugal». Mémoire Service géologique Portugal 2: 1-63.
- Antunes, M.; Mateus, O. (2003). «Dinosaurs of Portugal». Comptes Rendus Palevol 2 (1): 77-95. doi:10.1016/S1631-0683(03)00003-4.
- Janensch, W. (1935-1936). "Die Schädel der Sauropoden Brachiosaurus, Barosaurus und Dicraeosaurus aus den Tendaguru-Schichten Deutsch-Ostafrikas". Palaeontographica (Suppl. 7) 2: 147-298
- Taylor, M.P. (2011). «Correction: A re-evaluation of Brachiosaurus altithorax Riggs 1903 (Dinosauria, Sauropoda) and its generic separation from Giraffatitan brancai (Janensch 1914)".». Journal of Vertebrate Paleontology 31 (3): 727. doi:10.1080/02724634.2011.557115.
- de Lapparent, A.F. (1960): "Les dinosauriens du "continental intercalaire" du Sahara central" ("The dinosaurs of the "continental intercalaire" of the central Sahara.") Mémoires de la Société Géologic de France, Nouvelle Série 88A vol.39(1-6):1-57. [in French; a translated version Archivado el 17 de junio de 2021 en Wayback Machine., by Matthew Carrano (pdf, no figures), is available through the Polyglot Paleontologist]
- Mannion, P. D.; Upchurch, P.; Barnes, R. N.; Mateus, O. (2013). «Osteology of the Late Jurassic Portuguese sauropod dinosaur Lusotitan atalaiensis (Macronaria) and the evolutionary history of basal titanosauriforms» (PDF). Zoological Journal of the Linnean Society 168: 98-206. doi:10.1111/zoj.12029. Archivado desde el original el 11 de mayo de 2013. Consultado el 14 de julio de 2017.
- Barrett, Paul M.; Upchurch, Paul (2005). «Sauropodomorph diversity through time». En Curry Rogers; Kristina A.; Wilson, Jeffrey A., eds. The Sauropods: Evolution and Paleobiology. Berkeley, CA: University of California. pp. 125–156. ISBN 978-0-520-24623-2.
- Mallison, H. (2011). "Rearing Giants – kinetic-dynamic modeling of sauropod bipedal and tripodal poses." In Klein, N., Remes, K., Gee, C. & Sander M. (eds): Biology of the Sauropod Dinosaurs: Understanding the life of giants. Life of the Past (series ed. Farlow, J.). Bloomington, IN: Indiana University Press.
- Sander, P.M.; Christian, A.; Clauss, M.; Fechner, R.; Gee, C.T.; Griebeler, E.-M.; Gunga, H.-C.; Hummel, J.; Mallison, H.; Perry, S.F.; Preuschoft, H.; Rauhut, O.W.M.; Remes, K.; Tütken, T.; Wings, O.; Witzel, U. (2010). «Biology of the sauropod dinosaurs: the evolution of gigantism». Biology Reviews 86 (1): 117-155. PMC 3045712. PMID 21251189. doi:10.1111/j.1469-185X.2010.00137.x.
- Russell, D. A. (1989). An Odyssey in Time: Dinosaurs of North America. Minocqua, Wisconsin: NorthWord Press. p. 78. ISBN 978-1-55971-038-1.
- Bailey, Jack Bowman (1997). "Neural spine elongation in dinosaurs: Sailbacks or buffalo-backs?" Journal of Paleontology 71, 6: 1124-1146
- Russell, D. A. (1989). An Odyssey in Time: Dinosaurs of North America. Minocqua, Wisconsin: NorthWord Press. pp. 64-70. ISBN 978-1-55971-038-1.
- Engelmann, G.F.; Chure, D.J.; Fiorillo, A.R. (2004). «The implications of a dry climate for the paleoecology of the fauna of the Upper Jurassic Morrison Formation». Sedimentary Geology 167 (3-4): 297-308. doi:10.1016/j.sedgeo.2004.01.008.
- Carpenter, K. (2006). «Biggest of the big: a critical re-evaluation of the mega-sauropod Amphicoelias fragillimus». En Foster, J. R.; and Lucas, S. G. (eds.), ed. Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin, 36. Albuquerque, New Mexico: New Mexico Museum of Natural History and Science. pp. 131-138.
- Chure, D.J.; Litwin, R.; Hasiotis, S.T.; Evanoff, E.; Carpenter, K. (2006). «The fauna and flora of the Morrison Formation: 2006». En Foster, J.R.; and Lucas, S.G. (eds.), ed. Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin, 36. Albuquerque, New Mexico: New Mexico Museum of Natural History and Science. pp. 233-248.
- Turner, C.E. and Peterson, F., (1999). "Biostratigraphy of dinosaurs in the Upper Jurassic Morrison Formation of the Western Interior, U.S.A." Pp. 77–114 in Gillette, D.D. (ed.), Vertebrate Paleontology in Utah. Utah Geological Survey Miscellaneous Publication 99-1.
- Foster, J. (2007). "Appendix." Jurassic West: The Dinosaurs of the Morrison Formation and Their World. Indiana University Press. pp. 327–329.
- Chure, Daniel J.; Litwin, Ron; Hasiotis, Stephen T.; Evanoff, Emmett; Carpenter, Kenneth (2006). «The fauna and flora of the Morrison Formation: 2006». En Foster, John R.; Lucas, Spencer G., eds. Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin, 36. Albuquerque, New Mexico: New Mexico Museum of Natural History and Science. pp. 233-248.
- Dodson, Peter; Behrensmeyer, A.K.; Bakker, Robert T.; McIntosh, John S. (1980). «Taphonomy and paleoecology of the dinosaur beds of the Jurassic Morrison Formation». Paleobiology 6 (2): 208-232.
- Foster, John R. (2003). Paleoecological Analysis of the Vertebrate Fauna of the Morrison Formation (Upper Jurassic), Rocky Mountain Region, U.S.A. New Mexico Museum of Natural History and Science Bulletin, 23. Albuquerque, New Mexico: New Mexico Museum of Natural History and Science. p. 29.
- Mateus, Octávio (2006). «Jurassic dinosaurs from the Morrison Formation (USA), the Lourinhã and Alcobaça Formations (Portugal), and the Tendaguru Beds (Tanzania): A comparison». En Foster, John R.; Lucas, Spencer G., eds. Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin, 36. Albuquerque, New Mexico: New Mexico Museum of Natural History and Science. pp. 223-231.
- Carpenter, Kenneth (2006). «Biggest of the big: a critical re-evaluation of the mega-sauropod Amphicoelias fragillimus». En Foster, John R.; Lucas, Spencer G., eds. Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin, 36. Albuquerque, New Mexico: New Mexico Museum of Natural History and Science. pp. 131-138.
- «JPL Small-Body Database Browser: 9954 Brachiosaurus (1991 GX7)». NASA. Consultado el 28 de abril de 2007.
- Williams, Gareth. «Minor Planet Names: Alphabetical List». Smithsonian Astrophysical Observatory. Consultado el 10 de febrero de 2007.
- «Ronto». Databank. Star Wars.com. Archivado desde el original el 3 de octubre de 2008. Consultado el 13 de enero de 2009.
- «The sauropods of Star Wars: Special Edition». Consultado el 19 de mayo de 2017.
- «Expect Awe-Struck Travelers. United Terminal to become new home for Field Museum's gigant Brachiosaurus» (en inglés). The Field Museum. 26 de noviembre de 1999. Archivado desde el original el 19 de julio de 2008. Consultado el 15 de febrero de 2010.
Véase también
Enlaces externos
 Portal:Dinosaurios. Contenido relacionado con Dinosaurios.
Portal:Dinosaurios. Contenido relacionado con Dinosaurios. Wikimedia Commons alberga una categoría multimedia sobre Brachiosaurus altithorax.
Wikimedia Commons alberga una categoría multimedia sobre Brachiosaurus altithorax. Wikispecies tiene un artículo sobre Brachiosaurus altithorax.
Wikispecies tiene un artículo sobre Brachiosaurus altithorax.- Brachiosaurus en Dinosaurier-info (en alemán)
- Brachiosaurus dentro de Brachiosauridae en The Thescelsosaurus (en inglés)
- Significado y pronunciación del nombre (en inglés)
- Why was Brachiosaurus that big? Short summary of a peer review paper
- (en inglés) Información general incluidos los especímenes.
| Control de autoridades |
|
|---|
 Datos: Q14397
Datos: Q14397- Multimedia: Brachiosaurus / Q14397
- Especies: Brachiosaurus