Hirundo rustica
La golondrina común (Hirundo rustica)[5] es una especie de ave paseriforme de la familia Hirundinidae de hábitos migratorios. Se trata de la golondrina con la mayor área de distribución del mundo. Habita en Europa, Asia, África, América y parte de Australasia.
| Golondrina común | ||
|---|---|---|
 Subespecie europea (H. r. rustica) en Dinamarca. | ||
| Estado de conservación | ||
 Preocupación menor (UICN 3.1)[1][2][3] | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Aves | |
| Orden: | Passeriformes | |
| Familia: | Hirundinidae | |
| Género: | Hirundo | |
| Especie: |
H. rustica Linnaeus, 1758[2][3][4] | |
| Distribución | ||
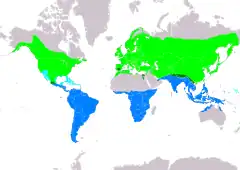 Distribución de Hirundo rustica Área de cría. Área de ocupación sedentaria. Área de paso. Área de invernada. | ||
Existen seis subespecies de golondrina común, que se reproducen a lo largo del hemisferio norte. Cuatro de ellas son fuertemente migratorias y sus cuarteles de invernada cubren gran parte del hemisferio sur, llegando a Argentina central, Sudáfrica y el norte de Australia.[6] Su vasta área de distribución y enorme población implican que la especie se encuentra fuera de peligro de extinción,[2][3] aunque pueda haber decrecimientos poblacionales locales debido a amenazas específicas.[6][7]
La golondrina común es un ave de campo abierto que normalmente utiliza estructuras construidas por el ser humano para reproducirse y que, en consecuencia, se ha diseminado con la expansión humana. Construye nidos en forma de taza,[8] con bolitas de barro en graneros y construcciones similares, y se alimenta de insectos que atrapa en vuelo.[9]
Su alimentación insectívora contribuye a la tolerancia de los humanos hacia esta especie; en el pasado esta aceptación se vio reforzada por supersticiones respecto del ave y su nido. Existen frecuentes referencias culturales a la golondrina común en escritos literarios tanto debido a su proximidad con el ser humano como por su migración anual. Suelen volar en conjuntos.[10]
Descripción
Se trata de un ave pequeña.[11] El macho adulto de la subespecie nominal (H. r. rustica) mide entre 14,6 y 19,9 cm de longitud, incluyendo los 2 a 7 cm de las alargadas plumas externas de la cola. Tiene una envergadura de entre 32 y 34,5 cm y pesa entre 16 y 22 g. Su parte superior es azul metálico y su frente, barbilla y garganta, rojizos. Una gruesa franja azul oscuro separa la garganta del pecho y abdomen blancuzcos. Las plumas externas de la cola son alargadas, lo que le brinda el distintivo aspecto profundamente ahorquillado de las golondrinas. Presenta una línea de manchas blancas a lo largo del extremo externo de la parte superior de la cola.[9] La apariencia de la hembra es similar a la del macho, pero las plumas de la cola son más cortas, el azul de la parte superior y de la banda del pecho es menos lustroso y el pecho y abdomen son más pálidos. El juvenil es más marrón y exhibe una cara rojiza más pálida y partes inferiores más blancas; carece de las largas plumas de la cola de los adultos.[6]
La distintiva combinación del rostro rojizo y la banda pectoral azul facilitan la distinción entre la golondrina común adulta y las especies africanas de Hirundo y de la golondrina australiana (Hirundo neoxena), cuya área de distribución se superpone en Australasia.[6] En África, la cola más corta de los juveniles puede producir una confusión con el juvenil de la golondrina de pecho rojo (Hirundo lucida), pero esta última presenta una banda pectoral más angosta y más blanca en la cola.[12]
El canto de la golondrina común es un gorjeo que a menudo termina en un su-seer en el que la segunda nota es más alta que la primera si bien su altura tonal es decreciente. Las llamadas incluyen un witt o un witt-witt o un fuerte splee-plink cuando están exaltadas o cuando intentan ahuyentar depredadores de las cercanías del nido.[9] Las llamadas de alarma incluyen un agudo siflitt para depredadores como los gatos y un flitt-flitt para aves de presa.[13] Esta especie es bastante tranquila en sus cuarteles de invernada.[14]
La muda de las plumas de vuelo tiene lugar en los cuarteles de invernada, lo que dificulta la identificación subespecífica de los individuos en invierno. La muda posnupcial es muy gradual y en Europa comienza en agosto antes de la migración con la muda del plumaje corporal y a veces con la de las plumas cobertoras medias de las alas. En la India, la muda se produciría lenta e irregularmente, extendiéndose sobre la mayor parte del invierno y no habiendo concluido aún en abril en algunos especímenes.[8]
A diferencia de los vencejos (Apus apus), las golondrinas se pueden posar horizontalmente, ya que la longitud de sus patas les proporcionan un apoyo adecuado para iniciar el vuelo de nuevo desde el suelo.
Taxonomía

La golondrina común fue descrita como especie por Carlos Linneo en 1758 en la décima edición de su obra Systema naturae bajo el nombre científico de Hirundo rustica.[15] Hirundo significa “golondrina” en latín; rusticus quiere decir “campestre”. Es la única de las especies del género Hirundo que se ha extendido a las Américas, siendo la mayoría de ellas nativa de África.
Existen pocos problemas taxonómicos dentro del género, si bien la golondrina de pecho rojo (Hirundo lucida) ―una especie sedentaria del oeste de África, la cuenca del Congo y Etiopía― era anteriormente considerada una subespecie de golondrina común. La golondrina de pecho rojo es ligeramente más pequeña que su pariente migratorio, presenta una franja pectoral azul más angosta y las plumas de la cola de los adultos son más cortas. En vuelo, exhibe una coloración más pálida en sus partes inferiores.[12]
Subespecies

Existen seis subespecies de golondrina común ampliamente reconocidas. Se han propuesto subespecies adicionales o alternativas para Asia Oriental, incluyendo saturata (Robert Ridgway, 1883),[4][16] kamtschatica (Benedykt Dybowski, 1883),[4] mandschurica (Wilhelm Meise, 1934)[4][16] y ambigua (Erwin Stresemann, 1940).[4][8][17] Dada la incierta validez de estas formas,[4][8][16] este artículo sigue la clasificación de Turner y Rose (1989):[6]
- H. r. rustica, la subespecie nominal, se reproduce desde Europa, Asia Menor e Irak hacia el este hasta la cuenca del río Yeniséi, el oeste del macizo de Altái, Sinkiang (China) y Sikkim (India),[4] tan al norte como el círculo polar ártico y hacia el sur hasta el norte de África. Migra a África, Arabia y partes del subcontinente indio,[6] como Sind, Punyab y Sri Lanka.[8][18] Las golondrinas comunes que pasan el invierno en África austral llegan de gran parte de Eurasia, desde al menos el meridiano 91 este; alrededor del 80 % de las golondrinas que migran a África austral proceden de Rusia.[19] Esta subespecie se caracteriza por presentar una franja pectoral completa.[4][8] Existen especímenes ocasionales a lo largo del área de distribución reproductiva de H. r. rustica cuya banda pectoral está más o menos difuminada con rojizo, pero estos ejemplares son escasos y su banda pectoral no está difuminada en el mismo grado que en gutturalis, lo que facilitaría la identificación de ambas subespecies en los cuarteles de invernada.[8] Sus partes inferiores se vuelven más blancas con el desgaste de las plumas, pero algunos individuos pueden tenerlas blancas incluso con su plumaje recientemente mudado. Otras coloraciones posibles son rosáceo, rosáceo pálido, crema, blancuzco y blanco sucio.[8] El brillo metálico de sus partes superiores es azul violáceo.[4][8] Es más grande que gutturalis.[8]
- H. r. transitiva fue descrita por Ernst Hartert en 1910. Se reproduce en el Medio Oriente, desde el sur de Turquía hasta Israel, y es parcialmente sedentaria, aunque algunos ejemplares pasan el invierno en África Oriental. Presenta partes inferiores rojo-anaranjadas y un quiebro en la franja pectoral.[6]
- H. r. savignii, la subespecie sedentaria de Egipto, fue descrita por James Francis Stephens en 1817 y bautizada en honor al zoólogo francés Marie Jules César Lelorgne de Savigny.[20] Se asemeja a transitiva, que también exhibe parte inferiores rojo-anaranjadas, pero savignii presenta una franja pectoral completa y un matiz rojizo más profundo en las partes inferiores.[13]
- H. r. gutturalis, descrita por Giovanni Antonio Scopoli en 1786,[4][16] presenta partes inferiores típicamente ocre oxidado, pero el color varía a lo largo de estas (no es uniforme como en tytleri).[4] Exhibe un quiebro en la franja pectoral y el brillo metálico de sus partes superiores es azul verdoso.[4][8] Se reproduce en Asia oriental, al este de tytleri y de la subespecie nominal; entre Yakutsk y las costas del mar de Ojotsk y en Kamchatka se da un progresivo reemplazo entre esta subespecie y tytleri en favor de una u otra según la dirección a la que se apunte.[4] Se hibrida con tytleri en la región del río Amur. Se cree que las dos subespecies del este de Asia estuvieron alguna vez geográficamente separadas, pero los sitios de nidificación proporcionados por la expansión humana permitieron que sus áreas de distribución se superpusieran.[6] Su principal área de distribución reproductiva es Japón y Corea. Las poblaciones que se reproducen en los Himalayas centrales y orientales —donde está presente solo en colonias muy separadas—[8] han sido incluidas en gutturalis,[21] si bien ha habido desacuerdos sobre su identidad y fueron descriptas por Stresemann (1940) como una subespecie aparte: ambigua.[4][8][17] La subespecie gutturalis parece ser un visitante mucho más común en el Sudeste Asiático que tytleri y se adentra más hacia el sur.[4] También migra a las Filipinas, a la India (principalmente al este y al sur) y a Sri Lanka.[8] H. r. gutturalis es un divagante ocasional en Alaska y Washington,[22] pero es fácilmente distinguible de la subespecie americana, H. r. erythrogaster, por el tono rojizo de las partes inferiores de esta última.[6]
- H. r. tytleri, descrita por Thomas C. Jerdon en 1864 y bautizada en honor al soldado, naturalista y fotógrafo británico Robert Christopher Tytler,[4][16] presenta partes inferiores casi uniformemente castañas[4] y una franja pectoral incompleta. La cola es más larga.[23] Se reproduce en el sur de Siberia desde la cuenca del río Angará hacia el este hasta Yakutsk y el río Oliokma y hacia el sur hasta el norte de Mongolia. Pasa el invierno en el sudeste asiático, hacia el sur hasta el oeste de Malasia, y en la parte oriental del subcontinente indio. Parece ser más rara en la China mediterránea, país en el que ha sido observada principalmente migrando hacia el noreste y sobre las costas.[4]
- H. r. erythrogaster, la subespecie americana, fue descrita por Pieter Boddaert en 1783.[4][16] Difiere de la subespecie europea en que sus partes inferiores son más rojizas y su banda pectoral azul es más angosta, a menudo incompleta. Se reproduce a lo largo de América del Norte, desde Alaska hasta el sur de México, y migra a las Antillas Menores, Costa Rica, Panamá y Sudamérica para pasar el invierno.[14] Algunas pasan el invierno en las regiones más meridionales de su área de reproducción. La ruta migratoria de esta subespecie se estrecha en Centroamérica, por lo que es abundante durante la migración en las tierras bajas de ambas costas.[24]
Según el autor que se siga, saturata ha sido considerada sinónimo de erythrogaster, gutturalis o tytleri.[4][16] Por su parte, kamtschatica y mandschurica han sido vinculadas a erythrogaster, a saturata —ella misma no reconocida universalmente— y a gutturalis.[4][16] Por último, se ha tomado a ambigua como sinónimo de la subespecie nominal[8] y de gutturalis, siendo la segunda opción la más aceptada.[4]
Hábitat y área de distribución
_in_AP_W_IMG_3861.jpg.webp)
El hábitat preferido de la golondrina común es campo abierto con vegetación baja, como pasturas, prados y terrenos agrícolas, preferentemente con agua en las cercanías. Esta golondrina evita áreas de vegetación densa, empinadas o muy urbanizadas. La presencia de estructuras abiertas y accesibles como graneros, establos y tajeas que proporcionen sitios de nidificación y de ubicaciones expuestas como cables, bordes de techos y ramas desnudas para posarse también son importantes en la elección de esta ave de sus lugares de reproducción.[9][11] Durante la migración, tiende a volar sobre áreas abiertas, a menudo cerca del agua o bordeando cadenas montañosas.[11]
Se reproduce en la región holártica,[11] desde el nivel del mar hasta típicamente los 2700 m,[3] pero hasta los 3000 m en el Cáucaso[9] y América del norte,[11] y solo está ausente en los desiertos y las regiones frías más septentrionales de los continentes. Su área de distribución reproductiva incluye América del norte, norte de Europa, Asia norcentral, norte de África, Medio Oriente, sur de China y Japón; migra a Sudamérica, Asia meridional, Indonesia y Micronesia.[11] Las aves que se reproducen en el este de Asia pasan el invierno a lo largo del Asia tropical, desde la India y Sri Lanka hasta Indonesia y Nueva Guinea. Cantidades cada vez mayores migran al norte de Australia.
En gran parte de su área de distribución, evita las ciudades y en Europa es remplazada en las áreas urbanas por el avión común (Delichon urbicum). Sin embargo, en Honshū, la golondrina común es un ave más bien urbana, reemplazándola la golondrina dáurica (Cecropis daurica) en áreas rurales.[6] Se ha registrado la reproducción de la golondrina común en las regiones más templadas de su área de distribución invernal, como las montañas de Tailandia y Argentina central.[6][25]
Los individuos tienden a regresar a pasar el invierno al mismo lugar cada año.[26] En invierno, la golondrina común es cosmopolita en cuanto a su elección de hábitats, evitando solo los bosques densos y los desiertos.[27] Es más común en hábitats abiertos y de vegetación baja, como la sabana y los ranchos, y en Venezuela, Sudáfrica y Trinidad y Tobago fue descrita como particularmente apegada a campos de caña de azúcar quemados o ya cosechados.[14][28][29]
En África tropical y austral, esta especie es altamente gregaria y se congrega en dormideros de hasta varios millones de individuos en carrizales, campos de caña de azúcar, maíz, hierba de elefante (Pennisetum spp.) o formaciones vegetales de estructura similar.[30] Por ejemplo, el pequeño carrizal de Mount Moreland, cerca de Durban en el este de Sudáfrica, atrae cada noche a más de tres millones de golondrinas, más del 8 % de los ejemplares que se reproducen en Europa.[7] También los hay de solo cientos de individuos. Normalmente, los dormideros se ubican en hábitats muy abiertos, en los que no haya obstáculos que dificulten el ascenso y descenso de las golondrinas.[30] En ausencia de dormideros adecuados, en ocasiones descansan sobre cables donde están más expuestas a los depredadores.[31]
El 23 de diciembre de 1912 se registró el primer caso de una golondrina común que había migrado desde el Reino Unido a Sudáfrica; el ejemplar había sido anillado en Staffordshire y fue recuperado en Natal.[32] Estudios isotópicos han demostrado que durante el invierno las diferentes poblaciones se alimentan en hábitats distintos: las aves que se reproducen en el Reino Unido se alimentan mayormente sobre llanuras, mientras que los ejemplares suizos hacen un mayor uso de los bosques.[33] Otro estudio isotópico dio como resultado que una misma población reproductiva de Dinamarca pasaba el invierno en dos áreas distintas.[34] Tal como habría de esperarse para un migrador de larga distancia, esta ave ha sido encontrada como divagante en puntos tan distantes como Hawái, las Bermudas, Groenlandia, Tristán de Acuña y las Malvinas.[6]
Comportamiento
Se trata de una especie diurna y migratoria. A menudo son observadas en grandes grupos posados sobre cables de teléfono u otras estructuras elevadas. Anidan en colonias, probablemente debido a la distribución de los sitios de nidificación de alta calidad. Emplean tanto vocalizaciones como lenguaje corporal (posturas y movimientos) para comunicarse. Cantan individualmente y a coro. Presentan una amplia variedad de llamados utilizados en diferentes situaciones: llamadas de alarma en presencia de depredadores, llamadas de cortejo y llamadas para los polluelos en el nido. Los polluelos emiten un trino débil cuando piden alimento. Las golondrinas comunes también producen chasquidos cerrando las mandíbulas rápidamente.[11]
En un dormidero de gran tamaño en Cross River (Nigeria), las golondrinas comenzaban a producir chasquidos con sus picos y a gorjear veinte minutos antes de la salida del sol. En los siguientes minutos, la intensidad de las vocalizaciones aumentaba. Muchos ejemplares comenzaban a arreglar su plumaje, a extender las alas y a aletear. Las primeras golondrinas en partir normalmente lo hacían entre cero y nueve minutos antes de la salida del sol (en pocas ocasiones, entre uno y tres minutos después de la salida del sol), para ser seguidas por sus congéneres de manera masiva en promedio cinco minutos después. La partida de todas las golondrinas del dormidero tomaba entre veinte y cuarenta y nueve minutos, aunque el 90% de ellas lo abandonaba en menos de diez minutos. La mayoría de las golondrinas salían volando hacia arriba, a toda velocidad y tan verticalmente como les fuera posible, describiendo a veces ceñidas espirales para ganar altura. Las bandadas se dispersaban cuando ya se habían distanciado del dormidero. Por la tarde, las aves se arremolinaban en densas masas hasta que atardeciera, a gran altura debido a la presencia de alcotanes africanos (Falco cuvierii), y luego se apresuraban a descender masiva y abruptamente en una especie de tornado invertido. La mayor parte de ellas intentaba asegurarse una posición lo más alta posible sobre la colina en la que se encontraba el dormidero.[30]
Alimentación
La golondrina común tiene hábitos similares a otros insectívoros aéreos, incluyendo otras especies de golondrinas y los filogenéticamente desvinculados apódidos. No es un volador particularmente veloz, con una velocidad estimada de unos once metros por segundo, de hasta veinte metros por segundo, y un batimiento alar de aproximadamente cinco y hasta siete o nueve veces por segundo,[35][36] pero tiene la maniobrabilidad suficiente para atrapar insectos en vuelo. A menudo se la ve volando a alturas relativamente bajas en áreas abiertas o semiabiertas.
La golondrina común se alimenta típicamente a siete u ocho metros por encima de aguas poco profundas o del suelo,[9] a menudo siguiendo animales, humanos o maquinaria agrícola para atrapar los insectos ahuyentados,[9][11] pero ocasionalmente captura sus presas de la superficie del agua, paredes o plantas.[9] Los dípteros (moscas y mosquitos, entre otros), ortópteros (saltamontes y grillos), anisópteros (libélulas), coleópteros (escarabajos), heteróceros (polillas) y otros insectos voladores componen el 99 % de su dieta.[11] En su área de distribución reproductiva, los dípteros de gran tamaño constituyen alrededor del 70 % de su dieta, siendo los áfidos otro componente significativo. Sin embargo, en Europa, las golondrinas comunes consumen menos áfidos que el avión común (Delichon urbicum) y el avión zapador (Riparia riparia).[9] En sus cuarteles de invernada, los himenópteros, especialmente las hormigas voladoras, representan una importante fuente de alimento. Durante el período de puesta de los huevos, las golondrinas comunes cazan en pareja, pero en otras circunstancias a menudo forman grandes bandadas.[6]
La golondrina común bebe rozando el agua de lagos o ríos en vuelo.[11] Se baña dando chapuzones en el agua por un instante mientras vuela.[26] También es capaz de alimentar a sus polluelos mientras vuela.[11]
Un estudio en Virginia Occidental encontró que las golondrinas comunes se alimentaban dentro de un radio de 1,2 km de sus nidos. En Europa, se alimentan solo alrededor de unos 500 m desde sus nidos.[11] Tras la temporada reproductiva, se reúnen en dormideros comunales. Los carrizales suelen ser un lugar privilegiado en la elección de las golondrinas, que se arremolinan masivamente antes de descender sobre los carrizos.[13] Los carrizales son una importante fuente de alimento antes de y durante la migración. Si bien la golondrina común es un migrador diurno que puede alimentarse en vuelo mientras viaja a bajas alturas sobre el suelo o el agua, los carrizales les permiten establecer o reabastecer sus reservas de grasa.[37] Las golondrinas muestran un aumento de peso de entre 2 y 4 gramos justo antes de migrar al norte hacia su área de distribución reproductiva, normalmente al final del ciclo de muda del plumaje.[30]
Reproducción
Las golondrinas comunes se reproducen generalmente entre mayo y agosto, pero esto varía mucho según el lugar. Pueden reproducirse ya en la primera temporada reproductiva posterior a su nacimiento. Generalmente, las golondrinas jóvenes no depositan tantos huevos como las adultas.[11]
Estas aves asedian a intrusos tales como los gatos y aves del género Accipiter que se aventuran en las cercanías del nido, volando muy cerca de estos depredadores.[38] El parasitismo de puesta por parte de tordos cabecicafé (Molothrus ater) en América del norte y de los cucos en Eurasia es infrecuente.[9][11] Al menos en lo que respecta al tordo cabecicafé, se han registrado casos en los que el polluelo parásito fue criado exitosamente y logró abandonar el nido, lo que demuestra que la golondrina común es un huésped adecuado para esta especie.[11]
Se ha registrado la hibridación de la golondrina común con la golondrina gorginegra (Petrochelidon pyrrhonota) y la golondrina de cueva (Petrochelidon fulva) en América del norte, y con el avión común (Delichon urbicum) en Eurasia. De este último entrecruzamiento resulta uno de los híbridos paseriformes más comunes.[38]
Emparejamiento
Los machos regresan a las áreas de reproducción antes que las hembras y escogen un sitio para el nido, que es anunciado a las hembras con un vuelo en círculo y el canto. También intentan atraerlas desplegando sus colas. Las hembras se sienten particularmente atraídas hacia los machos con alas y colas simétricas. Los machos que exhiben un mayor grado de simetría consiguen compañera más rápidamente que los asimétricos. La asimetría puede ser el resultado de factores genéticos como la endogamia y las mutaciones o de factores ambientales como la deficiencia alimentaria, la infección de parásitos o la presencia de patógenos. Los individuos afectados por estos factores no solo presentan un mayor grado de asimetría sino también una menor fortaleza y longevidad. Por lo tanto, las hembras que escogen machos simétricos estarían seleccionando machos superiores.[11]
El éxito reproductivo de los machos también está relacionado con la longitud de su cola, siendo las más largas más atractivas para las hembras.[9][11][38][39] Los machos con las plumas de la cola más largas son generalmente más longevos y más resistentes a las enfermedades, por lo que las hembras que los escogen obtienen una ventaja reproductiva indirecta de esta elección, pues plumas largas en la cola señalan la superioridad genética de un individuo que producirá una descendencia más vital.[11][38] Los machos con colas largas también tienen manchas blancas más grandes en la cola y, debido a que los piojos aviares prefieren las plumas blancas, la presencia de estas grandes manchas blancas sin daños producidos por los parásitos da indicios de la calidad reproductiva de un individuo; existe una relación positiva entre el tamaño de las manchas y el número de descendientes producidos por temporada.[40]
Los machos del norte de Europa tienen colas más largas que los del sur; mientras que en España las colas de los machos son solo un 5 % más largas que las de las hembras, en Finlandia la diferencia es de un 20 %. En Dinamarca, la longitud promedio de la cola del macho se incrementó en un 9 % entre 1984 y 2004, pero es posible que en el futuro los cambios climáticos lleven al desarrollo de colas más cortas a medida que los veranos se vuelvan más cálidos y secos.[41] También existe evidencia de que los machos tienden a escoger compañeras con colas largas.[11]
Los individuos de ambos sexos defienden el nido, pero los machos son particularmente agresivos y territoriales.[6] Las parejas que se han reproducido exitosamente pueden durar varios años.[11] Por otro lado, las cópulas fuera de la pareja son comunes, por lo que esta especie es genéticamente polígama, aunque sea socialmente monógama.[11][42] Los machos vigilan a las hembras activamente para evitar ser “engañados”[43] y pueden emplear llamadas de alarma falsa para interrumpir intentos de cópulas entre sus parejas y otros machos.[44]
Los machos sin compañera a menudo se asocian con alguna pareja durante incluso toda la temporada reproductiva. Aunque estos «colaboradores» no suelen alimentar a los polluelos, pueden ayudar en la construcción y defensa del nido, incubación de los huevos y empollando a los pequeños. Los colaboradores son predominantemente machos.[11]
Construcción del nido

La golondrina común anida típicamente en el interior de edificaciones accesibles como graneros y establos y debajo de puentes y embarcaderos. Antes de que hubiera abundancia de estructuras construidas por el ser humano, las golondrinas comunes anidaban sobre acantilados o en cuevas, pero esto es raro en la actualidad.
El prolijo nido en forma de taza es ubicado sobre una viga o contra una proyección vertical adecuada.[8] Es construido por ambos miembros de la pareja, aunque más a menudo por la hembra, siendo recolectadas las bolitas de barro en sus picos y recubiertas con pasto, plumas, algas de agua dulce y marinas y otros materiales suaves.[6][11][45] Las golondrinas comunes pueden anidar en colonias en donde haya una suficiente cantidad de sitios de nidificación de alta calidad y, dentro de la colonia, cada pareja defiende un territorio alrededor del nido, que, en Europa, es de entre cuatro y ocho metros cuadrados.[11]
En América del norte al menos, las golondrinas comunes se involucran con frecuencia en una relación mutualista con las águilas pescadoras (Pandion haliaetus). Las golondrinas construyen sus nidos debajo de los de las águilas pescadoras, obteniendo así protección de otras aves rapaces que son ahuyentadas por esta águila que se alimenta exclusivamente de peces. A su vez, los llamados de alerta de las golondrinas alertan a las águilas pescadoras de la presencia de estos depredadores.[11]
Nidada y cría de los polluelos
Las hembras depositan de dos a siete huevos ―por lo general, cuatro o cinco―, blancos con manchas rojizas. Los huevos miden 20 por 14 milímetros y pesan 1,9 gramos, de los que el 5 % corresponden al cascarón. En Europa, la hembra se encarga de casi toda la incubación, pero en América del norte el macho puede incubar hasta el 25 % del tiempo. El periodo de incubación es normalmente de entre 14 y 19 días, a los que se suman otros 18 a 23 días antes de que los polluelos altriciales abandonen el nido. Los pequeños ya voladores permanecen junto a sus padres y son alimentados por ellos durante alrededor de una semana más.[6] Para la segunda semana después de haber dejado el nido, los polluelos ya se han dispersado y a menudo viajan a otras colonias de golondrinas. Ambos padres alimentan y protegen a los pequeños y remueven los sacos fecales del nido, si bien las hembras proporcionan un mayor cuidado parental. Los padres pueden alimentar a los polluelos en el nido hasta 400 veces por días. Las golondrinas comunes alimentan a sus pequeños con insectos comprimidos en una bolita, que es transportada al nido en la garganta del adulto.[11] En ocasiones, los polluelos de la primera nidada ayudan en la alimentación de los de la segunda.[6][11]
Normalmente crían dos nidadas por temporada.[6][11] El nido en el que criaron a la primera es reutilizado para la segunda y reparado y vuelto a usar en años siguientes.[6] La supervivencia de los nidos durante 10 o 15 años es común con el mantenimiento adecuado y se registró uno que fue ocupado durante 48 años.[10]
El porcentaje de los polluelos que rompen el cascarón es de un 90 %. La mortalidad promedio es de entre un 70 y un 80 % en el primer año y de un 40 a 70 % para los adultos.[6] Aunque la edad récord sea de más de 11 años, la mayoría no supera los cuatro.[6][11] Los polluelos tienen bocas rojas y prominentes, un rasgo que según se ha demostrado induce a los padres a alimentarlos. La intensidad del color de sus bocas está relacionado con su inmunocompetencia y los polluelos de nidadas de gran tamaño tienen bocas menos vívidas.[46]
Depredadores y parásitos
Las golondrinas comunes (y otros paseriformes pequeños) a menudo presentan característicos huecos en las plumas de las alas y la cola. Se sugirió que estos huecos serían ocasionados por piojos aviares tales como Machaerilaemus malleus y Myrsidea rustica. Sin embargo, otros estudios apuntan a que podrían ser causados por ischnóceros del género Brueelia. Entre las especies de piojos de las que la golondrina común es un huésped se encuentran Brueelia domestica y Philopterus microsomaticus.[47][48] En Texas, Oeciacus vicarius, que a menudo infesta al avión roquero americano (Petrochelidon pyrrhonota), también ha sido descubierto sobre la golondrina común.[49]
Los dormideros comunales atraen a los depredadores: numerosas especies de falcónidos y accipítridos aprovechan estas oportunidades.[30] Entre sus depredadores se encuentran el ser humano —la especie que causa la mayor cantidad de bajas a la golondrina común—,[30] gavilán chikra (Accipiter badius),[30] gavilán de Cooper (Accipiter cooperii),[11] gavilancito chico (Accipiter minullus),[30] esparvero chico (Accipiter striatus),[11] azor de Toussenel (Accipiter toussenelii),[30] aguilucho lagunero (Circus aeruginosus),[30] cernícalo del Amur (Falco amurensis),[30] cernícalo pizarroso (Falco ardosiaceus),[30] halcón borni (Falco biarmicus),[30] halcón de cuello rojo (Falco chicquera),[30] alcotán africano (Falco cuvierii),[30] halcón peregrino (Falco peregrinus),[30][50] cernícalo americano (Falco sparverius),[11] alcotán europeo (Falco subbuteo),[30] gavilán gabar (Micronisus gabar),[30] milano negro (Milvus migrans ssp. parasitus),[30] aguilucho caricalvo común (Polyboroides typus ssp. pectoralis),[30] tecolote oriental (Megascops asio),[11] autillo europeo (Otus scops),[30] cárabo africano (Strix woodfordii),[30] lechuza de los campanarios (Tyto alba),[30] cuco senegalés (Centropus senegalensis),[30] gaviotas (Laridae), zanate cola de bote (Quiscalus major), zanate norteño (Quiscalus quiscula), gato doméstico (Felis silvestris), lince rojo (Lynx rufus), comadrejas (Mustela spp.), mapache (Procyon lotor), rata parda (Rattus norvegicus), ardillas (Sciuridae), serpientes, etc.[11] Se sabe que los murciélagos depredadores tales como el falso vampiro mayor (Megaderma lyra) se alimentan de golondrinas comunes.[51] La mayoría de los depredadores ataca a los polluelos, pero los gavilanes, halcones y búhos cazan a los adultos. Las golondrinas comunes evaden a los depredadores con su vuelo ágil y construyendo sus nidos en lugares de difícil acceso para ellos.[11]
La partida y el regreso de las golondrinas de y a sus dormideros están altamente sincronizados y son previsibles. Más del 90 % de las aves de un dormidero de 1,5 millones de ejemplares en Nigeria salía y volvía dentro de los diez minutos alrededor de la salida y de la puesta del sol respectivamente, en masas densas y de rápido movimiento. Los alcotanes africanos sincronizaban su llegada con la actividad de las golondrinas, arribando el primero de ellos entre 2 y 14 minutos antes de la partida o del regreso del primer individuo, y se iban marchando una vez que la mayoría de las golondrinas se hubiera ido. Al atardecer, los primeros llegaban al dormidero entre 15 y 20 minutos antes de la puesta del sol, cuando normalmente las golondrinas aún estaban ausentes. Las golondrinas intentaban reducir el riesgo de depredación congregándose en enormes cantidades, acotando su partida y regreso a un pequeño lapso y realizando ascensos y descensos rápidos y sincronizados para evitar a los depredadores.[30]
Cuando las golondrinas regresaban al atardecer, reaccionaban a la presencia de alcotanes ganando altura en el aire y compactándose en densas masas, por lo que los alcotanes debían entonces volar más alto que al amanecer para acercárseles. La caza por parte de estos depredadores temprano en la mañana era muy diferente, pues las golondrinas debían ascender desde el nivel del suelo y respondían a la presencia de alcotanes volando bajo abandonando el dormidero a alta velocidad y no alejándose de los contornos de la hierba de elefante hasta que alcanzaran el borde de zonas arboladas. Estas bandadas eran atacadas a través de la persecución o en cortos y repetidos picados, lo que causaba que las golondrinas se dispersaran en todas direcciones. Cuando las golondrinas abandonaban el dormidero en ascenso vertical, los alcotanes las atacaban cayendo sobre ellas desde más arriba, virando hacia arriba después de cada picado.[30]
Tratándose de cazadores aéreos, los alcotanes solo podían atacar efectivamente a las golondrinas cuando estas se encontraban presentes en el dormidero y en el aire, es decir, en el reducido lapso de 30 minutos alrededor de la puesta y de la salida del sol. Sin embargo, parecían evitar en gran medida las arremetidas contra grandes masas, las que quizá les resultaran difíciles de abordar. Por lo general, ignoraban las enormes cantidades que partían masivamente, a gran velocidad y en dirección ascendente al amanecer y las que se congregaban a gran altura al atardecer para luego caer en picada sobre el dormidero, concentrándose en individuos rezagados y en pequeños grupos, tempranos o tardíos. Si bien se lanzaban a la caza de individuos aislados y de bandadas de entre dos y 15 000 individuos, eran más exitosos cuando se embarcaban en la cacería de grupos de menos de 50 individuos. El riesgo de depredación para las golondrinas tomadas individualmente decrecía dramáticamente con el tamaño de la bandada.[30]
Estado de conservación
La golondrina común no se aproxima siquiera al umbral los criterios de superficie del área de distribución y de tamaño de la población para especies vulnerables de Lista Roja de la UICN: tiene una enorme área de distribución que se extiende sobre unos 43,4 millones de kilómetros cuadrados y se estima que su población global supera los 190 millones de individuos. A pesar de que la tendencia poblacional parece ser negativa, se cree que esta disminución en sus números no es lo suficientemente rápida como para que la especie se acerque siquiera al umbral del criterio de decrecimiento poblacional para especies vulnerables (esto es, un decrecimiento de más del 30 % en diez años o tres generaciones).[2] Por estas razones, la golondrina común fue calificada como «bajo preocupación menor» en la Lista Roja de 2012,[3] y no tiene un estatus especial bajo la Convención sobre el Comercio Internacional de Especies Amenazadas de Fauna y Flora Silvestres.[11]
Históricamente, esta especie se ha beneficiado de la tala de los bosques, que crea los hábitats abiertos que prefiere, y de la presencia humana, que le proporciona abundancia de sitios de nidificación seguros. Ha habido decrecimientos locales debido al uso de DDT en Israel en la década de 1950, a causa de la competencia por los sitios de nidificación con el gorrión común (Passer domesticus) en Estados Unidos en el siglo XIX y por la intensificación de la agricultura, que reduce la disponibilidad de insectos, en partes de Europa y Asia en la actualidad. Sin embargo, se ha registrado un incremento en América del norte durante el siglo XX debido a la mayor disponibilidad de sitios de nidificación y a la subsecuente expansión en su área de distribución, incluyendo la colonización del norte de Alberta.[6] En 2006, se afirmó que las poblaciones de golondrinas en el Reino Unido habían estado decayendo, probablemente debido a la sequía y al uso de pesticidas en su ruta migratoria y a la transformación a la que estaban sujetas las edificaciones rurales y otros sitios de nidificación favorecidos en el propio Reino Unido.[7] Por otro lado, según información publicada en 2007 y 2008 respectivamente, la especie habría sufrido un decrecimiento poblacional pequeño o estadísticamente insignificante en los anteriores 40 años en América del norte y se habría mantenido estable en Europa desde 1980.[2][3] Otras fuentes, sin embargo, indican que en el periodo comprendido entre 1996 y 2021 su población ha decrecido hasta el 45%, al acabar con sus puntos de cría en las ciudades, la competencia de aves invasoras como la cotorra y la disminución de ciertos insectos de los que se alimenta. [52]
El cambio climático puede afectar a las golondrinas comunes.[41] Las golondrinas en África y durante su migración exhiben grandes variaciones en su masa corporal y reservas de grasa dependiendo de las condiciones atmosféricas con las que se encuentren.[30] La sequía ocasiona pérdida de peso y una lenta renovación de las plumas, y la expansión del Sahara hará de este desierto un obstáculo aún mayor para las aves europeas.[41] En un período de sequía, las golondrinas que pasaban el invierno en Botsuana mostraron bajo peso y nada de grasa, mientras que años de abundantes precipitaciones ―y, por lo tanto, de gran disponibilidad de insectos― resultaron en mayores pesos y reservas de grasa. De manera similar, en Nigeria en febrero de 2001, el soplido del Harmattan coincidió con un marcado descenso en la masa corporal y con una falta de grasa en las golondrinas, registrándose pesos más elevados tanto antes como después del levantamiento de ese viento seco.[30] A su vez, los veranos cálidos y secos reducirán la disponibilidad de insectos con los que alimentar a lo polluelos. Inversamente, las primaveras más cálidas podrían alargar su temporada reproductiva y contribuir a la producción de una mayor cantidad de polluelos.[41] En 2014, la golondrina común fue declarada Ave del Año por SEO/BirdLife, la entidad científica dedicada al estudio y conservación de las aves en España desde 1954. Según SEO/BirdLife, sus datos de seguimiento de poblaciones por todo el territorio indican que la golondrina ha perdido el 30 % de su población en España en la última década,[53] debido entre otros motivos al uso de pesticidas en el campo, el abandono agrario, la destrucción de edificios tradicionales y los nuevos modos de arquitectura.
Relación con el ser humano y referencias culturales
_-_Hirundo_rustica_(030b).jpg.webp)
En el Viejo Mundo, las golondrinas comunes parecen haber hecho uso de los puentes y otras estructuras construidas por el ser humano desde tiempos inmemoriales.[54] Puede encontrarse una referencia temprana a esta ave en las Geórgicas de Virgilio (29 a. C.): […] garrula quam tignis nidum suspendat hirundo. («[…] la golondrina gorjeadora cuelga su nido de las vigas».).[55] Se cree que las golondrinas comunes comenzaron a adosar sus nidos a las construcciones de los nativos americanos a comienzos del siglo XIX. Es probable que la consiguiente expansión del ser humano en América del norte resultara en una dramática diseminación de la especie a lo largo del continente.[56]
Existen muchas referencias a la migración al norte de las golondrinas comunes como “ícono de la primavera y epítome del verano” en el hemisferio septentrional.[7] La golondrina común simboliza la llegada de la primavera y el amor en Pervigilium Veneris, un poema latino tardío. En La tierra baldía, T. S. Eliot citó la línea Quando fiam uti chelidon [ut tacere desinam]? («¿Cuándo seré como una golondrina, así podré dejar de estar en silencio?»). Esto refiere a una versión del mito de Filomela en la que ella se convierte en un ruiseñor y su hermana Procne en una golondrina; en versiones menos comunes, las especies aparecen invertidas.[57] De manera inversa, la imagen de una bandada de golondrinas volando en su migración al sur concluye la oda «Al Otoño» de John Keats:
¿Dónde están las canciones de primavera? ¡Ah! ¿Dónde?
Ni pienses más en ellas, pues ya tienes tu música,
y únense golondrinas, gorjeando, en el cielo.
cuando estriadas nubes florecen el suave
morir del día y tiñen de rosa los rastrojos;
entonces el doliente coro de los mosquitos
entre sauces del río se lamenta, elevándose
o bajando, según el soplar de la brisa;
y balan los crecidos corderos en los montes;
canta el grillo en el seto; y ya, con trino blando,
en el jardín cercado, el petirrojo silba
En el Evangelio apócrifo del Pseudo Tomás, se cuenta que Jesús, con cinco años, "se encontraba un día jugando en el cauce de un arroyo... hizo una masa blanda de barro y formó con ella doce golondrinas".[58]
Las golondrinas son mencionadas en varias obras de William Shakespeare por su ligereza en vuelo; por ejemplo True hope is swift, and flies with swallow's wings […] (“La verdadera esperanza es veloz y vuela con alas de golondrina […]”) del quinto acto de Ricardo III. Shakespeare también se refiere a la migración anual de la golondrina en The Winter's Tale, cuarto acto: Daffodils, That come before the swallow dares, and take The winds of March with beauty […] (“Los narcisos, Que llegan antes de que las golondrinas se atrevan y toman Los vientos de marzo con belleza […]”).
Las golondrinas, utilizando un recurso literario como el hipérbaton, aparecen en una de las Rimas más famosas de Gustavo Adolfo Bécquer.
Volverán las oscuras golondrinas
en tu balcón sus nidos a colgar,
y otra vez con el ala a sus cristales
jugando llamarán.Pero aquellas que el vuelo refrenaban
tu hermosura y mi dicha a contemplar,
aquellas que aprendieron nuestros nombres...
ésas... ¡no volverán!Volverán las tupidas madreselvas
de tu jardín las tapias a escalar,
y otra vez a la tarde aún más hermosas
sus flores se abrirán.Pero aquellas cuajadas de rocío
cuyas gotas mirábamos temblar
y caer como lágrimas del día...
ésas... ¡no volverán!Volverán del amor en tus oídos
las palabras ardientes a sonar;
tu corazón de su profundo sueño
tal vez despertará.Pero mudo y absorto y de rodillas,
nadie así te amará.
como se adora a Dios ante su altar,
como yo te he querido..., desengáñate,
Oscar Wilde tiene como coprotagonista a una golondrina en su cuento El príncipe feliz. Y el poeta Swinburne interpela a la golondrina en su poema "Itylus", aludiendo una versión tebana del mito griego de Aedón o Edón donde Quelidón o Quelidonia es transformada en golondrina y Aedón en ruiseñor.[59]
El poeta Eduardo Fraile publicó su libro Balada de las golondrinas (Pre-Textos, 2009) en homenaje a las golondrinas de su pueblo, Castrodeza.[60]
Gilbert White estudió a la golondrina común en detalle en su obra The Natural History of Selborne, pero incluso este atento observador no estaba seguro de si las golondrinas migraban o hibernaban en invierno.[61] Las golondrinas comunes consumen enormes cantidades de insectos y son importantes en el control de sus poblaciones.[11] En el pasado, la tolerancia a este insectívoro beneficioso se vio reforzada por supersticiones en torno al daño a sus nidos. Se creía que dicho acto podría llevar a que las vacas produjeran leche con sangre o dejaran de producirla en absoluto o a que las gallinas dejaran de poner huevos.[10] Por otro lado, en Nigeria, son cazadas a cientos de miles por año, lo que hace del ser humano el principal depredador de esta especie por un amplio margen.[30] La golondrina común es el ave nacional de Estonia.[62]
Notas y referencias
- BirdLife International (2016). «Hirundo rustica». Lista Roja de especies amenazadas de la UICN 2017-1 (en inglés). ISSN 2307-8235. Consultado el 8 de julio de 2017.
- «BirdLife International Species factsheet: Hirundo rustica» (en inglés). BirdLife International. Consultado el 17 de septiembre de 2012.
- «Barn Swallow (Hirundo rustica) – BirdLife species facsheet» (en inglés). BirdLife International. Consultado el 17 de septiembre de 2012.
- Dickinson, Edward C.; Dekker, René W. R. J. (2001). «Systematic notes on Asian birds. 13. A preliminary review of the Hirundinidae». Zoologische Verhandelingen (en inglés) (Leiden: Rijksmuseum van Natuurlijke Historie) 335: 127-144. ISSN 0024-1652. Consultado el 18 de septiembre de 2012.
- Bernis, F; De Juana, E; Del Hoyo, J; Fernández-Cruz, M; Ferrer, X; Sáez-Royuela, R; Sargatal, J (2004). «Nombres en castellano de las aves del mundo recomendados por la Sociedad Española de Ornitología (Novena parte: Orden Passeriformes, Familias Cotingidae a Motacillidae)». Ardeola. Handbook of the Birds of the World (Madrid: SEO/BirdLife) 51 (2): 491-499. ISSN 0570-7358. Consultado el 11 de junio de 2018.
- Turner, Angela K.; Rose, Chris (1989). Swallows & martins: an identification guide and handbook (en inglés). Boston: Houghton Mifflin. pp. 164-169. ISBN 0-395-51174-7.
- «World Cup airport 'threatens swallow population'». The Guardian (en inglés). 16 de noviembre de 2006. Consultado el 17 de septiembre de 2012.
- Vaurie, Charles (23 de julio de 1951). «Notes on some Asiatic swallows». American Museum Novitates (en inglés) (Nueva York: American Museum of Natural History) 1529: 1-47. Consultado el 19 de septiembre de 2012.
- Snow, David W.; Perrins, Christopher M., eds. (1998). The birds of the Western Palearctic. Volume 2, Passerines (en inglés). Edición concisa basada en The Handbook of the birds of Europe, the Middle East and North Africa. Oxford: Oxford University Press. pp. 1061-1064. ISBN 0-19-854099-X.
- Cocker, Mark; Mabey, Richard (2005). Birds Britannica (en inglés). Londres: Chatto & Windus. ISBN 0-7011-6907-9.
- Dewey, Tanya; Roth, Chava (2002). «ADW: Hirundo rustica: Information». Animal Diversity Web (en inglés). Míchigan: University of Michigan Museum of Zoology. Consultado el 20 de septiembre de 2012.
- Barlow, Clive; Wacher, Tim (1997). A Field Guide to birds of The Gambia and Senegal (en inglés). Ilustrado por Tony Disley. Robertsbridge: Pica Press. p. 279. ISBN 1-873403-32-1.
- Svensson, Lars; Grant, Peter (1999). Collins Bird Guide (en inglés). Ilustrado por Killian Mullarney y Dan Zetterström. Londres: HarperCollins. p. 242. ISBN 0-00-219728-6.
- Hilty, Steven L. (2003). Birds of Venezuela (en inglés). Londres: Christopher Helm. p. 691. ISBN 0-7136-6418-5.
- Linneo, Carlos (1758). Laurentii Salvii, ed. Systema naturæ per regna tria naturæ, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis (en latín). Tomo 1 (décima edición). Salvius publ. p. 191. Consultado el 17 de septiembre de 2012.
- Dickinson, Edward C.; Eck, Siegfried; Milensky, Christopher M. (2002). «Systematic notes on Asian birds. 31. Eastern races of the barn swallow Hirundo rustica Linnaeus, 1758». Zoologische Verhandelingen (en inglés) (Leiden: Rijksmuseum van Natuurlijke Historie) 340: 201-203. ISSN 0024-1652. Consultado el 17 de septiembre de 2012.
- Stresemann, E. (1940). «Welche Rasse von Hirundo rustica bretet in Sikkim?». Ornithologischen Monatsbericht (en alemán) 48 (3): 88-89.
- Whistler, H. (1940). «The Common Swallow Hirundo rustica rustica in Ceylon». British Ornithologists' Union (en inglés) (Ibis) 82 (3): 539.
- Oatley, Terry B. (noviembre de 1996). «Bird ringing across the world». Euring Newsletter - Volumen 1 (en inglés). Euring. Archivado desde el original el 15 de noviembre de 2012. Consultado el 20 de septiembre de 2012.
- Dekker, René W. R. J. (octubre de 2003). «Type specimens of birds in the National Museum of Natural History, Leiden. Part 2: Passerines: Eurylaimidae - Eopsaltriidae (Peter’s sequence)». NNM Technical Bulletin (en inglés) (Leiden: Nationaal Natuurhistorisch Museum) 6: 20. ISSN 1387-0211. Consultado el 20 de septiembre de 2012.
- Whistler, H. (1937). «The breeding Swallow of the Western Himalayas». British Ornithologists' Union (en inglés) (Ibis) 79 (2): 413-415.
- Sibley, David (2000). The North American Bird Guide (en inglés). Pica Press. ISBN 1-873403-98-4.
- Rasmussen, Pamela C.; Anderton, John C. (2005). Birds of South Asia: The Ripley Guide (en inglés). Instituto Smithsoniano y Lynx Edicions. ISBN 84-87334-67-9.
- Stiles, Gary; Skutch, Alexander (2003). A guide to the Birds of Costa Rica (en inglés). Ithaca (Nueva York): Cornell University Press. p. 343. ISBN 0-8014-2287-6.
- Lekagul, Boonsong; Round, Philip D. (1991). A Guide to the Birds of Thailand (en inglés). Ilustrado por Mongkol Wongkalasin y Kamol Komolphalin. Bangkok: Saha Karn Baet. p. 234. ISBN 974-85673-6-2.
- Burton, Robert (1985). Bird behaviour (en inglés). Londres: Granada. ISBN 0-246-12440-7.
- Sinclair, Ian; Hockey, Phil; Tarboton, Warwick (2002). SASOL Birds of Southern Africa (en inglés). Ciudad del Cabo: Struik. p. 294. ISBN 1-86872-721-1.
- Froneman, Albert; Bortle, Jon; Merritt, Ron (abril de 2007). «Draft swallow monitoring and bird aircraft interaction». Environmental Impact Assessment Report (en inglés) (Dube TradePort Environmental Impact Assessment Information Center).
- Ffrench, Richard (1991). A Guide to the Birds of Trinidad and Tobago (en inglés) (Segunda edición). Ithaca (Nueva York): Comstock Publishing. pp. 315-316. ISBN 0-8014-9792-2.
- Bijlsma, Rob G.; van den Brink, Bennie (2005). «A Barn Swallow Hirundo rustica roost under attack: timing and risks in the presence of African Hobbies Falco cuvieri». Ardea 93 (1): 37-48. Archivado desde el original el 12 de febrero de 2012. Consultado el 21 de septiembre de 2012.
- George, P. V. (1965). «Swallows Hirundo rustica Linnaeus roosting on wires». Journal of the Bombay Natural History Society (en inglés) (Bombay: Bombay Natural History Society) 62 (1): 160.
- Wernham, Chris, ed. (2002). The Migration Atlas: Movements of the Birds of Britain and Ireland (en inglés). T & AD Poyser. p. 462. ISBN 0-7136-6514-9.
- Evans, Karl L.; Wadron, Susan; Bradbury, Richars B. (2003). «Segregation in the African wintering grounds of English and Swiss Barn Swallows Hirundo rustica: a stable isotope study». Bird Study (en inglés) (Taylor & Francis) 50 (3): 294-299. Consultado el 23 de septiembre de 2012.
- Møller, Anders Pape; Hobson, Keith A. (7 de julio de 2004). «Heterogeneity in stable isotope profiles predicts coexistence of populations of barn swallows Hirundo rustica differing in morphology and reproductive performance». Proceedings of the Royal Society of London, B (en inglés) (Londres: Royal Society) 271 (1546): 1355-1362. ISSN 0962-8452. Consultado el 23 de septiembre de 2012.
- Liechti, Felix; Bruderer, Lukas (15 de agosto de 2002). «Wingbeat frequency of barn swallows and house martins: a comparison between free flight and wind tunnel experiments». The Journal of Experimental Biology (en inglés) (The Company of Biologists) 205 (16): 2461-2467.
- Park, Kirsty; Rosén, Mikael; Hedenström, Anders (1 de agosto de 2001). «Kinematics of the barn swallow (Hirundo rustica) over a wide range of speeds in a wind tunnel». The Journal of Experimental Biology (en inglés) (The Company of Biologists) 204 (15): 2741-2750. ISSN 0022-0949.
- Pilastro, Andrea; Pilastro, Andrea (diciembre de 1998). «The EURING Swallow Project in Italy». Euring Newsletter - Volumen 2 (en inglés). Euring. Archivado desde el original el 3 de diciembre de 2007.
- Møller, Anders Pape (1994). Sexual Selection and the Barn Swallow (en inglés). Ilustrado por Jens Gregersen. Oxford: Oxford University Press. p. 245. ISBN 0-19-854028-0.
- Saino, Nicola; Romano, Maria; Sacchi, Roberto; Ninni, Paola; Galeotti, Paolo; Møller, Anders Pape (2003). «Do male barn swallows (Hirundo rustica) experience a trade-off between the expression of multiple sexual signals?». Behavioral Ecology and Sociobiology (en inglés) (Springer-Verlag) 54 (5): 465-471.
- Kose, Mati; Mänd, Raivo; Møller, Anders Pape (diciembre de 1999). «Sexual selection for white tail spots in the barn swallow in relation to habitat choice by feather lice». Animal Behaviour (en inglés) (Elsevier) 58 (6): 1201-1205.
- Turner, Angela (2010). «Climate change: a Swallow's eye view». British Birds (en inglés) 102 (1): 3-16.
- Møller, Anders Pape; Tegelstrom, Håkan (1997). «Extra-pair paternity and tail ornamentation in the barn swallow Hirundo rustica». Behavioral Ecology and Sociobiology (en inglés) (Springer-Verlag) 41 (5): 353-360.
- Møller, Anders Pape (1985). «Mixed reproductive strategy and mate guarding in a semi-colonial passerine, the swallow Hirundo rustica». Behavioral Ecology and Sociobiology (en inglés) (Springer-Verlag) 17 (4): 401-408.
- Møller, Anders Pape (1990). «Deceptive use of alarm calls by male swallows, Hirundo rustica: a new paternity guard». Behavioral Ecology (en inglés) (International Society for Behavioural Ecology) 1 (1): 1-6.
- Duffin, K. (1973). «Barn Swallows use freshwater and marine algae in nest construction». The Wilson Bulletin (en inglés) (Wilson Ornithological Society) 85: 237-238.
- Saino, Nicola; Ambrosini, Roberto; Martinelli, Roberta; Ninni, Paola; Møller, Anders Pape (2003). «Gape coloration reliably reflects immunocompetence of barn swallow (Hirundo rustica) nestlings». Behavioral Ecology (en inglés) (International Society for Behavioural Ecology) 14 (1): 16-22.
- Møller, Anders Pape (1991). «Parasites, sexual ornaments and mate choice in the Barn Swallow Hirundo rustica». En Loye, J. E.; Zuk, M., ed. Bird-parasite interactions: Ecology, evolution, and behaviour (en inglés). Oxford: Oxford University Press. pp. 328-343.
- Vas, Zoltán; Csörgő, Tibor; Møller, Anders P.; Rózsa, Lajos (diciembre de 2008). «The feather holes on the barn swallow Hirundo rustica and other small passerines are probably caused by Brueelia spp. lice». Journal of Parasitology (en inglés) (American Society of Parasitologists) 94 (6): 1438-1440. ISSN 0022-3395. Archivado desde el original el 30 de julio de 2013. Consultado el 19 de septiembre de 2012.
- Kopachena, Jeffrey G.; Cochran, Beverly L.; Nichols, Tammy B. (diciembre de 2007). «The incidence of American swallow bugs (Oeciacus vicarius) in barn swallow (Hirundo rustica) colonies in northeast Texas». Journal of Vector Ecology (en inglés) 32 (2): 280-284. ISSN 1081-1710.
- Ezaki, Yasuo; Mizota, Hiromi (diciembre de 2006). «Wintering of a Peregrine Falcon on an electricity pylon and its food in a suburban area of western Japan». Ornithological Science (en inglés) (The Ornithological Society of Japan) 5 (2): 211-216.
- Sugathan, R. (1988). «Movement of the Eastern Swallow (Hirundo rustica gutturalis) ringed at Mootpuzha, Kerala». Journal of the Bombay Natural History Society (en inglés) (Bombay: Bombay Natural History Society) 85 (2): 428-429.
- «España se queda sin golondrinas: así se extingue la emblemática ave de Bécquer». www.msn.com. Consultado el 27 de noviembre de 2021.
- «Nos quedamos sin golondrinas». SEO/BirdLife. 27 de marzo de 2014. Consultado el 27 de noviembre de 2021.
- Turner, Angela K. (2006). The Barn Swallow (en inglés). Ilustrado por Ian Rendall. Londres: T. & A. D. Poyser. ISBN 0-7136-6558-0.
- Virgilio (29 a. C.). Geórgicas (en latín). Libro IV, línea 307.
- Williams, Nigel (4 de abril de 2006). «Swallows track human moves». Current Biology (en inglés) (Cell Press) 16 (7): R231.
- Nims, John Frederick (1981). The Harper Anthology of Poetry (en inglés). Nueva York: Harper and Row. ISBN 0-06-044846-6.
- Santos Otero, Aurelio de (1956). «Apócrifos de la infancia». Los Evangelios Apócrifos. Biblioteca de Autores Cristianos. p. 303.
- «Quelidonia o Quelidón». Enciclonet 3.0. Enciclonet. 2019.
- La poesía también sueña con golondrinas heroicas
- Gilbert, White (1789). The Natural History and Antiquities of Selborne (en inglés). Londres: T. Bensley. pp. 167-168. ISBN 0-905418-96-4.
- «National symbols of Estonia» (en inglés). The Estonia Institute. Archivado desde el original el 14 de enero de 2012.
Enlaces externos
 Wikimedia Commons alberga una categoría multimedia sobre Hirundo rustica.
Wikimedia Commons alberga una categoría multimedia sobre Hirundo rustica. Wikispecies tiene un artículo sobre Hirundo rustica.
Wikispecies tiene un artículo sobre Hirundo rustica.
- Fauna Ibérica: la golondrina común. Información sobre especies animales de la península ibérica.
- Identificación de sexos y edades en el Atlas de identificación de las aves de Aragón.
| Control de autoridades |
|
|---|
 Datos: Q25429
Datos: Q25429- Multimedia: Hirundo rustica / Q25429
 Citas célebres: Golondrina
Citas célebres: Golondrina- Especies: Hirundo rustica