Presión osmótica
La presión osmótica puede definirse como la presión que se debe aplicar a una solución para detener el flujo neto de disolvente a través de una membrana semipermeable.[1] La presión osmótica es una de las cuatro propiedades coligativas de las soluciones (dependen del número de partículas en disolución, sin importar su naturaleza). Se trata de una de las características principales a tener en cuenta en las relaciones de los líquidos que constituyen el medio interno de los seres vivos, ya que la membrana plasmática regula la entrada y salida de soluto al medio extracelular que la rodea, ejerciendo como barrera de control.
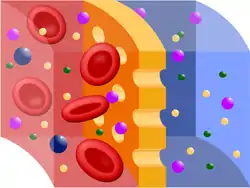
Cuando se colocan soluciones de distinta concentración, separadas por una membrana semipermeable (membrana que deja pasar las moléculas de disolvente pero no las de los solutos), las moléculas de disolvente, pasan habitualmente desde la solución con menor concentración de solutos a la de mayor concentración. Este fenómeno recibe el nombre de ósmosis, palabra que deriva del griego osmos, que significa "impulso".[2] Al suceder la ósmosis, se reduce la diferencia de presión osmótica en ambos lados de la membrana semipermeable, llegando al equilibrio de presión osmótica.
Presión osmótica de equilibrio
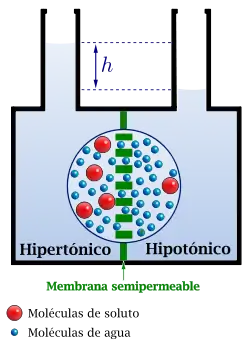
Esquema del modo de acción de la presión osmótica. En azul se representan las moléculas de disolvente y en rojo las de soluto. La disolución más concentrada se denomina hipertónica y la diluida hipotónica. La presión osmótica, consecuencia de la diferencia inicial de concentraciones, produce el pasaje de moléculas de solvente, a través de la membrana, hacia la parte de mayor concentración; de esta manera va disminuyendo la diferencia de concentración y por lo tanto también la diferencia de presión osmótica. Además aparece una diferencia de altura h y en consecuencia una presión hidrostática que, a diferencia de la osmótica, tiende a empujar el solvente hacia la parte diluida. La diferencia de altura aumenta y por lo tanto también aumenta esa presión hidrostática. Finalmente las presiones osmótica e hidrostática se igualan, deteniéndose el flujo neto de moléculas hacia la parte de mayor concentración. La diferencia de concentración se ha reducido pero no ha desaparecido: la parte concentrada sigue siendo más concentrada porque la presión hidrostática impide que la presión osmótica alcance la igualación de concentraciones.
Historia
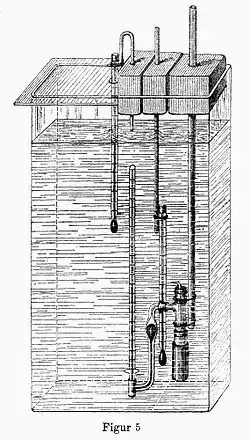
Las primeras investigaciones sobre la presión osmótica fueron realizadas en 1748 por el abad francés Jean Antoine Nollet, cuando era profesor de física en el colegio de Navarra, quien descubrió la existencia de las membranas semipermeables. Nollet obtuvo una membrana a partir de una vejiga de cerdo, colocó alcohol a un lado y agua al otro, y observó que el agua fluía a través de la vejiga para mezclarse con el alcohol, pero el alcohol no lo hacía.[3]
No obstante, el descubrimiento de la ósmosis en membranas semipermeables se le atribuye a Henri Dutrochet, considerado uno de los grandes fisiólogos del siglo XIX, en el año 1828.[4] Dutrochet descubrió este fenómeno al observar que la difusión de disolvente a través de una membrana semipermeable ocurría siempre desde la disolución de menor concentración de un soluto, el cual no podía atravesarla, hasta la disolución de mayor concentración; además, el disolvente que fluye es capaz de ejercer una presión sobre la membrana, la presión osmótica. Dutrochet construyó el primer dispositivo experimental para observar la presencia de la presión osmótica, denominado osmómetro. Al descubrir la ósmosis declaró:
Este descubrimiento que he hecho pertenece a una clase nueva de fenómenos físicos que sin duda alguna intervienen fuertemente en los procesos vitales.[3]
De este modo, Dutrochet intuyó la importancia del fenómeno en las células, las cuales absorberían o retendrían determinadas sustancias.
El inglés Thomas Graham descubrió, en 1854, que las sustancias coloidales no atravesaban ciertas membranas. Sin embargo, los primeros estudios cuantitativos datan de 1877 y se deben al alemán Wilhelm Pfeffer,[5] profesor de botánica en Tubingen y Leipzig, quien preparó una membrana semipermeable artificial precipitando hexacianoferrato (II) de cobre (II), Cu2[Fe(CN)6], sobre las paredes de un vaso poroso.[6] Pfeffer, a partir de la medición de presiones osmóticas en multitud de disoluciones de solutos no volátiles, llegó a las siguientes conclusiones:
- 1. A presión constante, la presión osmótica es directamente proporcional a la concentración de soluto.
- 2. La presión osmótica de una concentración determinada es directamente proporcional a la temperatura.
- 3. A una temperatura determinada, dos disoluciones con el mismo número de moles tienen la misma presión osmótica.[7]
Pfeffer midió la presión osmótica en soluciones de azúcar y otras moléculas orgánicas, logrando presiones de hasta algo más de 200 atmósferas.[3]
El neerlandés Jacobus Henricus van 't Hoff realizó un estudio sistemático de las propiedades coligativas de las disoluciones, que publicó en 1885.[8] En este artículo van 't Hoff fórmula una expresión, para disoluciones diluidas, que relaciona la presión osmótica con la concentración del soluto, la cual es similar a la ecuación de los gases ideales y proporciona la primera teoría para explicar la presión osmótica:

donde:
- , es la presión osmótica, en atmósferas (atm).
- , es la constante universal de los gases ideales (aunque a pesar de su nombre no sólo se aplica a gases, como es el caso). Su valor es de 0,082 atm·L·K-1·mol-1.
- , es la temperatura absoluta, en kelvin (K).
- , es la concentración molar o molaridad de la disolución, medida en mol·L-1.







Teorías
Teoría de van 't Hoff
Se han propuesto diversas teorías para explicar la causa de la ósmosis. La primera teoría fue la del bombardeo de van't Hoff, que está basada en la analogía entre la ecuación de la presión osmótica y la ley de los gases ideales. Van't Hoff describió la presión osmótica como el resultado de las colisiones de las moléculas de soluto contra la membrana semipermeable, y supuso que las moléculas de disolvente no contribuían de ninguna manera. Con este modelo, la presión osmótica de una disolución es la misma presión que un gas ideal ejercería si ocupase el mismo volumen de la disolución.
Otras teorías
La suposición en la teoría de van 't Hoff de que las moléculas del disolvente no ejercen ningún efecto sobre la presión osmótica constituye un interrogante, ya que estas partículas se encuentran golpeando continuamente la membrana semipermeable. De este problema surge otra teoría, que considera el bombardeo de las moléculas de disolvente; según esta teoría las moléculas de disolvente bombardean la membrana semipermeable de manera desigual y provocan la ósmosis y la presión osmótica, aunque la diferencia de presiones aumente con la cantidad de soluto.
Una tercera teoría explica la ósmosis en base al descenso de la presión de vapor, el cual da lugar a que el disolvente se difunda a través de la membrana hasta que se igualan las dos presiones. Si se aplica una presión igual a la presión osmótica de la disolución se saturará la destilación, y se necesitaría una presión mayor para invertir la dirección. El flujo de disolvente a través de una membrana semipermeable es análogo al flujo de disolvente en la fase vapor que se produce si se dejan, dentro de un recipiente cerrado, muestras del disolvente y de una disolución. Como la presión de vapor del disolvente es mayor, se produce una transferencia neta de disolvente hacia la disolución. El flujo de disolvente continúa hasta que todo el disolvente ha pasado a la disolución.
Magnitud de la presión osmótica
La presión osmótica, como su nombre indica, es una presión, y por tanto tiene las mismas unidades que el resto de presiones, es decir, Pascales (Pa) en el Sistema Internacional, aunque tradicionalmente también se utilizan las atmósferas (atm).
La molaridad mide la cantidad de masa del soluto por volumen de disolución. La molaridad y la presión osmótica son dos magnitudes relacionadas proporcionalmente; el aumento o disminución de una de ellas produce el mismo efecto en la otra, aunque en distinta proporción. Del mismo modo, la temperatura (medida en kelvin, K) también posee la misma relación con la presión osmótica.
A continuación se muestra una tabla con los valores de la presión osmótica correspondientes a diferentes concentraciones de sacarosa a una temperatura constante de 293 K (20 °C). La concentración se expresa en molalidad y no en molaridad, según la ecuación de Morse, pero las diferencias son mínimas.
| Presión osmótica experimental de disoluciones de sacarosa a 20 °C[9] | |||||||
| Molalidad (moles de sacarosa/kg de agua) | 0,1 | 1,0 | 2,0 | 3,0 | 4,0 | 6,0 | |
|---|---|---|---|---|---|---|---|
| Presión osmótica (atm) | 2,47 | 27,2 | 58,4 | 95,2 | 139,0 | 232,3 | |
Comparación entre presión osmótica y presión hidrostática
Al tratarse ambas de presiones, se puede comparar el valor de la presión osmótica con el de la presión hidrostática en determinadas situaciones. La presión hidrostática viene dada por:

donde ρ es la densidad del líquido (para agua: ρagua = 1 000 kg/m³), g es la aceleración de la gravedad (en la superficie terrestre: g = 9,81 m/s²), y h es la profundidad, expresada en metros.
Con esta fórmula, se puede comparar a qué profundidad de agua correspondería cualquier valor de la presión osmótica de la tabla anterior. Por ejemplo, a molalidad 1 molsac/kgagua corresponde una presión de 27,2 atm. De este modo, despejando h:

Y convirtiendo atmósferas a pascales (27,2 atm 2 750 000 Pa), obtenemos finalmente la altura:


Así, la presión osmótica sobre la membrana semipermeable que ejerce la disolución de un mol de sacarosa en un kilogramo de agua equivale a la presión a 281 m bajo el agua, un valor realmente elevado.
La presión osmótica como proceso termodinámico irreversible
Desde el punto de vista de la física, en un sistema binario no reaccionante, en el que los componentes no llevan carga eléctrica y existe una temperatura uniforme e igual para los dos reservorios, se tiene que la producción de entropía es:[10]

donde los flujos son simplemente el flujo de difusión relativo del compuesto 1 y el flujo relativo de velocidades de los componentes:

Las fuerzas termodinámicas son diferencias entre magnitudes intensivas entre los dos reservoirs: potencial químico y presión:

Las leyes fenomenológicas son:

De lo que se deduce, para una situación estacionaria () que una diferencia de concentraciones en los reservorios provoca una diferencia de presiones y viceversa. Son los fenómenos de ósmosis y ósmosis inversa, dados por la relación:


A la diferencia de presiones que provoca una determinada diferencia de concentración se denomina presión osmótica. Los coeficientes son los coeficientes fenomenológicos. Además las relaciones de Onsager-Casimir nos dicen que la paridad del proceso es positiva y .




Relación entre presión osmótica y entropía
Desde el punto de vista de la termodinámica, se puede relacionar la presión osmótica con la entropía para explicar el sentido de flujo del disolvente. El paso de disolvente hacia la disolución representa un aumento de entropía del sistema. Las moléculas de soluto aumentan su desorden al diluir la disolución mezclándose con el disolvente que atraviesa la membrana, ya que las moléculas de soluto tienen más espacio para moverse, lo que representa un mayor número de posiciones disponibles (mayor desorden), y, por tanto, una mayor entropía. También las moléculas de disolvente tienen mayor desorden en una disolución que en estado puro. Si el disolvente pasase desde la disolución al disolvente puro el orden aumentaría, lo cual disminuiría la entropía y no se cumpliría el segundo principio de la termodinámica. Todo esto demostrable matemáticamente.[11]
Relaciones con otras magnitudes físicas y químicas
Disoluciones diluidas de no electrolitos
En el caso de disoluciones diluidas de no electrolitos, se aplica directamente la ecuación de van 't Hoff. Si la presión osmótica se produce entre dos disoluciones, en lugar de entre una disolución y el disolvente puro, en la ecuación figura la diferencia de concentraciones, :


Sin embargo, la ecuación obtenida por el químico estadounidense Harmon Northrop Morse se ajusta mejor a las observaciones experimentales. Morse sustituyó la concentración expresada en molaridad de la ecuación de van 't Hoff, por la expresada en molalidad, m.[12] Así, la ecuación de Morse queda como sigue:

Realmente, la ecuación de Morse expresada en molalidad solamente es correcta si el disolvente es agua, ya que el volumen de agua expresado en litros coincide con su masa expresada en kilogramos.
| Deducción termodinámica de las ecuaciones de Morse y van 't Hoff[13] |
| Si se produce flujo neto de disolvente hacia la disolución concentrada es debido a que el sistema formado no está en equilibrio, lo cual se puede representar mediante la diferencia existente entre los potenciales químicos de la disolución y del disolvente puro:
donde * representa que el disolvente es puro. Cuando se alcanza el equilibrio, la presión en la disolución es mayor que la presión en el disolvente en una cantidad (presión osmótica). Se tendrá entonces el equilibrio: El potencial químico de la disolución se podrá expresar en función del potencial químico del disolvente y de su fracción molar (), quedando la ecuación anterior como sigue: la cual se puede reorganizar, obteniendo: El miembro de la derecha corresponde a la diferencia de potenciales químicos del disolvente puro a dos presiones diferentes que se pueden relacionar con su volumen molar, con la ecuación: Para esta ecuación se pueden realizar diversas aproximaciones:
mediante una serie de McLaurin, aproximando solamente al primer término: De esta manera, la presión osmótica queda en función de la fracción molar de soluto:
Obteniendo la presión osmótica en función del número de moles. De esta forma se obtiene la ecuación de Morse: siendo el volumen de disolvente.
donde (c es la concentración molar de la disolución). De este modo se obtiene la ecuación de van 't Hoff: |

















Disoluciones diluidas de electrolitos
Jacobus Henricus van 't Hoff descubrió que las disoluciones de electrolitos no se comportan como las soluciones de no electrolitos, e introdujo el llamado factor de van't Hoff, un factor determinado empíricamente simbolizado por la letra , y definido como el cociente entre el valor experimental de la presión osmótica media y el valor teórico que se deduce con la ecuación para disoluciones de no electrolitos:


De esta manera la nueva ecuación para la presión osmótica queda de la siguiente forma:

Este factor de corrección fue explicado por el químico sueco Svante August Arrhenius, quien entre 1883 y 1887 desarrolló la teoría de la disociación electrolítica,[14] según la cual las moléculas de los electrolitos se disocian en mayor o menor medida en iones cargados eléctricamente, capaces de transportar la corriente eléctrica, y existiendo un equilibrio entre las moléculas no disociadas y los iones. Por tanto, en disolución hay más partículas que las que se han disuelto ya que estas se rompen, se disocian, en partes más pequeñas. La presión osmótica, como las otras propiedades coligativas, depende del número total de partículas en disolución, de manera que en una disolución de electrolitos se tendrán más partículas que las disueltas, las cuales se han de tener en cuenta para calcular la concentración total. A modo de ejemplo, podemos suponer un electrolito binario como el cloruro de calcio, CaCl2, el cual alcanza el equilibrio de disociación en agua según la siguiente ecuación:

Por tanto, en disolución, hay más partículas que las disueltas inicialmente porque parte de estas partículas se han disociado en iones.[13]
Disoluciones reales
Para disoluciones concentradas, se ha de utilizar la actividad (más concretamente su logaritmo, ) en lugar de las fracciones molares, y se debe tener en cuenta la influencia de la presión en el volumen molar del disolvente (en la deducción termodinámica se considera el volumen de disolvente constante). De esta manera se obtiene una expresión más precisa, aplicable a las disoluciones concentradas que contienen un término añadido donde aparece la presión osmótica al cuadrado:


donde es el coeficiente de compresibilidad isotermo del disolvente.[13]

| Deducción termodinámica de la ley de la presión osmótica para disoluciones reales[13] |
| Partiendo de las ecuaciones obtenidas anteriormente en la deducción termodinámica de las ecuaciones de Morse y van 't Hoff:
es posible obtener el volumen molar en función del coeficiente de compresibilidad isotermo del disolvente, , y por tanto: Integrando se obtiene: Por lo que podemos igualar las expresiones correspondientes a la diferencia de potencial químico del disolvente a distinta presión: Así, despejando el logaritmo de la actividad, queda: |



En el caso de electrolitos, el coeficiente de actividad del disolvente permanece cercano a la unidad, aunque la concentración de electrolitos sea alta, y por tanto, con coeficientes de actividad del electrolito alejados de la unidad. Por este motivo, el coeficiente de actividad del disolvente no resulta adecuado para caracterizar las propiedades reales de la disolución electrolítica concentrada y se define el coeficiente osmótico (), por parte del químico danés Niels J. Bjerrum, de la siguiente manera:


Así, la expresión de la presión osmótica para electrolitos queda corregida con el coeficiente osmótico:[13]

La presión osmótica en la naturaleza
La presión osmótica en el medio interno


La ósmosis tiene una gran importancia en los seres vivos. Las células de los organismos están rodeadas por fluidos acuosos, como la sangre, la linfa, o la savia, que contienen concentraciones de diferentes solutos. Las membranas celulares son permeables al agua, al oxígeno, al nitrógeno, al dióxido de carbono, y a otras moléculas orgánicas de pequeño tamaño, como glucosa o aminoácidos, mientras que son impermeables a las moléculas poliméricas, como proteínas y polisacáridos. En cambio, los iones inorgánicos y los disacáridos, como la sacarosa, pasan muy lentamente a través de las membranas celulares.
Las células también tienen la capacidad de transportar especies químicas a través de su membrana desde una región de baja concentración de la especie a una región de concentración más elevada, en sentido contrario al del flujo espontáneo. Los mecanismos que originan este tipo de transporte, denominado transporte activo, son complejos y todavía no se conocen totalmente. Un ejemplo típico de transporte activo es el de cationes potasio, K+, hacia el interior de las células desde los líquidos circundantes, que tienen menor concentración de cationes potasio.
En ausencia de transporte activo, la membrana celular permite el paso de moléculas de agua y de todos los solutos permeables hasta que se igualen sus respectivos potenciales químicos a ambos lados de la membrana. No obstante, existe un gran número de especies, tanto en el fluido que rodea la célula como en el fluido celular o citoplasma, que no pueden atravesar la membrana. Si la concentración total de este soluto es más grande en el fluido que rodea la célula, esta perderá agua por ósmosis, y se dice que el fluido circundante es hipertónico respecto al fluido celular (tiene mayor presión osmótica). En caso contrario, cuando la concentración total del soluto que no puede atravesar la membrana es mayor en el fluido de la célula, esta ganará agua del líquido hipotónico circundante (de menor presión osmótica). Cuando no se produce transferencia neta de agua entre el fluido celular y el que rodea la célula, se dice que los dos fluidos son isotónicos, es decir, tienen la misma presión osmótica. La sangre y la linfa son aproximadamente isotónicos respecto de las células de un organismo.
Los líquidos de las inyecciones contienen una disolución salina isotónica con la sangre, porque si se inyectara agua directamente, los eritrocitos de la sangre la absorberían por ósmosis hasta estallar.[7]
Osmorreceptores

Existen unas células especializadas en detectar cambios de la presión osmótica en los líquidos corporales (plasma sanguíneo, fluido intercelular...), llamadas osmorreceptores. Los osmorreceptores pueden localizarse en algunas estructuras, como en el hipotálamo, en las arterias, entre otros lugares. Mediante estas células, el organismo puede regular la concentración de sales en sus fluidos, la osmolalidad.[15] En los seres humanos, son osmorreceptores los núcleos supraóptico o paraventricular. Por medio de las hormonas antidiuréticas (también llamada vasopresina, ADH o pitresina), los osmorreceptores inhiben la diuresis con tal de mantener la presión osmótica de la sangre correspondiente a una osmolalidad media de 285 osmol/kg. Es suficiente una variación de tan solo 3 osmol/kg para activar este mecanismo.[16]
La absorción de agua en los vegetales

Los vegetales utilizan la presión osmótica para hacer ascender agua a través del xilema desde las raíces hasta las hojas. Así se ha comprobado en los arces el comenzar la primavera y se inicia el movimiento de la savia. Las raíces tienen una gran concentración de azúcares almacenados, producidos durante el verano. Cuando se funde la nieve, el agua llega en grandes cantidades a las raíces y entra dentro de ellas a través de pelos absorbentes, y por efecto de la presión osmótica la savia sube hacia las hojas. Sin embargo, en la mayoría de los vegetales no es posible explicar la ascensión de la savia hasta las hojas por medio de la presión osmótica. En las partes más elevadas del vegetal, la savia asciende debido al descenso de presión provocado por la transpiración de las hojas. En ocasiones, en lugar de que las raíces presenten grandes concentraciones de azúcares, se encuentran rodeadas de agua muy salada, produciéndose el fenómeno conocido como ósmosis inversa; este es el caso de los mangles.[17]
Organismos osmófilos
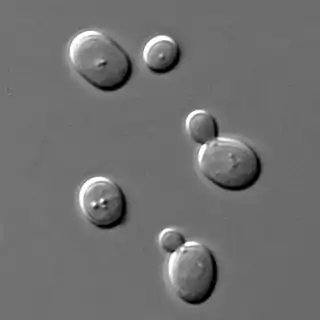
Se denominan osmófilos los organismos extremófilos que pueden crecer en ambientes con una alta concentración de azúcares.[15] Casi todos los microorganismos osmófilos son levaduras. Los osmófilos son similares a los organismos halófilos (medios salinos). La alta concentración de azúcar representa un factor limitante en el crecimiento de muchos microorganismos, pero los osmófilos logran protegerse contra la elevada presión osmótica por medio de la síntesis de osmoprotectores, como alcoholes y aminoácidos. Estos organismos son importantes porque causan el deterioro de productos como los zumos de frutas, los jarabes o la miel, además de tener aplicaciones industriales como la fermentación de la cerveza. Algunos ejemplos de este tipo de microorganismos osmófilos son: Saccharomyces rouxii, Saccharomyces bailii, Debaryomyces, y Saccharomyces cerevisiae, la levadura de la cerveza. "Saccharomyces" deriva del griego, que significa "moho del azúcar".
Aplicaciones
Científicas
Una de las aplicaciones científicas de la presión osmótica es la determinación de masas moleculares o masas molares, de macromoléculas. A partir de la ecuación de van't Hoff se puede despejar la masa molar, M, del soluto disuelto:

Despejando la masa molar, ésta queda en función de constantes y variables que se pueden medir en el laboratorio: la masa disuelta de soluto, la constante de los gases ideales, R, la temperatura absoluta, T, el volumen de disolución, V, y la presión osmótica, :

Analíticas
En los laboratorios se puede determinar la concentración total de partículas en disoluciones osmóticamente activas, lo que se conoce como osmolaridad u osmolalidad, midiendo la presión osmótica. Los aparatos que se utilizan se denominan osmómetros de membrana. Se trata de aparatos automatizados que permiten la realización de las determinaciones de presión osmótica de forma rápida.
Industriales

En la industria se emplea la ósmosis inversa. Se trata de un procedimiento que invierte la difusión normal de disolvente desde la disolución diluida hacia la disolución concentrada, aplicando una presión sobre la disolución concentrada que supera a la presión osmótica, permitiendo el paso de disolvente desde la disolución concentrada a la diluida.
Los campos de aplicación de la ósmosis inversa son cada vez más numerosos. Las áreas de la ingeniería que más se han visto beneficiadas con estas aplicaciones son principalmente las industrias de los alimentos, del papel, la biomedicina, la petroquímica, la nuclear y, entre otras, la de separación o purificación de gases, donde se obtiene hidrógeno de la disociación del amonio, sulfuro de hidrógeno del gas natural de las refinerías, helio del gas natural, así como el enriquecimiento de oxígeno a partir del aire. También cabe destacar la recuperación de sustancias valiosas en fluidos de deshechos industriales. No obstante, la aplicación más importante es la desalinización del agua salada marina para su potabilización, lo cual ha permitido abastecer con agua de calidad a poblaciones situadas en lugares donde los recursos hidráulicos son insuficientes.
Véase también
Referencias
- Voet, Donald; Voet, Judith G.; Pratt, Charlotte W. (2001). Fundamentals of biochemistry. Wiley. p. 30. ISBN 9780471417590.
- Dorland (2005). Dorland Diccionario enciclopédico ilustrado de medicina (30ª edición). Elsevier España. p. 1407. ISBN 978-84-8174-790-4. Consultado el 29 de noviembre de 2009.
- Castillo, L.F. (1997). El fenómeno mágico de la ósmosis. Fondo de Cultura Económica. ISBN 968-16-5241-X. Consultado el 2 de diciembre de 2009.
- Dutrochet, H. (1828). Nouvelles Recherches sur l'Endosmose et l'Exosmose, suivies de l'application expérimentales de ces actions physiques à la solution du problême de l'irritabilité végétale (en francés). Universidad de Oxford. Consultado el 2 de diciembre de 2009.
- Pfeffer, W. (1921). Osmotische Untersuchungen; Studien zur Zellmechanik (en alemán). W. Engelmann. Consultado el 2 de diciembre de 2009.
- Babor, J.A.; Ibarz, J. (1979). Química General Moderna (8ª edición). Marín. ISBN 84-7102-997-9.
- Brillas, Enric (2004). Conceptes de termodinàmica química i cinètica (en catalán). Edicions Universitat Barcelona. ISBN 978-8447528424. Consultado el 15 de diciembre de 2009.
- Van 't Hoff, J.H. (1885). L'équilibre chimique dans les systèmes gazeux ou dessus à l'état dilué.
- Glasstone, S. (1977). Termodinámica para químicos (5a edición). Aguilar. ISBN 84-03-20059-5.
- S.R. de Groor & P.Mazur. Non-equilibrium thermodynamics, 1962. Dover publications. pg. 435.
- Gillespie, R.J.; Beltrán, A. (1990). Química. Reverté. ISBN 84-291-7188-6.
- Morse, H.N. (1914). The osmotic pressure of aqueous solutions, report on investigations made in the Chemical Laboratory of the Johns Hopkins University during the years 1899-1913 (en inglés). Washington, Carnegie Inst. p. 222. Consultado el 4 de diciembre de 2009.
- Rodríguez, J.A.; Ruiz, J.J.; Urieta, J.S. (2000). Termodinámica química. Síntesis. ISBN 84-7738-581-5.
- Arrhenius, S. (1887). «On the Dissociation of Substances Dissolved in Water». Zeitschrift fur physikalische Chemie I: 631. Traducción al inglés. Consultado el 4 de diciembre de 2009.
- Attwood, Teresa K.; Cammack, Richard (2006). Oxford dictionary of biochemistry and molecular biology (en inglés) (2ª edición). Oxford University Press. ISBN 0198529171. Consultado el 5 de diciembre de 2009.
- Lote, Christopher J. (2000). Principles of renal physiology (en inglés) (4ª edición). Springer. pp. 97-100. ISBN 978-0792360742. Consultado el 5 de diciembre de 2009.
- Cromer, A.H. (1998). Física para las ciencias de la vida (2ª edición). Reverté. p. 221. ISBN 842911808X. Consultado el 5 de diciembre de 2009.
Bibliografía
- Castillo, L.F. (1997). El fenómeno mágico de la ósmosis. Fondo de Cultura Económica. ISBN 968-16-5241-X.
Enlaces externos
- Simulación interactiva de un proceso de ósmosis.
- Simulación interactiva de un proceso de ósmosis (para ordenadores poco potentes).
| Control de autoridades |
|
|---|
 Datos: Q193135
Datos: Q193135 Multimedia: Osmotic pressure / Q193135
Multimedia: Osmotic pressure / Q193135