Factor de crecimiento insulínico tipo 1
El factor de crecimiento insulínico tipo 1, también conocido como somatomedina C, o IGF-1 (del inglés: insulin-like growth factor 1) es una proteína que en humanos es codificada por el gen IGF1.[1][2] Se le ha referido al IGF-1 como "factor de sulfatación"[3] y sus efectos fueron denominados "actividad insulínica no suprimible" en los años 1970.
| Factor de crecimiento insulínico tipo 1 (somatomedina C) | ||||
|---|---|---|---|---|
 | ||||
| Estructuras disponibles | ||||
| PDB | ||||
| Identificadores | ||||
| Símbolos | IGF1 (HGNC: 5464) IGF-I; IGF1A; IGFI | |||
| Identificadores externos | ||||
| Locus | Cr. 12 q23.2 | |||
| Patrón de expresión de ARNm | ||||
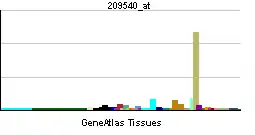 | ||||
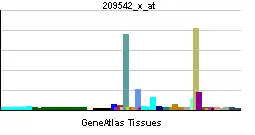 | ||||
| Ortólogos | ||||
| Especies |
| |||
| Entrez |
| |||
| Ensembl |
| |||
| UniProt |
| |||
| RefSeq (ARNm) |
| |||
| RefSeq (proteína) NCBI |
| |||
| Ubicación (UCSC) |
| |||
| PubMed (Búsqueda) |
| |||

El IGF-1 es una hormona similar en estructura molecular a la insulina. Desempeña un importante papel en el crecimiento infantil (los mayores niveles se producen en la pubertad, los menores en la infancia y la vejez), y en el adulto continúa teniendo efectos anabolizantes.
El IGF-1 consiste de 70 aminoácidos en una sola cadena con tres puentes disulfuro intramoleculares; su peso molecular es de 7649 daltons.
Síntesis y circulación
El IGF-1 es una proteína liberada por muchos tejidos y afecta prácticamente a casi todas las células del cuerpo. Los principales órganos sintetizadores del IGF-1 es el hígado, aunque también se produce a nivel local en la placenta, el corazón, el pulmón, el riñón, el páncreas, el bazo, el intestino delgado, los testículos, los ovarios, el intestino grueso, el cerebro, la médula ósea y la hipófisis. La producción es estimulada por la hormona del crecimiento (GH) y puede ser retardada por la desnutrición, la falta de sensibilidad a la hormona del crecimiento, la falta de receptores de hormona del crecimiento, o fallas en la ruta de señalización post-receptores (segundo mensajero) de GH incluyendo la SHP2 y STAT5B. Aproximadamente el 98% del IGF-1 siempre está unido a una de 6 proteínas fijadoras (IGFBP). La IGFBP3, la proteína más abundante, representa el 80% de todas las uniones del IGF. El IGF-1 se une a la IGFBP-3. en una proporción molar 1:1. Esta proteína forma un complejo ternario de 140.000 daltons con el IGF-1 y con una subunidad ácida-lábil.
En experimentos con ratas, la cantidad de IGF-1 mRNA en el hígado fue positivamente asociada con la caseína dietética y negativamente asociada con una dieta libre de proteínas.[4]
Recientemente, fue desarrollado un sistema eficiente de expresión de plantas para producir IGF-I humana biológicamente activa recombinante (rhIGF-I) en granos de arroz transgénicos.[5]
Los seres humanos producen aproximadamente 30 µg (microgramos) de IGF-1 al día hasta cumplir los 30 años y desde este momento la producción decrece con la edad.
Mecanismo de acción
Su acción principal es mediada por la unión a su receptor específico, el receptor de factor de crecimiento insulínico tipo 1, abreviado como IGF1R, presente en muchos tipos de tejidos. En la unión al IGF1R, un receptor tirosina quinasa, inicia la señalización intracelular; el IGF-1 es uno de los activadores naturales más potentes de la transducción de señal PKB, un estimulador del crecimiento y proliferación celular, y un potente inhibidor de la muerte celular programada.
El IGF-1 es un mediador principal de los efectos de la hormona del crecimiento (GH). La hormona del crecimiento es producida en la adenohipófisis y liberada al torrente sanguíneo, y luego estimula el hígado a producir IGF-1. El IGF-1 luego estimula el crecimiento del cuerpo de forma sistémica, y tiene efectos promotores del crecimiento en casi todas las células del cuerpo, especialmente el músculo esquelético, cartílago, hueso, hígado, nervios, piel, células hematopoyéticas, y pulmón. Además de los efectos similares a la insulina, el IGF-1 también puede regular el desarrollo y crecimiento celular, especialmente en las células nerviosas, como también la síntesis de ADN celular.
Por lo tanto, la deficiencia de ya sea de hormona del crecimiento o IGF-1 resultaría en una estatura disminuida. Los niños deficientes de GH son dados GH recombinante para incrementar su tamaño. Los humanos deficientes de IGF-1, quienes están clasificados de padecer del síndrome de Laron, o enanismo de Laron, son tratados con IGF-1 recombinante. En el ganado bovino, el IGF-1 circulante está relacionado con el desempeño reproductivo.[6]
Receptores
El IGF-1 se une a al menos a dos receptores de la membrana celular: el receptor de IGF-1 (IGF1R), y el receptor de insulina. El IGF-1 tiene una alta afinidad por el receptor de IGF-1, y una baja afinidad por el receptor de insulina. Estos receptores son tirosina quinasa (significando que señalizan causando la adición de una molécula de fosfato en ciertas tirosinas). El IGF-1 activa el receptor insulínico aproximadamente a una potencia 0.1x veces que la insulina.
El IGF-1 es producido durante toda la vida. Los mayores niveles se producen durante el crecimiento pubertad, los menores en la infancia y la vejez.
Otras IGF-BPs (proteínas fijadoras/transportadoras) son inhibitorias. Por ejemplo, ambas IGFBP-2 y IGFBP-5 se unen al IGF-1 con una afinidad mayor que la afinidad con que el IGF-1 se une con su receptor. Por lo tanto, el incremento de los niveles séricos de estas dos IGF-BPs resultaría en una disminución en la actividad del IGF-1.
Contribución al envejecimiento
Es ampliamente aceptado que la señalización a través de la ruta de receptores de insulina/IGF-1 es un contribuyente significativo en el proceso de envejecimiento biológico en muchos organismos. Esta línea de investigación adquirió importancia con el trabajo de Cynthia Kenyon, quien mostró que las mutaciones en el gen Daf-2 podría duplicar la vida de un nematodo C. elegans.[7][8] El gen daf-2 codifica los receptores de insulina/IGF-1 de dicho nematodo.
La señalización insulina/IGF-1 es conservada desde las lombrices a los humanos. Según estudios posteriores al trabajo de Kenyon, las mutaciones que reducen la señalización insulina/IGF-1 han demostrado desacelerar el proceso degenerativo del envejecimiento y extender la vida de una amplia gama de organismos, incluyendo Drosophila melanogaster, ratones,[9] y posiblemente humanos.[10][11][12][13]
Se cree también, que la reducción de la señalización IGF-1 contribuye a los efectos "anti-envejecimiento" en la restricción calórica.[14]
Factores que influyen los niveles en circulación
Los factores que son conocidos por causar variaciones en los niveles de hormona del crecimiento (GH) e IGF-1 en circulación incluyen: genética, la hora del día, la edad, sexo, ejercicio, niveles de estrés, niveles de nutrición e índice de masa corporal (IMC), estado de salud, raza, estado de estrógenos, e ingesta de xenobióticos.[15] La inclusión de ingesta de xenobióticos como factor que influye el estado de GH-IGF circulante resalta el hecho que el eje GH-IGF es un potencial objetivo de ciertos químicos perturbadores endocrinos - véase interruptor endocrino.
Enfermedades de deficiencia y resistencia
Se han descrito enfermedades raras por fallos en la producción o respuesta al IGF-I, que resultan en una alteración específica del crecimiento. Uno de estos trastornos, el síndrome de Laron, no responde en absoluto al tratamiento con hormona del crecimiento debido a una falta de receptores de GH. La FDA ha agrupado estas enfermedades en un trastorno llamado deficiencia primaria de IGF (IGFD) severa. Los pacientes afectados presentan niveles normales a elevados de GH, una altura por debajo de -3 desviaciones estándar (DS), y niveles de IGF por debajo de -3 DS. La deficiencia primaria de IGF severa incluye pacientes con mutaciones en el receptor de GH, mutaciones post-receptor o mutaciones de IGF. Como resultado para la apoptosis, los pacientes no responden al tratamiento con GH.
La ruta de señalización del IGF parece desempeñar un papel importante en el cáncer. Varios estudios han demostrado que niveles altos de IGF aumentan el riesgo de cáncer.[16] Estudios hechos en células de cáncer de pulmón muestran que los medicamentos que inhiben esta señalización podrían ser en el futuro una potente arma terapéutica contra el cáncer.[17]
Utilidades clínicas
Como prueba diagnóstica
| Rangos de referencia para IGF-1[18] (en ng/mL) | ||||
|---|---|---|---|---|
| Edad | Mujeres | Hombres | ||
| 2,5.ª percentil | 97,5.ª percentil | 2,5.ª percentil | 97,5.ª percentil | |
| 20 | 111 | 423 | 156 | 385 |
| 25 | 102 | 360 | 119 | 343 |
| 30 | 94 | 309 | 97 | 306 |
| 35 | 86 | 271 | 84 | 275 |
| 40 | 79 | 246 | 76 | 251 |
| 45 | 73 | 232 | 71 | 233 |
| 50 | 68 | 228 | 66 | 221 |
| 55 | 64 | 231 | 61 | 214 |
| 60 | 61 | 237 | 55 | 211 |
| 65 | 59 | 241 | 49 | 209 |
| 70 | 57 | 237 | 46 | 207 |
| 75 | 55 | 219 | 48 | 202 |
Los niveles de IGF-I se pueden medir en sangre, con un rango de normalidad de 10 a 1000 ng/ml. Como los niveles no fluctúan mucho a lo largo del día para cada persona, se utilizan en pruebas de screening para detectar la deficiencia y el exceso de GH.
La interpretación de los niveles de IGF-I es complicada, dada la amplitud del rango de normalidad, y sus variaciones por edad, sexo y estado puberal. Clínicamente, alteraciones significativas pueden estar enmascaradas por dicha amplitud de rango. Suele resultar más útil la determinación secuencial de los niveles, especialmente en determinadas patologías hipofisarias, desnutrición y problemas del crecimiento.
Como agente terapéutico
La mecasermina (nombre comercial Increlex) es un análogo sintético del IGF-1 que está aprobado para el tratamiento de la falta de crecimiento.[19] El IGF-1 ha sido fabricado recombinantemente a gran escala utilizando la levadura y E.coli.
Se han realizado ensayos clínicos para evaluar la posible eficacia del IGF-I recombinante en una multitud de patologías: problemas del crecimiento, diabetes mellitus tipos 1 y 2, esclerosis lateral amiotrófica (ELA, también conocido como "enfermedad de Lou Gehrig"), quemados severos, y distrofia muscular miotónica. Los ensayos muestran gran eficacia en la diabetes mellitus, en cuanto a la reducción de los niveles de hemoglobina A1C, así como el consumo diario de insulina. Sin embargo, la empresa patrocinadora del ensayo (Genentech), discontinuó el ensayo debido a la exacerbación de la retinopatía diabética en ciertos pacientes. En cuanto a su uso para la ELA, los laboratorios Cephalon y Chiron llevaron a cabo dos ensayos: uno demostró su eficacia terapéutica y el resultado del segundo era ambiguo, por lo que su uso no fue aprobado por la FDA.
Sin embargo, debido a los esfuerzos del laboratorio Tercica, en agosto de 2005 la FDA aprobó el uso de un tipo de IGF-I recombinante, Increlex, como terapia sustituiva para pacientes con un déficit severo de IGF-I, tras un ensayo con 71 pacientes. En diciembre del mismo año, la FDA aprobó Inplex (del laboratorio Insmed), un complejo IGF-I/IGF BP-3. Este fármaco se inyecta en una sola dosis diaria, frente a las dos necesarias para Increlex, por lo que los efectos secundarios son menores para una misma eficacia.
Insmed fue acusado de infringir la licencia de patente de Tercica, por lo que fue llevado a los tribunales con la intención de que prohibieran la venta de Inplex.[20] En consecuencia, Increlex es actualmente el único fármaco derivado de IGF-I en el mercado estadounidense.[21]
En un ensayo clínico de un compuesto en investigación llamado MK-677, que eleva el IGF-1 de los pacientes, no resultó en una mejora de los síntomas del Alzheimer.[22] Otro ensayo demostró que el IGF-1 de Cephalon no retrasa la progresión de la debilidad en pacientes con ELA. Estudios previos de menor duración tuvieron resultados conflictivos.[23]
La IGFBP-3 es un portador del IGF-1, significando que el IGF-1 se une a la IGFBP-3, creando un complejo cuyo peso molecular combinado y afinidad de unión le permite al factor de crecimiento tener una vida media incrementada en suero. Sin su unión con la IGFBP-3, el IGF-1 es rápidamente eliminada a través del riñón, debido a su bajo peso molecular. Al estar unido a la IGFBP-3, el IGF-1 evade la eliminación renal. También, debido que la IGFBP-3 tiene una afinidad menor con el IGF-1 que el IGF-1 tiene con su receptor, el receptor de factor de crecimiento insulínico tipo 1 (IGFR), su unión con la IGFBP-3 no interfiere con su función. Por estas razones, una combinación IGF-1/IGFBP-3 fue aprobada para el tratamiento humano.
Se ha demostrado también que el IGF-1 es efectivo en accidentes cerebrovasculares, en modelos animales, cuando este es combinado con eritropoyetina. Se obtuvieron mejoras conductuales y celulares.[24]
Interacciones
Se ha demostrado que el factor de crecimiento insulínico tipo 1 se une e interactúa con las seis Proteínas Fijadoras de IGF-1 (IGFBP 1-6).
Se proporcionan referencias específicas para las interacciones con IGFBP3,[25][26][27][28][29][30] IGFBP4,[31][32] e IGFBP7.[33][34]
Referencias
- Höppener JW, de Pagter-Holthuizen P, Geurts van Kessel AH, Jansen M, Kittur SD, Antonarakis SE, Lips CJ, Sussenbach JS (1985). «The human gene encoding insulin-like growth factor I is located on chromosome 12». Hum. Genet. 69 (2): 157-60. PMID 2982726. doi:10.1007/BF00293288.
- Jansen M, van Schaik FM, Ricker AT, Bullock B, Woods DE, Gabbay KH, Nussbaum AL, Sussenbach JS, Van den Brande JL (1983). «Sequence of cDNA encoding human insulin-like growth factor I precursor». Nature 306 (5943): 609-11. PMID 6358902. doi:10.1038/306609a0.
- Salmon W, Daughaday W (1957). «A hormonally controlled serum factor which stimulates sulfate incorporation by cartilage in vitro». J Lab Clin Med 49 (6): 825-36. PMID 13429201.
- Miura, Y.; Kato, H.; Noguchi, T. (2007). «Effect of dietary proteins on insulin-like growth factor-1 (IGF-1) messenger ribonucleic acid content in rat liver». British Journal of Nutrition (en inglés) 67 (2): 257. PMID 1596498. doi:10.1079/BJN19920029.
- Cheung SC, Liu LZ, Lan LL, Liu QQ, Sun SS, Chan JC, Tong PC.,"Glucose lowering effect of transgenic human insulin-like growth factor-I from rice: in vitro and in vivo studies." BMC Biotechnol. 2011;11:37
- Yilmaz A, Davis ME, RCM Simmen RCM (1999). «Reproductive performance of bulls divergently selected on the basis of blood serum insulin-like growth factor I concentration». J Anim Sci 77 (4): 835-9.
- See publications documenting series of experiments at Cynthia Kenyon lab, in particular, Dorman JB, Albinder B, Shroyer T, Kenyon C (diciembre de 1995). «The age-1 and daf-2 genes function in a common pathway to control the lifespan of Caenorhabditis elegans». Genetics 141 (4): 1399-406. PMC 1206875. PMID 8601482.
- Apfeld J, Kenyon C (octubre de 1998). «Cell nonautonomy of C. elegans daf-2 function in the regulation of diapause and life span». Cell 95 (2): 199-210. PMID 9790527. doi:10.1016/S0092-8674(00)81751-1.
- Bartke, A (12 de enero de 2011). «Single-gene mutations and healthy ageing in mammals». Philos Trans R Soc Lond B Biol Sc 366 (1561): 28-34. PMID 21115527 doi 10.1098/rstb.2010.0281 10.1098/rstb.2010.0281. Consultado el 28 de septiembre de 2011.
- Guevara-Aguirre, J; .Guevara-Aguirre J, Balasubramanian P, Guevara-Aguirre M, Wei M, Madia F, Cheng CW, Hwang D, Martin-Montalvo A, Saavedra J, Ingles S, de Cabo R, Cohen P, Longo VD (16 de febrero de 2011). «Growth hormone receptor deficiency is associated with a major reduction in pro-aging signaling, cancer, and diabetes in humans.». Sci Transl Med. 3 (70): 70ra13. PMID 21325617.
- Pawlikowska, L; Hu D, Huntsman S, Sung A, Chu C, Chen J, Joyner AH, Schork NJ, Hsueh WC, Reiner AP, Psaty BM, Atzmon G, Barzilai N, Cummings SR, Browner WS, Kwok PY, Ziv E; Study of Osteoporotic Fractures (2019-08). «Association of common genetic variation in the insulin/IGF1 signaling pathway with human longevity.». Sci Transl Med. 8 (4): 460-72. PMID 19489743.
- Suh, Y; Suh Y; Atzmon G; Cho MO; Hwang D; Liu B; Leahy DJ; Barzilai N et al. (2008-03). «Functionally significant insulin-like growth factor I receptor mutations in centenarians». Aging Cell 105 (9): 3438-42. PMID 18316725.
- Van Heemst, D; Beekman M; Mooijaart SP; Heijmans BT; Brandt BW; Zwaan BJ; Slagboom PE; Westendorp RG (2005 Apr). «Reduced insulin/IGF-1 signalling and human longevity». Aging Cell 4 (2): 79-85. PMID 15771611.
- Barzilai, N; A Bartke (2009 Feb 19). «Biological approaches to mechanistically understand the healthy life span extension achieved by calorie restriction and modulation of hormones». J Gerontol A Biol Sci Med Sci 64 (2): 187-91. PMID 19228789 doi 10.1093/gerona/gln061 10.1093/gerona/gln061. Consultado el 28 de septiembre de 2011.
- Scarth J (2006). «Modulation of the growth hormone-insulin-like growth factor (GH-IGF) axis by pharmaceutical, nutraceutical and environmental xenobiotics: an emerging role for xenobiotic-metabolizing enzymes and the transcription factors regulating their expression. A review». Xenobiotica 36 (2-3): 119-218. PMID 16702112. doi:10.1080/00498250600621627.
- Smith GD, Gunnell D, Holly J (2000). «Cancer and insulin-like growth factor-I.». British Medical Journal 321: 847-8. doi:10.1136/bmj.321.7265.847.
- Velcheti V, Govindan R (2006). «Insulin-like growth factor and lung cancer». Journal of thoracic oncology : official publication of the International Association for the Study of Lung Cancer 1 (7): 607-10. PMID 17409926.
- Ranges estimated from quantile regression as shown in table 4 in: Friedrich, N.; Alte, D.; Völzke, H.; Spilcke-Liss, E.; Lüdemann, J.; Lerch, M. M.; Kohlmann, T.; Nauck, M. et al (2008). «Reference ranges of serum IGF-1 and IGFBP-3 levels in a general adult population: Results of the Study of Health in Pomerania (SHIP)». Growth Hormone & IGF Research (en inglés) 18 (3): 228-237. PMID 17997337. doi:10.1016/j.ghir.2007.09.005.
- Rosenbloom AL (agosto de 2007). «The role of recombinant insulin-like growth factor I in the treatment of the short child». Curr. Opin. Pediatr. 19 (4): 458-64. PMID 17630612. doi:10.1097/MOP.0b013e3282094126.
- Pollack A (17 de febrero de 2007). «Growth Drug Is Caught Up in Patent Fight». The New York Times. Consultado el 28 de marzo de 2010.
- Pollack A (7 de marzo de 2007). «To Settle Suit, Maker Agrees to Withdraw Growth Drug». The New York Times. Consultado el 28 de marzo de 2010.
- Sevigny JJ, Ryan JM, van Dyck CH, Peng Y, Lines CR, Nessly ML (noviembre de 2008). «Growth hormone secretagogue MK-677: no clinical effect on AD progression in a randomized trial». Neurology 71 (21): 1702-8. PMID 19015485. doi:10.1212/01.wnl.0000335163.88054.e7.
- Sorenson EJ, Windbank AJ, Mandrekar JN, Bamlet WR, Appel SH, Armon C, Barkhaus PE, Bosch P, Boylan K, David WS, Feldman E, Glass J, Gutmann L, Katz J, King W, Luciano CA, McCluskey LF, Nash S, Newman DS, Pascuzzi RM, Pioro E, Sams LJ, Scelsa S, Simpson EP, Subramony SH, Tiryaki E, Thornton CA (noviembre de 2008). «Subcutaneous IGF-1 is not beneficial in 2-year ALS trial». Neurology 71 (22): 1770-5. PMC 2617770. PMID 19029516. doi:10.1212/01.wnl.0000335970.78664.36. Resumen divulgativo – newswise.com.
- Fletcher L, Kohli S, Sprague SM, Scranton RA, Lipton SA, Parra A, Jimenez DF, Digicaylioglu M (julio de 2009). «Intranasal delivery of erythropoietin plus insulin-like growth factor-I for acute neuroprotection in stroke. Laboratory investigation». J. Neurosurg. 111 (1): 164-70. PMID 19284235. doi:10.3171/2009.2.JNS081199.
- Horton JK, Thimmaiah KN, Houghton JA, Horowitz ME, Houghton PJ (junio de 1989). «Modulation by verapamil of vincristine pharmacokinetics and toxicity in mice bearing human tumor xenografts». Biochem. Pharmacol. 38 (11): 1727-36. PMID 2735930. doi:10.1016/0006-2952(89)90405-X.
- Ueki I, Ooi GT, Tremblay ML, Hurst KR, Bach LA, Boisclair YR (junio de 2000). «Inactivation of the acid labile subunit gene in mice results in mild retardation of postnatal growth despite profound disruptions in the circulating insulin-like growth factor system». Proc. Natl. Acad. Sci. U.S.A. 97 (12): 6868-73. PMC 18767. PMID 10823924. doi:10.1073/pnas.120172697.
- Buckway CK, Wilson EM, Ahlsén M, Bang P, Oh Y, Rosenfeld RG (octubre de 2001). «Mutation of three critical amino acids of the N-terminal domain of IGF-binding protein-3 essential for high affinity IGF binding». J. Clin. Endocrinol. Metab. 86 (10): 4943-50. PMID 11600567. doi:10.1210/jc.86.10.4943.
- Cohen P, Graves HC, Peehl DM, Kamarei M, Giudice LC, Rosenfeld RG (octubre de 1992). «Prostate-specific antigen (PSA) is an insulin-like growth factor binding protein-3 protease found in seminal plasma». J. Clin. Endocrinol. Metab. 75 (4): 1046-53. PMID 1383255. doi:10.1210/jc.75.4.1046.
- Twigg SM, Baxter RC (marzo de 1998). «Insulin-like growth factor (IGF)-binding protein 5 forms an alternative ternary complex with IGFs and the acid-labile subunit». J. Biol. Chem. 273 (11): 6074-9. PMID 9497324. doi:10.1074/jbc.273.11.6074.
- Firth SM, Ganeshprasad U, Baxter RC (enero de 1998). «Structural determinants of ligand and cell surface binding of insulin-like growth factor-binding protein-3». J. Biol. Chem. 273 (5): 2631-8. PMID 9446566. doi:10.1074/jbc.273.5.2631.
- Bach LA, Hsieh S, Sakano K, Fujiwara H, Perdue JF, Rechler MM (mayo de 1993). «Binding of mutants of human insulin-like growth factor II to insulin-like growth factor binding proteins 1-6». J. Biol. Chem. 268 (13): 9246-54. PMID 7683646.
- Qin X, Strong DD, Baylink DJ, Mohan S (septiembre de 1998). «Structure-function analysis of the human insulin-like growth factor binding protein-4». J. Biol. Chem. 273 (36): 23509-16. PMID 9722589. doi:10.1074/jbc.273.36.23509.
- Ahmed S, Yamamoto K, Sato Y, Ogawa T, Herrmann A, Higashi S, Miyazaki K (octubre de 2003). «Proteolytic processing of IGFBP-related protein-1 (TAF/angiomodulin/mac25) modulates its biological activity». Biochem. Biophys. Res. Commun. 310 (2): 612-8. PMID 14521955. doi:10.1016/j.bbrc.2003.09.058.
- Oh Y, Nagalla SR, Yamanaka Y, Kim HS, Wilson E, Rosenfeld RG (noviembre de 1996). «Synthesis and characterization of insulin-like growth factor-binding protein (IGFBP)-7. Recombinant human mac25 protein specifically binds IGF-I and -II». J. Biol. Chem. 271 (48): 30322-5. PMID 8939990. doi:10.1074/jbc.271.48.30322.
Lectura adicional (en inglés)
- Butler AA, Yakar S, LeRoith D (2002). «Insulin-like growth factor-I: compartmentalization within the somatotropic axis?». News Physiol. Sci. 17: 82-5. PMID 11909998.
- Maccario M, Tassone F, Grottoli S, et al. (2002). «Neuroendocrine and metabolic determinants of the adaptation of GH/IGF-I axis to obesity». Ann. Endocrinol. (Paris) 63 (2 Pt 1): 140-4. PMID 11994678.
- Camacho-Hübner C, Woods KA, Clark AJ, Savage MO (2003). «Insulin-like growth factor (IGF)-I gene deletion». Reviews in endocrine & metabolic disorders 3 (4): 357-61. PMID 12424437. doi:10.1023/A:1020957809082.
- Trojan LA, Kopinski P, Wei MX, et al. (2004). «IGF-I: from diagnostic to triple-helix gene therapy of solid tumors». Acta Biochim. Pol. 49 (4): 979-90. PMID 12545204.
- Winn N, Paul A, Musaró A, Rosenthal N (2003). «Insulin-like growth factor isoforms in skeletal muscle aging, regeneration, and disease». Cold Spring Harb. Symp. Quant. Biol. 67: 507-18. PMID 12858577. doi:10.1101/sqb.2002.67.507.
- Delafontaine P, Song YH, Li Y (2005). «Expression, regulation, and function of IGF-1, IGF-1R, and IGF-1 binding proteins in blood vessels». Arterioscler. Thromb. Vasc. Biol. 24 (3): 435-44. PMID 14604834. doi:10.1161/01.ATV.0000105902.89459.09.
- Trejo JL, Carro E, Garcia-Galloway E, Torres-Aleman I (2004). «Role of insulin-like growth factor I signaling in neurodegenerative diseases». J. Mol. Med. 82 (3): 156-62. PMID 14647921. doi:10.1007/s00109-003-0499-7.
- Rabinovsky ED (2004). «The multifunctional role of IGF-1 in peripheral nerve regeneration». Neurol. Res. 26 (2): 204-10. PMID 15072640. doi:10.1179/016164104225013851.
- Rincon M, Muzumdar R, Atzmon G, Barzilai N (2005). «The paradox of the insulin/IGF-1 signaling pathway in longevity». Mech. Ageing Dev. 125 (6): 397-403. PMID 15272501. doi:10.1016/j.mad.2004.03.006.
- Conti E, Carrozza C, Capoluongo E, et al. (2005). «Insulin-like growth factor-1 as a vascular protective factor». Circulation 110 (15): 2260-5. PMID 15477425. doi:10.1161/01.CIR.0000144309.87183.FB.
- Wood AW, Duan C, Bern HA (2005). «Insulin-like growth factor signaling in fish». Int. Rev. Cytol. 243: 215-85. PMID 15797461. doi:10.1016/S0074-7696(05)43004-1.
- Sandhu MS (2005). «Insulin-like growth factor-I and risk of type 2 diabetes and coronary heart disease: molecular epidemiology». Endocrine development. Endocrine Development 9: 44-54. ISBN 3-8055-7926-8. PMID 15879687. doi:10.1159/000085755.
- Ye P, D'Ercole AJ (2006). «Insulin-like growth factor actions during development of neural stem cells and progenitors in the central nervous system». J. Neurosci. Res. 83 (1): 1-6. PMID 16294334. doi:10.1002/jnr.20688.
- Gómez JM (2006). «The role of insulin-like growth factor I components in the regulation of vitamin D». Current pharmaceutical biotechnology 7 (2): 125-32. PMID 16724947. doi:10.2174/138920106776597621.
- Federico G, Street ME, Maghnie M, et al. (2006). «Assessment of serum IGF-I concentrations in the diagnosis of isolated childhood-onset GH deficiency: a proposal of the Italian Society for Pediatric Endocrinology and Diabetes (SIEDP/ISPED)». J. Endocrinol. Invest. 29 (8): 732-7. PMID 17033263.
- Zakula Z, Koricanac G, Putnikovic B, et al. (2007). «Regulation of the inducible nitric oxide synthase and sodium pump in type 1 diabetes». Med. Hypotheses 69 (2): 302-6. PMID 17289286. doi:10.1016/j.mehy.2006.11.045.
- Trojan J, Cloix JF, Ardourel MY, et al. (2007). «Insulin-like growth factor type I biology and targeting in malignant gliomas». Neuroscience 145 (3): 795-811. PMID 17320297. doi:10.1016/j.neuroscience.2007.01.021.
Enlaces externos
- Silvia Fernández et alii, «Función neuroprotectora de los factores insulínicos», Mente y Cerebro, 38, 2009.
- MeSH: Insulin-Like+Growth+Factor+I (en inglés)
| Control de autoridades |
|
|---|
 Datos: Q12791246
Datos: Q12791246