Jianianhualong tengi
Jianianhualong tengi es la única especie conocida del género extinto Jianianhualong de dinosaurio terópodo trodóntido, que vivió a comienzos del período Cretácico, hace aproximadamente entre 125 millones de años, durante el Aptiense, en lo que hoy es Asia. Nombrado en 2017 por Xu Xing y sus colegas basada en un esqueleto articulado que preserva las plumas. Las plumas en el centro de la cola de Jianianhualong son asimétricas, siendo el primer registro de plumas asimétricas entre los trodóntidos. A pesar de las diferencias aerodinámicas de las plumas de vuelo de las aves modernas, las plumas en la paleta de la cola de Jianianhualong podrían haber funcionado en reducción de la fricción mientras el animal se movía. El descubrimiento de Jianianhualong apoya la noción de que las plumas asimétricas aparecieron al principio de la historia evolutiva de los paravianos.
| Jianianhualong | ||
|---|---|---|
| Rango temporal: 125 Ma Cretácico Inferior | ||
 Recreación | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Sauropsida | |
| (sin rango): | Maniraptora | |
| Superorden: | Dinosauria | |
| Orden: | Saurischia | |
| Suborden: | Theropoda | |
| (sin rango): | Paraves | |
| Familia: | †Troodontidae | |
| Género: |
Jianianhualong Xu et al., 2017 | |
| Especie: |
J. tengi Xu et al., 2017 | |
Jianianhualong posee una combinación de rasgos que se observan tanto los trodóntidos basales como en los derivados. Esto es consistente con su posición filogenética intermedia entre los trodóntidos. Esta mezcla de rasgos muestra una organización espacial distinta, con rasgos basales presentes en las extremidades anteriores y la pelvis, y rasgos derivados en el cráneo y las extremidades posteriores. Esto puede representar un caso de evolución en mosaico, donde la selección natural actúa sobre la forma del cuerpo de manera modular. Patrones de transición similares de los rasgos se observan en Sinusonasus, un pariente cercano de Jianianhualong. Ecológicamente hablando, Jianianhualong y Sinusonasus son parte de la considerable diversidad de trodóntidos que está presente dentro de la Formación Yixian .
Descripción
El espécimen tipo conservado de Jianianhualong mide aproximadamente 1 metro, como falta la parte posterior de la cola, el animal probablemente habría tenido alrededor de 1,12 metros de largo en vida. Escalado desde una longitud del hueso del muslo de 11,7 centímetros, Xu et al. Encontró que pesaba 2,4 kilogramos.[1][2] A pesar de su pequeño tamaño, el espécimen estaba completamente desarrollado, como lo indica la fusión de las suturas en las vértebras.[1][3][4]
Mandíbula superior
El cráneo de Jianianhualong es pequeño. La mandíbula tiene aproximadamente el 75% de la longitud del fémur. Es más o menos triangular cuando se ve de lado, con un hocico corto y un techo de cráneo ancho como su pariente Mei long.[5] El aspecto corto del hocico se ve algo exagerado por el hecho de que faltan los huesos delanteros del hocico, los premaxilares. El hueso que se conserva al frente en el fósil es el maxilar , cuyo cuerpo principal es más alto que largo. Esto distingue a Jianianhualong de todos los demás miembros de la Troodontidae,[6][7] junto con el margen superior del hueso formando un gran ángulo de 45° con el margen inferior, su pariente Saurornithoides relativo tiene un ángulo de 6°[8] y Sinovenator 36°.[9] Hay una cresta en la rama yugal que se extiende hacia atrás del hueso, a diferencia de Byronosaurus,[10] Sinovenator,[9] Sinusonasus,[11] Xixiasaurus,[12] y Zanabazar,[8] pero también está acompañado por un surco, que se ve en Byronosaurus.[13] La rama yugal relativamente profunda une Jianianhualong con los trodóntidos derivados pero no con Sinovenator.[1][6][7]
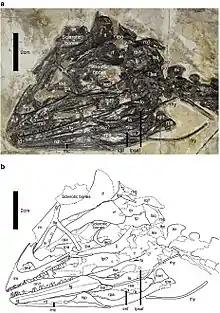
Detrás del maxilar, la depresión en la pared del cráneo conocida como fosa antorbital es grande y aproximadamente triangular,[6] y tiene dos aberturas grandes, la fenestra antorbital y la fenestra maxilar alargada que se extienden desde el margen superior hasta la parte inferior Margen de la depresión. En general, esta condición se asemeja a la de los trodóntidos derivados,[6][8][14] Sinornithoides , Sinovenator y Sinosunasus tienen una abertura adicional conocida como apertura promaxilar.[9][11][15] Sin embargo, el borde de la fosa es agudo como el de Byronosaurus,[13][16] y la barra de hueso que separa la fenestra es estrecha como en Sinovenator.[7] Bordeando la parte superior y posterior de la depresión está el lagrimal en forma de T, que tiene una rama larga que se proyecta hacia adelante y una pestaña en la unión entre las ramas que se proyectan hacia atrás y hacia abajo, al igual que con otros trodóntidos.[17] La rama descendente es casi igual a la longitud de la rama que se proyecta hacia adelante, más larga que en otros trodóntidos,[6] y la rama descendente también tiene una cresta única en su parte frontal.[1]
El hueso nasal es corto y ancho como el de Mei[5] y tiene una fila de foramenes, pequeñas aberturas para las arterias y los nervios, como en otros trodóntidos. También al igual que otros trodóntidos, el hueso frontal se curva hacia atrás suavemente en el margen exterior que bordea la cuenca del ojo.[6] En la parte inferior del hueso, hay una arista prominente conocida como crista cranii, que se aleja gradualmente del borde de la cavidad ocular hacia la línea media de la parte posterior del hueso. La punta frontal de la cresta hace contacto con el lagrimal, con una articulación relativamente simple a diferencia de la de Stenonychosaurus.[18] La superficie interior de la frontal sugiere detalles acerca de la caja craneal , incluyendo el ensanchado tracto olfativo y trasera de la cerebro. Al igual que otros trodóntidos, hay una cresta en la rama que proyecta hacia delante del yugal.[19] También al igual que otros trodóntidos, hay una depresión en el corto y profundo proceso de proyección hacia atrás, donde se superpone con el hueso cuadratojugal.[7] La rama subtriangular y curvada hacia atrás del hueso cuadrado , que se articula con el hueso pterigoideo , está más cerca de la parte inferior del hueso que en la mayoría de los demás terópodos no aviares. Comparado con Sinovenator, la rama yugal larga del hueso postorbital no es tan larga..[1][7]
Mandíbula inferior
En general, la mandíbula inferior es más o menos triangular cuando se ve desde el lado, con un margen superior recto y un margen inferior curvo, y no se recorta a diferencia de Sinovenator.[7] El dentario, el hueso de la mandíbula inferior frontal que tiene dientes, es largo, ocupa la mitad de la longitud de la mandíbula, poco profundo y triangular, y tiene un surco que se ensancha hacia la parte posterior del hueso, como en otros trodóntidos.[6] También, por lo general, los trodóntidos son los muchos agujeros hacia la parte frontal del dentario, que están particularmente concentrados dentro del surco.[13] Más atrás, hay un canal ubicado cerca de la parte posterior de la superficie superior del hueso surangular, que no se ve en otros trodóntidos. Los bordes surangulares de la fenestra mandibular, la abertura en la pared externa posterior de la mandíbula, desde arriba y desde atrás. Debajo de la fenestra, el hueso angular divide al dentario y la fenestra mandibular por una rama robusta que se proyecta hacia delante y que se curva fuertemente hacia arriba. Esto se ve en Sinovenator, otros trodóntidos, y también algunos dromeosáuridos[7] puede ser una característica definitoria de los trodóntidos o deinonicosaurianos. En el interior de la mandíbula, el hueso esplenial sub-triangular no tiene una muesca en forma de V en la parte posterior, a diferencia de la de Deinonychus y otros terópodos.[20] Detrás de lo esplenial, el hueso prearticular se curva hacia la parte inferior de la mandíbula, con una hoja delgada en la parte delantera y un eje redondeado en la parte posterior. Esto es similar a otros maniraptoranos , el grupo más grande al que pertenecen los trodóntidos.[1]
Dientes
Los maxilares de Jianianhualong, sus huesos principales de la mandíbula superior, tienen 21 dientes en cada lado, mientras que los dentarios tienen 25 en cada lado. El número en los premaxilares es desconocido, aunque es probable que hayan sido cuatro por lado. Al igual que los de otros trodóntidos, los dientes están fuertemente recurvados y distribuidos de manera desigual.[6] Los dientes en la parte frontal de las mandíbulas están muy apretados, mientras que son más pequeños y más espaciados en la parte posterior de las mandíbulas. Característica de los trodóntidos, los dientes tienen una corona corta, con relaciones de altura a anchura que disminuyen de 1.5 en el primer diente a 1.2 en el sexto diente a lo largo de la mandíbula. Los dientes cerca de la parte frontal de las mandíbulas carecen de denticulos, mientras que los dientes más atrás tienen denticulos en sus bordes posteriores. Estas estrías son finas, como en Sinovenator,[7] en lugar de robustas como en los trodóntidos derivados.[6] Al igual que los trodóntidos contemporáneos, el último diente maxilar se ubica cerca del extremo posterior de la rama yugal del maxilar, que está mucho más atrás que la mayoría de los celurosaurianos. Inusualmente, a diferencia de los dientes maxilares, varios dientes frontales en el dentario parecen estar inclinados hacia delante, son "procumbentes". Esta condición está presente en varios dromeosáuridos contemporáneos de Jianianhualong, a pesar de que no son parientes cercanos.[1][7]
Esqueleto axial

La columna vertebral del fósil de Jianianhualong está casi completa. Sin embargo, las vértebras individuales no siempre son fáciles de discernir. El cuello, es decir, las vértebras cervicales tiene 16 centímetros de largo, el torso, es decir, las vértebras dorsales, tiene 17 centímetros de longitud y la cola, es decir, las vértebras caudales, tiene 54 centímetros largo. En general, la cola habría sido 3,9 veces la longitud del fémur; Puede haber habido 31 vértebras caudales en total.[1]
Cuello
De manera única entre los trodóntidos, la espina neural del axis, la segunda vértebra del cuello, tiene un margen superior convexo y la parte posterior está muy expandida. La tercera vértebra del cuello tiene una longitud moderada del centro, cuerpo vertebral, de 12 milímetros Y tiene una espina neural corta. Asociada pero separada de esta cervical está la tercera costilla del cuello, que es larga y delgada, y se superpone a la tercera y cuarta vértebras del cuello. La cuartoa vértebra del cuello es un poco más larga con una costilla más robusta, y tiene una espina neural rectangular 1,5 veces más larga que alta. La quinta a la séptima vértebras tienen la mayor longitud, con la sexta y la séptima con cojinete neumático, invadido por sacos de aire, un sistemas de foramina o aberturas, fosas o depresiones y láminas o crestas en sus lados. Sus costillas correspondientes son ligeramente más largas que sus cuerpos vertebrales. Más atrás, las vértebras del cuello restantes son significativamente más cortas y tienen costillas con cabezas expandidas de igual longitud a sus cuerpos vertebrales.[1]
Torso
Las vértebras delanteras del torso son relativamente cortas, con las vértebras medias y traseras relativamente más largas. No hay aberturas ni depresiones neumáticas, pleurocoelos, en los lados de las vértebras delanteras, las vértebras medias y posteriores probablemente fueron similares a este respecto. Los cuerpos de las vértebras medias y posteriores son bajos, con una relación longitud-altura de 2,0, al igual que otros pequeños deinonicosaurianos.[7] Al igual que otros miembros basales, menos especializados, de los paravianos, las espinas neurales de la espalda tienen forma de abanico, y las espinas neurales pertenecen a las vértebras traseras que se adhieren a las mitades traseras de los cuerpos vertebrales correspondientes.[7] Ninguna de las costillas del torso tiene un proceso uncinato parecido a un ave, un gancho para la unión de los músculos que ayudan a la respiración, y el esternón tampoco está osificados. Esto es similar a otros trodóntidos.[6] Al igual que Velociraptor y otros terópodos, el primer par de gastralia, costillas del abdomen, cerca de la parte frontal del torso es notablemente más robusto, particularmente en los segmentos de la porción central.[1][21]
Cola
Similar a otros paravianos, las vértebras de Jianianhualong aumentan a lo largo de la cola, por ejemplo, la novena vértebra de la cola es 1,7 veces la longitud de la octava, mientras que la 15 vértebra, entre la más larga, con una longitud de 23,3 milímetros, es aproximadamente el doble que la sexta. Desde la 23 vértebra hacia atrás, los lados se comprimen de tal manera que se vuelven sub-triangulares en lugar de rectangulares en sección transversal; esta transición se produce en dos tercios de la longitud de la cola, como en Sinovenator. Desde la 24 vértebra hacia atrás, la superficie articular delantera de los cuerpos vertebrales se vuelve más profunda que la superficie articular trasera. No hay evidencia de un surco en la parte inferior de los cuerpos de la cola delantera, aunque la mayoría de los celurosaurios poseen uno. Al igual que con otros trodóntidos, los procesos articulares o cigapófisis de las vértebras de la cola son pequeños y casi horizontales, en lugar de ser tan largos como en otros tetanuros.[1][6]
Hay espinas neurales en las primeras ocho vértebras de la cola; se desconoce si las espinas neurales se reemplazan por un surco más hacia atrás en la serie, como sucede en otros trodóntidos.[6] En la parte inferior de la cola, los cheurones Los huesos separados que sobresalen hacia abajo son como placas. Los cheurones que se articulan con la primera a quinta vértebra son mucho más altos que largos, los que articulan con la sexta a novena vértebra son más largos que altos, aunque todavía son relativamente altos y aquellos que se articulan con la novena, décima y otras vértebras son muy superficiales y algo aplanados. En las vértebras hacia la parte posterior de la cola, las proyecciones hacia delante y hacia atrás de los cheurones son aproximadamente iguales en longitud. En estos cheurones, la proyección hacia adelante es contundente, mientras que la proyección hacia atrás es algo bifurcada o bifurcada. A diferencia de Sinusonasus, donde todos los cheurones entran en contacto unos con otros,[11] hay algunas brechas entre los cheurones de Jianianhualong.[1]
Cintura escapular y brazo
La escápula, la parte superior de la cintura escapular, que tiene una cuchilla en forma de correa y tiene una longitud aproximadamente igual al húmero, se conserva en un ángulo de 40 ° con respecto al eje largo de las vértebras del tronco. Esto es más similar a los especímenes articulados de terópodos más basales, a diferencia de los paravianos, en los que la escápula es casi paralela a las vértebras.[7] Al igual que otros miembros de la Pennaraptora, posee un coracoides grande de cuatro lados,[22] la parte inferior de la cintura escapular, es aproximadamente 1,3 veces más largo que ancho.[23] La parte inferior del coracoides se extiende hacia atrás con una proyección en forma de gancho, el proceso posglenoideo, que es similar en longitud al de Sinornithoides pero más largo que el de Sinovenator.[7] A diferencia de Sinornithoides,[24] la furcula es robusta, aplanada y en forma de U como la de Mei.[1]
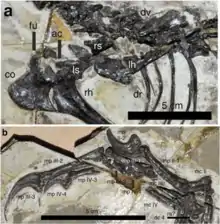
Al igual que los trodóntidos derivados, el hueso de la parte superior del brazo es aproximadamente el 70% de la longitud del hueso del muslo. Sin embargo, en la parte superior del hueso del brazo, la cresta deltopectoral, la placa ósea que sirve para la inserción muscular, es corta en comparación con Linhevenator y otros trodóntidos derivados,[19] que ocupan solo el 20% de la longitud del eje. El cúbito, el hueso principal de la parte inferior del brazo, que es el 88% de la longitud de la parte superior del brazo, presenta un ligero arco hacia atrás. Mei muestra un arco mucho más fuerte,[5] mientras que está ausente en la mayoría de los otros trodóntidos.[22] El olecranon del cúbito, el proceso para la unión de los músculos que estiran el brazo, se desarrolla levemente y parece estar acompañado por un reborde prominente más abajo en el hueso. En comparación con el cúbito, el radio, el hueso del brazo inferior paralelo, es ligeramente más delgado, con un diámetro del eje que es del 76% del cúbito. Contrariamente a la mayoría de los otros terópodos, la parte superior del radio lleva un arco hacia atrás similar a la del cúbito.[1]
La mano tiene tres dedos, típicos de la Maniraptora, que respectivamente llevan dos, tres y cuatro falanges. Los autores consideran que los tres dígitos representan el segundo, el tercero y el cuarto, mientras que la mayoría de los otros investigadores los ven como el primero, el segundo y el tercero.[25] La longitud total de la mano es de 112 milímetros, 140% de la parte superior del brazo y 95% del hueso del muslo. En la muñeca, el carpo semilunar, un hueso plano y curvado, es relativamente pequeño; La superficie superior es fuertemente convexa y tiene una ranura transversal única. Entre los tres huesos metacarpianos en la parte media, el final del metacarpiano IV, correspondiente al tercer dígito presente, se extiende ligeramente más allá del final del metacarpiano III, que corresponde al segundo dígito presente, que de otro modo no se ve fuera de la Avialae . Sin embargo, las aves enantiornithes como Patagopteryx también poseen este rasgo.[26] El extremo superior del metacarpiano IV también está situado más hacia abajo que el del metacarpiano III, que es el caso de muchos paravianos.[1]
Varias características distintivas de Jianianhualong se encuentran más abajo entre las falanges. La falange II-1, la primera falange del primer dígito o dedo, es inusualmente larga, siendo más corta que el metacarpiano III, lleva un "talón" en su extremo superior, así como una gran ranura en la superficie interior que se extiende desde la parte superior del hueso por más de la mitad de su longitud. Además, la falange III-2 también es muy larga, es más larga que el metacarpiano III, un rasgo que solo se ve en los ornitomimosaurianos basales , así como en los paravianos Xiaotingia y Yixianosaurus.[23] Las garras o falanges unguales son muy robustas, con un ancho basal de al menos el 50% de su longitud total. La garra del primer dedo es la más grande. Al igual que con los dromeosáuridos, la pequeña garra del tercer dedo tiene un leve arco hacia arriba, pero todas las garras en las manos de los dromeosáuridos están arqueadas.[1][27]
Cintura pelvica y patas

La pelvis de Jianianhualong es en general similar a la de Sinovenator. Al igual que en Archaeopteryx y los dromeosáuridos basales, el ilion , el hueso superior de la pelvis, es pequeño, con un 60% de la longitud del fémur.[28] La mayoría de los demás terópodos tienen un ilion que tiene al menos el 70% de la longitud del fémur. También inusualmente entre los terópodos, el margen superior del ilion es cóncavo. La "pierna" que se proyecta hacia adelante del ilion, el pedúnculo púbico, tiene una depresión grande y orientada hacia delante en su lado, que sirve como un accesorio para el músculo protractor del fémur, que mueve el hueso del muslo hacia adelante, como "musculus cuppedicus".[29] Debajo del ilion, los huesos púbicos izquierdo y derecho se unen en el "delantal púbico" inusualmente amplio, una lámina ósea de conexión, que es característica de los trodóntidos,[30] con una cresta en la superficie posterior como en Sinovenator.[1]
Detrás del hueso púbico, el isquion es una placa ósea corta que tiene el 48% de la longitud del hueso púbico. Al igual que en los dromeosáuridos basales y las aves basales, hay una proyección dirigida hacia arriba cerca de la parte superior del borde posterior del isquion, donde se articula con el ilion.[28][32] Más abajo, el borde del hueso se curva hacia adentro, creando un perfil hueco, seguido de una segunda proyección hacia arriba, una combinación que también se ve en Archaeopteryx. El proceso del obturador que se proyecta hacia adelante en el borde frontal del isquion se encuentra cerca de la parte inferior del hueso como en los dromeosáuridos basales y las aves basales, pero es pequeño como el de Archaeopteryx. Hay un pequeño receso en el margen del proceso, Buitreraptor y Sinornithosaurus tienen un receso similar pero comparativamente más grande.[1]

Como con la mayoría de los terópodos, el fémur se curva hacia delante.[33] El extremo inferior del fémur es más robusto que el extremo superior, un rasgo compartido con Anchiornis.[34] Entre los cuatro huesos metatarsianos del pie medio, el metatarsiano II es el más corto y más delgado, con un 88% de la longitud del metatarsiano III, además del metatarsiano I, correspondiente al hallux o dígito I. Metatarsal III tiene el 72% de la longitud del hueso del muslo, que es más corto que el tercer metatarsiano de Sinovenator que tiene el 90% de la longitud del hueso del muslo. A diferencia de otros trodóntidos,[33] el extremo inferior del metatarsiano II no tiene una articulación articulada pero está redondeado, y también falta una pestaña en el fondo del metatarsiano IV, su ausencia es una característica distintiva de Jianianhualong. Sin embargo, el metatarsiano IV es en general robusto como en otros trodóntidos.[33] En general, las falanges del hallux son largas, con un 31% de la longitud del metatarsiano III; casi exactamente la misma proporción está presente en Stenonychosaurus.[1]
En el segundo dedo del pie, dígito II, la falange II-1, el primero del pie, tiene un extremo superior alto, y es aproximadamente 1,5 veces la longitud de la falange II-2, el segundo. Como en otros deinonicosaurianos derivados, la falange II-2 tiene un prominente "talón" en su extremo superior, que se extiende significativamente.[21][20][35][36] El extremo inferior de la falange II-2 tiene extensiones por debajo y por encima del diámetro del eje, un rasgo que también se ve en otros trodóntidos y dromeosáuridos basales que habrían mejorado el arco rotacional del articulación.[37] Al igual que los trodóntidos y dromeosáuridos derivados, la "garra de la hoz", el ungual del dígito II, está fuertemente curvada y tiene un tubérculo flexor prominente, una elevación que sirve como accesorio para el tendón que dobla la garra.[19][35] también es mucho más grande que las otras falanges en el segundo dedo del pie, siendo 1.4 veces la longitud de la falange II-1 mientras que tienen la misma longitud en el pie de Sinovenator.[1]
Plumas
Las plumas se conservan a lo largo de toda la columna vertebral de Jianianhualong. Sin embargo, solo aquellos a lo largo de la cola se muestran con mucho detalle. En la parte inferior del cuello, las plumas se conservan con una longitud de al menos 30 milímetros. Las que están a lo largo de la espalda y sobre la cadera son más largas, con una longitud de aproximadamente 75 milímetros. Detrás de la tibia o la espinilla, también hay algunas plumas que miden 70 milímetros de longitud. Además, algunas plumas mal conservadas están asociadas con el brazo, tanto el húmero como el cúbito, que tienen una longitud desconocida.[1]
Al igual que la de Jinfengopteryx , Anchornis y Archaeopteryx , la cola de Jianianhualong sostiene un abanico de plumas penaceas similar a una fronda en toda su longitud. A diferencia de Archaeopteryx y potencialmente Jinfengopteryx ,[38][39] las plumas son curvas, con al final de la cola un borde frontal convexo y un borde posterior cóncavo; en la base de la cola es al revés. Las plumas del centro de la cola son de 120 milímetros de largo y al menos 10 milímetros de ancho. Como en Archeopteryx al menos algunas de las largas plumas penaceas que recubren la cola son asimétricas, una pluma en el centro de la cola tiene una paleta de arrastre que es aproximadamente dos veces más ancha que la paleta delantera, con los filamentos respectivamente en ángulos de 10° y 15° para las dos paletas. En Archaeopteryx, las plumas al final de la cola, que no se conservan en Jianianhualong, son simétricas y tienen puntas redondeadas.[1][39]
Descubrimiento e investigación
Jianianhualong es conocido por el holotipo y única muestra, un esqueleto casi completo que conserva las plumas y le falta solo la punta de la cola. Está articulado y comprimido sobre una placa de piedra. Este espécimen holotipo se almacena bajo el número de colección DLXH 1218 en el Museo Dalian Xinghai, provincia de Liaoning, China. Se excavó con herramientas mecánicas, martillos y cinceles de geólogo, de las rocas del Cretácico temprano[40] de la formación Yixian en la localidad de Baicai Gou, en chino, 白菜 白菜, ubicada en el Condado de Yixian de Liaoning.[1] La cama Dakangpu, equivalente a las camas Dawangzhangzi, Daxinfangzi, Hejiaxin, Sichakou y Senjiru,[41] de la cual Baicai Gou forma parte,[42] está situada en el centro de la formación. La parte específica de las Camas de las que se origina Jianianhualong tenía una fecha en 2006 y 2008 de aproximadamente 124,4 millones de años, lo que corresponde al Aptiense temprano.[43][44][45]
En 2017, Jianianhualong fue nombrado y descrito por un equipo de investigadores que incluye a Xu Xing , Philip J. Currie , Michael Pittman, Xing Lida, Meng Qingjin, Lü Junchang , Hu Dongyu y Yu Congyu, en un trabajo de investigación publicado en Nature Communications . Para investigar el plumaje preservado de la muestra, Xu y sus colegas utilizaron la fluorescencia estimulada por láser utilizando un láser de 405 nanometros.[46] El nombre del género combina el nombre de una compañía que patrocinó la investigación de este taxón, Jianianhua, 嘉年華 , y el sufijo long, 龍, que significa "dragón". El nombre específico, tengi , honra a la Sra. Teng Fangfang que adquirió el espécimen y lo puso a disposición para su estudio.[1]
Clasificación
Una serie de características permiten identificar a Jianianhualong como miembro de los Troodontidae. Estos incluyen la rama y la brida proyectadas hacia delante largas del arco lagrimal, la fuerza sobre el hueso nasal, la transición suave entre la cavidad del ojo y la rama que se proyecta hacia atrás del hueso frontal, la cresta en la rama que proyecta hacia delante del hueso yugal, el dentario triangular que lleva un surco ensanchado. La robusta rama proyectada hacia delante del hueso surangular, el número relativamente grande de dientes desigualmente distribuidos, los cheurones aplanados con proyecciones romas hacia adelante y proyecciones bifurcadas hacia atrás; y el ancho y plano "delantal púbico" formado por los huesos púbicos.[1][6]
Dentro de los trodóntidos, Jianianhualong muestra una combinación única de características de los trodóntidos basales y derivados, organizados en distintas regiones de su cuerpo. Las extremidades anteriores y la pelvis se parecen en gran medida a los trodóntidos basales, como Sinovenator, la cresta deltopectoral corta en el húmero, la mano larga, el metacarpiano II y la falange III-2 en la mano, el pequeño ilion, la cresta en el delantal púbico, y el isquion corto con dos proyecciones y un proceso de obturador cerca de la parte inferior del hueso.[6][7] Al mismo tiempo, el cráneo y las extremidades posteriores se asemejan a los trodóntidos derivados, la rama yugal profunda del maxilar que tiene un surco prominente, la fosa antorbitaria sub-triangular, la fenestra maxilar relativamente larga, la rama yugal corta del hueso postorbital, el largo hallux el metatarsiano delgado y corto II; el "talón" en la falange II-2 y la gran garra curvada.[6] Sinosonasus muestra una condición similar: el cráneo es como el de los trodoótidos basales, y la pelvis y la extremidad posterior son como los de los trodoótidos derivados.[1]
La conjunción de características basales y derivadas en distintas regiones del cuerpo en Jianianhualong y Sinosonasus representan un posible caso de evolución en mosaico, en el que la selección evolutiva actúa sobre distintos "módulos" del cuerpo.[47] De hecho, el patrón en el que están presentes las características basales y derivadas parece seguir los dominios del cuerpo que están regulados por los genes Hox, que se han discutido previamente en el contexto de pterosaurios como Darwinopterus.[48] Sin embargo, esta hipótesis se ve debilitada por la presencia de características en ambos trondóntidos que no siguen el patrón. En Jianianhualong, por ejemplo, la fenestra antorbital grande, la fila de dientes largos del maxilar y los dientes finamente serrados representan características basales en un cráneo que de otra manera se deriva. De manera similar, el dígito II del pie en Sinusonasus no es tan especializado como el de los trondóntidos derivados, a pesar de que el miembro posterior se deriva en general. Por el contrario, la extremidad anterior de Jianianhualong es corta en general como en los trondóntidos derivados, a pesar de la presencia de rasgos basales.[1]
Una reconstrucción del estado ancestral realizada junto con el análisis filogenético sugiere que la asimetría de las plumas del ala evolucionó solo una vez, en el último ancestro común de las paraves. Las plumas asimétricas de la cola también habrían evolucionado una vez, en el antepasado común de todos los paravianos, excluyendo los escansoriopterígidos y avialanos. Sin embargo, dada la incertidumbre de si los dromeosáuridos poseían plumas de alas asimétricas, el apoyo para una aparición única de plumas de alas asimétricas en el antepasado común de los Paraves es equívoco, ya que el escenario alternativo que evolucionó dos veces es plausible. Así queda la clara posibilidad de que los trodóntidos tuvieran plumas de cola asimétricas pero plumas de brazos simétricas, lo que sugiere que la asimetría en la cola apareció primero, pero esto no está respaldado por la reconstrucción del estado ancestral.[1]
Filogenia
Filogenéticamente , Xu y sus colegas encontraron a Jianianhualong en una politomía con Sinusonasus y varios otros trodoótidos intermedios.[1]

| Paraves |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Paleobiología
Jianianhualong conserva la primera evidencia de plumas en un incontestable trodóntido, y las afinidades de los taxones, como Anchiornis, Eosinopteryx, Jinfengopteryx y Xiaotingia, han sido cuestionadas por varios estudios.[38][49][50][51] Además, Jianianhualong representa el primer registro de plumas asimétricas entre los trodóntidos, haciendo de estos el segundo grupo entre los terópodos no aves que poseen plumas asimétricas. Las plumas asimétricas del brazo se encuentran en los microraptorinos Microraptor, Changyuraptor y posiblemente Zhenyuanlong.[52][53] Las plumas asimétricas se correlacionan con las capacidades de vuelo entre los paravianos,[54] y su aparición marcó un paso importante en la evolución de las plumas aviares modernas.[55][56][57] Este rasgo puede haber sido seleccionado debido a su aumento de la locomoción en el suelo o en el aire.[1][58]
Sin embargo, la presencia de plumas asimétricas en Jianianhualong no garantiza que sea capaz de volar de volar.[59] Las plumas en la cola de Jianianhualong ciertamente podrían haber funcionado para mantener la estabilidad de las plumas, las barbas del borde delantero, con un ángulo bajo de 10 °, aumentaron la rigidez de la paleta[60] o controlando el flujo de aire, a través de la variación de la inclinación de la hoja de las plumas.[61][62] Sin embargo, estas plumas difieren de las de las aves modernas en que las barbas del borde posterior también tienen un ángulo relativamente bajo de aproximadamente 15 °. Así, aerodinámicamente hablando, las plumas en la cola de Jianianhualong probablemente no tenía la misma función que las de las aves modernas.[59] En cualquier caso, el contorno subtriangular de la hoja de la cola, así como las ranuras que separan las puntas de las plumas individuales, habrían proporcionado una reducción de la resistencia cuando la cola estaba en uso.[1][63][64]
Paleoecología
Junto con Mei, Sinovenator, Sinusonasus y Jinfengopteryx, Jianianhualong es el quinto trodóntido nombrado que se descubrió en la Biota de Jehol, con los otros tres primeros trodóntido que también se originaron en la formación Yixian. Con Jianianhualong y Sinusonasus que representan formas "transicionales" y más derivadas de trodóntidos en comparación con Mei y Sinovenator, la diversidad de trodóntidos dentro del Jehol fue apreciable.[1] Además de los trodóntidos, la formación Yixian también conserva un conjunto diverso de otros animales, tanto dinosaurios como no dinosaurios. El hadrosauroide Jinzhousaurus se conoce en Baicai Gou, la misma localidad que la de Jianianhualong, así como en el maniraptor Yixianosaurus, así como en cientos de especímenes del coristodero Hyphalosaurus .
Desde otras partes del lecho Dakangpu/Dawangzhangzi, se han descubierto ejemplares pertenecientes a las aves Confuciusornis, Hongshanornis, Grabauornis, Liaoxiornis, Longicrusavis , Shanweiniao, Shenqiornis y Zhongornis, el terópodo no aviar Sinosauropteryx, el ceratopsiano Psittacosaurus, el pterosaurio Cathayopterus, los lagartos Dalinghosaurus y Yabeinosaurus. Los coristodero Hyphalosaurus y Monjurosuchus, de los cuales el primero es muy abundante, la tortuga Manchurochelys. los mamíferos Acristatherium, Akidolestes , Chaoyangodens, Eomaia, Sinobaatar y Sinodelphys, la rana Liaobatrachus y peces, incluyendo Lycoptera, de los cuales L. davidi es numeroso, Peipiaosteus y Protopsephurus.
Ambientalmente, Jianianhualong vivía en un área lacustre, como lo demuestra la arensica presente en todas la cama Dawangzhangzi. Al igual que el resto de la formación Yixian, el ambiente representado por estos ensamblajes tenían un clima semiárido estacional, con una temperatura del aire media baja de 10 ± 4 °C. ELa flora local consiste en un conjunto mixto de gimnospermas, tales como Czekanowskia, Schizolepis y los efedráceas Amphiephedra, Chengia, Ephedrites y Liaoxia y también angiospermas tempranas, incluyendo Archaefructus , Hyrcantha, anteriormente Sinocarpus , Leefructus y Potamogeton. Leefructus en particular es una eudicota, estando entre los registros más antiguos del grupo.
Referencias
- Xu, X.; Currie, P.; Pittman, M.; Xing, L.; Meng, Q.; Lü, J.; Hu, D.; Yu, C. (2017). «Mosaic evolution in an asymmetrically feathered troodontid dinosaur with transitional features». Nature Communications 8: 14972. Bibcode:2017NatCo...814972X. PMC 5418581. PMID 28463233. doi:10.1038/ncomms14972.
- Campione, N.E.; Evans, D.C.; Brown, C.M.; Carrano, M.T. (2014). «Body mass estimation in non-avian bipeds using a theoretical conversion to quadruped stylopodial proportions». Methods in Ecology and Evolution 5 (9): 913-923. doi:10.1111/2041-210X.12226.
- Brochu, C.A. (1996). «Closure of neurocentral sutures during crocodilian ontogeny: Implications for maturity assessment in fossil archosaurs». Journal of Vertebrate Paleontology 16 (1): 49-62. doi:10.1080/02724634.1996.10011283.
- Irmis, R.B. (2007). «Axial skeleton ontogeny in the Parasuchia (Archosauria: Pseudosuchia) and its implications for ontogenetic determination in archosaurs». Journal of Vertebrate Paleontology 27 (2): 350-361. doi:10.1671/0272-4634(2007)27[350:ASOITP]2.0.CO;2.
- Xu, X.; Norell, M.A. (2004). «A new troodontid dinosaur from China with avian-like sleeping posture». Nature 431 (7010): 838-841. Bibcode:2004Natur.431..838X. PMID 15483610. doi:10.1038/nature02898.
- Makovicky, P.J.; Norell, M.A. (2004). «Troodontidae». En Weishampel, D.B.; Dodson, P.; Osmólska, H., eds. The Dinosauria (2nd edición). Berkeley: University of California Press. pp. 184-195.
- Xu, X. (2002). Deinonychosaurian Fossils From the Jehol Group of Western Liaoning and the Coelurosaurian Evolution (Tesis de Ph.D.). Beijing: Graduate School of the Chinese Academy of Sciences. pp. 1-322.
- Norell, M.A.; Makovicky, P.J.; Bever, G.S.; Balanoff, A.M.; Clark, J.M.; Barsbold, R.; Rowe, Timothy (2009). «A Review of the Mongolian Cretaceous Dinosaur Saurornithoides (Troodontidae: Theropoda)». American Museum Novitates 3654: 1-63. doi:10.1206/648.1. hdl:2246/5973.
- Xu, X.; Norell, M.A.; W., X.-L.; Makovicky, P.J.; Wu, X.-C. (2002). «A basal troodontid from the Early Cretaceous of China». Nature 415 (6873): 780-784. Bibcode:2002Natur.415..780X. PMID 11845206. doi:10.1038/415780a.
- Bever, G.S.; Norell, M.A. (2009). «The perinate skull of Byronosaurus (Troodontidae) with observations on the cranial ontogeny of paravian theropods». American Museum Novitates 3657: 1-51. doi:10.1206/650.1. hdl:2246/5980.
- Xu, X.; Wang, X.-L. (2004). «A New Troodontid (Theropoda: Troodontidae) from the Lower Cretaceous Yixian Formation of Western Liaoning, China». Acta Geologica Sinica 78 (1): 22-26. doi:10.1111/j.1755-6724.2004.tb00671.x.
- Lü, J.; Xu, L.; Liu, Y.; Zhang, X.; Jia, S.; Ji, Q. (2010). «A new troodontid (Theropoda: Troodontidae) from the Late Cretaceous of central China, and the radiation of Asian troodontids». Acta Palaeontologica Polonica 55 (3): 381-388. doi:10.4202/app.2009.0047.
- Makovicky, P.J.; Norell, M.A.; Clark, J.M.; Rowe, T.E. (2003). «Osteology and relationships of Byronosaurus jaffei (Theropoda: Troodontidae)». American Museum Novitates 3402: 1-32. doi:10.1206/0003-0082(2003)402<0001:oarobj>2.0.co;2. hdl:2246/2828.
- Senter, P.; Kirkland, J.I.; Bird, J.; Bartlett, J.A. (2010). «A New Troodontid Theropod Dinosaur from the Lower Cretaceous of Utah». PLoS ONE 5 (12): e14329. Bibcode:2010PLoSO...514329S. PMC 3002269. PMID 21179513. doi:10.1371/journal.pone.0014329.
- Russell, D.; Dong, Z. (1993). «A nearly complete skeleton of a new troodontid dinosaur from the Early Cretaceous of the Ordos Basin, Inner Mongolia, People's Republic of China». Canadian Journal of Earth Sciences 30 (10): 2163-2173. Bibcode:1993CaJES..30.2163R. doi:10.1139/e93-187.
- Norell, M.A.; Makovicky, P.J.; Clark, J.M. (2000). «A new troodontid theropod from Ukhaa Tolgod, Mongolia». Journal of Vertebrate Paleontology 20 (1): 7-11. doi:10.1671/0272-4634(2000)020[0007:ANTTFU]2.0.CO;2.
- Xu, X.; Pittman, M.; Sullivan, C.; Choiniere, J.N.; Tan, Q.W.; Clark, J.M.; Norell, M.A.; Shuo, W. (2015). «The taxonomic status of the Late Cretaceous dromaeosaurid Linheraptor exquisitus and its implications for dromaeosaurid systematics». Vertebrata PalAsiatica 53 (1): 29-62.
- Currie, P. J. (1985). «Cranial anatomy of Stenonychosaurus inequalis (Saurischia, Theropoda) and its bearing on the origin of birds». Canadian Journal of Earth Sciences 22 (11): 1643-1658. Bibcode:1985CaJES..22.1643C. doi:10.1139/e85-173.
- Xu, X.; Tan, Q.; Sullivan, C.; Han, F.; Xiao, D. (2011). «A Short-Armed Troodontid Dinosaur from the Upper Cretaceous of Inner Mongolia and Its Implications for Troodontid Evolution». PLoS ONE 6 (9): e22916. Bibcode:2011PLoSO...622916X. PMC 3168428. PMID 21915256. doi:10.1371/journal.pone.0022916.
- Ostrom, J.H. (1969). «Osteology of Deinonychus antirrhopus, an unusual theropod from the Lower Cretaceous of Montana». Peabody Museum of Natural History Bulletin 30: 1-165.
- Norell, M.A.; Makovicky, P.J. (1997). «Important features of the dromaeosaur skeleton: information from a new specimen». American Museum Novitates 3215: 1-28. hdl:2246/3557.
- Gauthier, J.A. (1986). «Saurischian monophyly and the origin of birds». Memoirs of the California Academy of Sciences 8. pp. 1-55. ISBN 978-0-940228-14-6.
- Xu, X.; Sullivan, C.; Wang, S. (2013). «The systematic position of the enigmatic theropod dinosaur Yixianosaurus longimanus». Vertebrata PalAsiatica 51 (3): 169-183. Archivado desde el original el 6 de octubre de 2017. Consultado el 23 de julio de 2019.
- Nesbitt, S.J.; Turner, A.H.; Spaulding, M.; Conrad, J.L.; Norell, M.A. (2009). «The theropod furcula». Journal of Morphology 270 (7): 856-879. PMID 19206153. doi:10.1002/jmor.10724.
- Xu, X.; Clark, J.M.; Mo, J.; Choiniere, J.; Forster, C.A.; Erickson, G.M.; Hone, D.W.E.; Sullivan, C.; Eberth, D.A.; Nesbitt, S.; Zhao, Q.; Hernandez, R.; Jia, C.-K.; Han, F.-L.; Guo, Y. (2009). «A Jurassic ceratosaur from China helps clarify avian digital homologies». Nature 459 (18): 940-944. Bibcode:2009Natur.459..940X. PMID 19536256. doi:10.1038/nature08124.
- Chiappe, L. (1996). «Late Cretaceous birds of southern South America: anatomy and systematics of Enantiornithes and Patagopteryx deferrariisi». En Arratia, G., ed. Contributions of Southern South America to Vertebrate Paleontology. Munchner Geowissenschaftliche Abhandlungen, Reihe A, Geologie und Palaontologie 30. Munich: Verlag Dr. Friedrich Pfeil. pp. 203-244.
- Senter, P.; Barsbold, R.; Britt, B.B.; Burnham, D.B. (2004). «Systematics and evolution of Dromaeosauridae (Dinosauria, Theropoda)». Bulletin of the Gunma Museum of Natural History 8: 1-20.
- Xu, X.; Zhou, Z.; Wang, X. (2000). «The smallest known non-avian theropod dinosaur». Nature 408 (6813): 705-708. Bibcode:2000Natur.408..705X. PMID 11130069. doi:10.1038/35047056.
- Rowe, T. (1986). «Homology and evolution of the deep dorsal thigh musculature in birds and other Reptilia». Journal of Morphology 189 (3): 327-346. PMID 29940710. doi:10.1002/jmor.1051890310.
- Russell, D.A.; Dong, Z. (1993). «The affinities of a new theropod from the Alxa Desert, Inner Mongolia, People's Republic of China». Canadian Journal of Earth Sciences 30 (10): 2107-2127. Bibcode:1993CaJES..30.2107R. doi:10.1139/e93-183.
- Xu, X.; Wang, X.-L.; Wu, X.-C. (1999). «A dromaeosaurid dinosaur with a filamentous integument from the Yixian Formation of China». Nature 401 (6750): 262-266. Bibcode:1999Natur.401..262X. doi:10.1038/45769.
- Norell, M.A.; Makovicky, P.J. (1999). «Important features of the dromaeosaurid skeleton II: information from newly collected specimens of Velociraptor mongoliensis». American Museum Novitates 3282: 1-45. hdl:2246/3025.
- Xu, X.; Zhao, Q.; Norell, M.; Sullivan, C.; Hone, D.; Erickson, G.; Wang, X.; Han, F. et al. (2009). «A new feathered maniraptoran dinosaur fossil that fills a morphological gap in avian origin». Chinese Science Bulletin 54 (3): 430-435. doi:10.1007/s11434-009-0009-6.
- Currie, P.J.; Peng, J. (1994). «A juvenile specimen of Saurornithoides mongoliensis from the Upper Cretaceous of northern China». Canadian Journal of Earth Sciences 30 (10): 2224-2230. Bibcode:1993CaJES..30.2224C. doi:10.1139/e93-193.
- Xu, X.; Zhao, Ji; Sullivan, C.; Tan, Q.-W.; Sander, M.; Ma, Q.-Y. (2012). «The taxonomy of the troodontid IVPP V 10597 reconsidered». Vertebrata PalAsiatica 50 (2): 140-150.
- Xu, X.; Wang, X.-L. (2000). «Troodontid-like pes in the dromaeosaurid Sinornithosaurus». Paleontological Society of Korea, Special Publication 4: 179-188. Archivado desde el original el 18 de mayo de 2015. Consultado el 23 de julio de 2019.
- Foth, C.; Tischlinger, H.; Rauhut, O.W.M. (2014). «New specimen of Archaeopteryx provides insights into the evolution of pennaceous feathers». Nature 511 (7507): 79-82. Bibcode:2014Natur.511...79F. PMID 24990749. doi:10.1038/nature13467.
- Ji, S.; Ji, Q. (2007). «Jinfengopteryx compared to Archaeopteryx, with comments on the mosaic evolution of long-tailed avialan birds». Acta Geologica Sinica 81 (3): 337-343. doi:10.1111/j.1755-6724.2007.tb00957.x.
- Swisher, C.C.; Wang, X.; Zhou, Z.; Wang, Y.; Jin, F.; Zhang, J.; Xu, X.; Zhang, J. et al. (2002). «Further support for a Cretaceous age for the feathered-dinosaur beds of Liaoning, China: New 40Ar/39Ar dating of the Yixian and Tuchengzi Formations». Chinese Science Bulletin 47 (2): 136-139. doi:10.1360/02tb9031 (inactivo 2019-03-07). Parámetro desconocido
|doi-broken-date=ignorado (ayuda); - Wang, X.-L.; Zhou, Z.-H. (2008). «Mesozoic Pompeii». En Chang, M.-M.; Chen, P.-J.; Wang, Y.-Q.; Wang, Y.; Miao, D.-S., eds. The Jehol Fossils: The Emergence of Feathered Dinosaurs, Beaked Birds and Flowering Plants (2nd edición). Amsterdam: Academic Press. pp. 19-38. ISBN 9780123741738.
- Wang, X.-L.; Xu, X. (2001). «A new iguanodontid (Jinzhousaurus yangi gen. et sp. nov.) from the Yixian Formation of western Liaoning, China». Chinese Science Bulletin 46 (19): 1669-1672. doi:10.1007/BF02900633.
- Sun, G.; Dilcher, D.L.; Wang, H.; Chen, Z. (2011). «A eudicot from the Early Cretaceous of China». Nature 471 (7340): 625-628. Bibcode:2011Natur.471..625S. PMID 21455178. doi:10.1038/nature09811.
- Liu, Y.; Liu, Y.; Ji, S.; Yang, Z. (2006). «U-Pb zircon age for the Daohugou Biota at Ningcheng of Inner Mongolia and comments on related issues». Chinese Science Bulletin 51 (21): 2634-2644. doi:10.1007/s11434-006-2165-2.
- Meng, F.X.; Gao, S.; Liu, X.M. (2008). «U-Pb zircon geochronology and geochemistry of volcanic rocks of the Yixian Formation in the Lingyuan area, western Liaoning, China». Geological Bulletin of China 27: 364-373.
- Kaye, T.G.; Falk, A.R.; Pittman, M.; Sereno, P.C.; Martin, L.D.; Burnham, D.A.; Gong, E.; Xu, X. et al. (2015). «Laser-Stimulated Fluorescence in Paleontology». PLoS ONE 10 (5): e0125923. PMC 4446324. PMID 26016843. doi:10.1371/journal.pone.0125923.
- Brandon, R.N. (1999). «The Units of Selection Revisited: The Modules of Selection». Biology and Philosophy 14 (2): 167-180. doi:10.1023/A:1006682200831.
- Lü, J.; Unwin, D.M.; Jin, X.; Liu, Y.; Ji, Q. (2010). «Evidence for modular evolution in a long-tailed pterosaur with a pterodactyloid skull». Proceedings of the Royal Society B 277 (1680): 383-389. PMC 2842655. PMID 19828548. doi:10.1098/rspb.2009.1603.
- Godefroit, P.; Cau, A.; Hu, D.-Y.; Escuillié, F.; Wu, W.; Dyke, G. (2013). «A Jurassic avialan dinosaur from China resolves the early phylogenetic history of birds». Nature 498 (7454): 359-362. Bibcode:2013Natur.498..359G. PMID 23719374. doi:10.1038/nature12168.
- Xu, X.; You, H.; Du, K.; Han, F. (2011). «An Archaeopteryx-like theropod from China and the origin of Avialae». Nature 475 (7357): 465-470. PMID 21796204. doi:10.1038/nature10288. Archivado desde el original el 20 de diciembre de 2016. Consultado el 25 de julio de 2019.
- Agnolín, F.L.; Novas, F.E. (2013). Avian Ancestors: A Review of the Phylogenetic Relationships of the Theropods Unenlagiidae, Microraptoria, Anchiornis and Scansoriopterygidae. SpringerBriefs in Earth System Sciences. Springer Netherlands. pp. 1-96. ISBN 978-94-007-5636-6. doi:10.1007/978-94-007-5637-3.
- Xu, X.; Zhou, Z.; Wang, X.; Kuang, X.; Zhang, F.; Du, X. (2003). «Four-winged dinosaurs from China». Nature 421 (6921): 335-340. Bibcode:2003Natur.421..335X. PMID 12540892. doi:10.1038/nature01342.
- Lü, J.; Brusatte, S.L. (2015). «A large, short-armed, winged dromaeosaurid (Dinosauria: Theropoda) from the Early Cretaceous of China and its implications for feather evolution». Scientific Reports 5: 11775. Bibcode:2015NatSR...511775L. PMC 4504142. PMID 26181289. doi:10.1038/srep11775.
- Feduccia, A.; Tordoff, H.B. (1979). «Feathers of Archaeopteryx: Asymmetric Vanes Indicate Aerodynamic Function». Science 203 (4384): 1021-1022. Bibcode:1979Sci...203.1021F. PMID 17811125. doi:10.1126/science.203.4384.1021.
- Xu, X.; Zhou, Z.; Dudley, R.; Mackem, S.; Chuong, C.-M.; Erickson, G.M.; Varricchio, D.J. (2014). «An integrative approach to understanding bird origins». Science 346 (6215): 1253293. PMID 25504729. doi:10.1126/science.1253293.
- Xu, X.; Guo, Y. (2009). «The origin and early evolution of feathers: insights from recent paleontological and neontological data». Vertebrata PalAsiatica 47 (4): 311-329.
- Prum, R.O (1999). «Development and Evolutionary Origin of Feathers». Journal of Experimental Zoology Part B: Molecular and Developmental Evolution 285 (4): 291-306. PMID 10578107. doi:10.1002/(SICI)1097-010X(19991215)285:4<291::AID-JEZ1>3.0.CO;2-9.
- Dececchi, T.A.; Larsson, H.C.E.; Habib, M.B. (2016). «The wings before the bird: an evaluation of flapping-based locomotory hypotheses in bird antecedents». PeerJ 4: e2159. PMC 4941780. PMID 27441115. doi:10.7717/peerj.2159.
- Feo, T.J.; Field, D.J.; Prum, R.O. (22 de marzo de 2015). «Barb geometry of asymmetrical feathers reveals a transitional morphology in the evolution of avian flight». Proceedings of the Royal Society of London B: Biological Sciences 282 (1803): 20142864. ISSN 0962-8452. PMC 4345455. PMID 25673687. doi:10.1098/rspb.2014.2864.
- Ennos, A.; Hickson, J.; Roberts, A. (1995). «Functional morphology of the vanes of the flight feathers of the pigeon Columba livia». Journal of Experimental Biology 198: 1219-1228.
- Norberg, U.M. (1995). «How a Long Tail and Changes in Mass and Wing Shape Affect the Cost for Flight in Animals». Functional Ecology 9 (1): 48-54. JSTOR 2390089. doi:10.2307/2390089.
- Norberg, R.A. (1995). «Feather asymmetry in Archaeopteryx». Nature 374 (6519): 211. Bibcode:1995Natur.374..211M. doi:10.1038/374211a0.
- Thomas, A.L.R. (1997). «On the Tails of Birds». BioScience 47 (4): 215-225. JSTOR 1313075. doi:10.2307/1313075.
- Tucker, V.A. (1995). «Drag reduction by wing tip slots in a gliding Harris' hawk, Parabuteo unicinctus». Journal of Experimental Zoology 198: 775-781.
| Control de autoridades |
|
|---|
 Datos: Q29715232
Datos: Q29715232 Multimedia: Jianianhualong / Q29715232
Multimedia: Jianianhualong / Q29715232