Macrófago
Los macrófagos (griego "gran comedor") (abreviado como Mφ, MΦ o MP) son células del sistema inmunitario que se localizan en los tejidos. Proceden de células precursoras de la médula ósea que se dividen dando monocitos (un tipo de leucocito), que tras atravesar el epitelio de los capilares y penetrar en el tejido conjuntivo se convierten en macrófagos. También pueden ingerir y destruir bacterias, células dañadas y eritrocitos gastados. Este proceso se llama fagocitosis.[1]
| Macrófago | ||
|---|---|---|
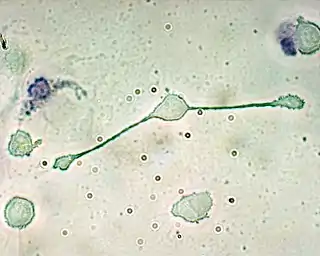 Un macrófago de ratón formando "pato" (pseudópodos) para engullir dos partículas, posiblemente patógenas. | ||
| Nombre y clasificación | ||
| Latín | Macrophagocytus | |
| TH | H2.00.03.0.01007 | |
| TH | H2.00.03.0.01007 | |
|
| ||
Descubrimiento de los macrófagos
Elie Metchnikoff, nacionalidad rusa, recibió el Premio Nobel de Fisiología o Medicina por sus trabajos sobre la inmunidad en el año 1908. Descubrió que ciertas células aisladas digerían partículas que él había introducido en el tubo digestivo de las larvas de peces con las que estudiaba. A estas células las llamó fagocitos, y más tarde los identificó como glóbulos blancos que formaban la primera línea de defensa contra las infecciones en los seres vivos.
El término macrófago fue asignado por Aschoff en 1924 a un conjunto de células componentes del sistema fagocítico mononuclear que está formado no solo por monocitos, macrófagos e histiocitos, sino también por fibroblastos, células endoteliales y células reticulares. Después de 1969, se definió el concepto de sistema fagocítico mononuclear, formado por una variedad de macrófagos, derivados de monocitos procedentes de la médula ósea, abandonándose el concepto de sistema retículo endotelial, que está constituido por células distintas funcional e inmunológicamente.
Formación de los macrófagos
Los macrófagos proceden de los monocitos.[2] Estos se forman en la médula ósea procedentes de células pluripotenciales de la serie granulocítico-monocítica, gracias al factor de crecimiento GM-CSF (granulocyte macrophage colony stimulating factor) y otras citoquinas, como la interleucina 3 (IL-3).
Cuando estos factores de crecimiento están presentes en la médula ósea, la célula progenitora prolifera y se diferencia a promonocitos, células que en divisiones celulares posteriores producirán monocitos.
La diferenciación celular de la célula madre está asociada con la expresión de receptores de membrana para citocinas específicas. Los monocitos permanecen en la médula ósea menos de 24 horas, después pasan a la circulación sanguínea y son distribuidos por todo el cuerpo. En esta etapa, se visualizan al microscopio en los frotis de sangre periférica como células de mayor tamaño, cuyo diámetro oscila entre 15 a 30 μm y poseen una alta relación núcleo/citoplasma.
En adultos sanos normales, la vida media de un monocito circulante se estima en 70 horas y en una proporción del 1 al 6 por ciento del total de leucocitos en un recuento normal en sangre periférica.
Una vez los monocitos salen de los capilares sanguíneos y se localizan en los tejidos se transforman en macrófagos. Esta diferenciación de monocito a macrófago afecta a gran cantidad de cambios como que la célula aumenta su tamaño de 5 a 10 veces, sus orgánulos incrementan tanto su número como su complejidad, adquiere capacidad fagocítica, produce altas concentraciones de enzimas líticas y empieza a secretar gran variedad de sustancias solubles que realizan diferentes funciones.
Los macrófagos están habitualmente en estado de reposo, pueden ser activados por gran variedad de estímulos durante la respuesta inmune. La fagocitosis de antígenos sirve como estímulo inicial; sin embargo, los macrófagos y su actividad pueden aumentarse por citocinas secretadas por linfocitos T colaboradores, o mediante el contacto con los mismos. Uno de los más potentes activadores de macrófagos es el interferón gamma. También son capaces de reconocer patrones moleculares asociados a patógenos.
Función de los macrófagos
- Fagocitosis: la función principal de los macrófagos es la de fagocitar todos los cuerpos extraños que se introducen en el organismo como las bacterias y sustancias de desecho de los tejidos. Los macrófagos son fagocitos junto con los neutrófilos y otras células. Los macrófagos tienen la capacidad de quimiotaxis, es decir la de ser atraídos y desplazados hacia una determinada localización por la presencia de determinados factores quimiotácticos para monocitos como interleucina-I, trombina, factor de crecimiento derivado de las plaquetas, factor del complemento C5a, fragmentos de colágeno, elastina, fibronectina, calicreína, activador del plasminógeno, inmunoglobulinas y leucotrienos.
- Inflamación: Los macrófagos forman parte del sistema inmunitario innato, es decir, inician una respuesta natural contra los microorganismos, porque los macrófagos expresan receptores de membrana para numerosas moléculas bacterianos, por ejemplo: receptor para lipopolisacáridos (CD14), receptores C11b/CD18, receptores para manosas, y receptor para glúcidos entre otros. Los macrófagos de los vertebrados y de los invertebrados participan en gran medida de la respuesta inmune innata a infecciones gracias a sus receptores scavengers o barredores, que poseen una especificidad a ligandos muy amplia como: lipoproteínas, proteínas, poli y oligonucleótidos, polisacáridos aniónicos, fosfolípidos y otras moléculas.
- Presentación de antígenos: cuando los macrófagos fagocitan un microbio, procesan y sitúan sus antígenos en la superficie externa de su membrana plasmática, donde serán reconocidos por los linfocitos T colaboradores; tras el reconocimiento, los T producen linfoquinas que activan a los linfocitos B. Por eso los macrófagos forman parte de las llamadas células presentadoras de antígenos, ya que poseen en sus membranas moléculas del complejo mayor de histocompatibilidad (MHC) de clase II. Los linfocitos B activados producen y liberan anticuerpos específicos a los antígenos presentados por el macrófago. Estos anticuerpos se adhieren a los antígenos de los microbios o de células invadidas por virus y así atraen con mayor avidez a los macrófagos para fagocitarlos.
- Reparación de tejidos: Los macrófagos además ingieren células muertas del anfitrión y participan en la reparación de tejidos dañados tras la reacción inmunitaria.
- Hemostasia: el macrófago produce una serie de sustancias que participan en la coagulación como son: proteína C, trombomodulina, factor tisular, factor VII, factor XIII y el inhibidor del activador del plasminógeno.
Sistema fagocítico mononuclear
El conjunto de células formado por los precursores de la médula ósea, los monocitos circulantes en la sangre y los macrófagos tisulares, se denomina sistema fagocítico mononuclear.
Los macrófagos reciben diferentes nombres según el lugar donde se encuentren, debido a que históricamente no se reconocían como el mismo tipo celular. Así, los macrófagos se llaman:[3]
- Osteoclastos: son los macrófagos del hueso, se encuentran principalmente en las trabéculas del hueso esponjoso.
- Microglía: es el conjunto de los macrófagos del sistema nervioso central.
- Células de Kupffer: son los macrófagos del hígado.
- Células espumosas: son las células dominantes de la ateroesclerosis, los macrófagos al fagocitar el colesterol cambian su estructura y se convierten en estas.
- Célula gigante de Langhans: Macrófago formado por la fusión de células epitelioides, se encuentran en la piel además están presente en las enfermedades de tipo granulomatoso.
- Histiocito: son los macrófagos del tejido conjuntivo.
- Células mesangiales intraglomerulares: son los macrófagos del riñón, también se les llama Macrófago renal residente.
- Macrófagos alveolares, células de polvo o neumocitos tipo III: son los macrófagos de los alveolos pulmonares.
- Macrófago de pulpa roja: son los macrófagos del bazo.
- Macrófago de cavidad peritoneal: son los macrófagos de la cavidad peritoneal.
- Macrófago lámina propia: son los macrófagos del intestino.
- Macrófago del seno subcapsular: son los macrófagos de los nódulos linfáticos.
- Macrófago de médula ósea: son los macrófagos de la médula ósea.
Subtipos de Macrófagos
Hay varias formas activadas de macrófagos. A pesar de un espectro de formas de activar los macrófagos, hay dos grupos principales designados M1 y M2. Macrófagos M1: como se mencionó anteriormente (anteriormente denominados macrófagos activados clásicamente),[4] los macrófagos "asesinos" M1 son activados por LPS e IFN-gamma, y secretan altos niveles de IL-12 y niveles bajos de IL-10. Los macrófagos M1 tienen funciones proinflamatorias, bactericidas y fagocíticas.[5] Por el contrario, la designación M2 de "reparación" (también denominada macrófagos activados alternativamente) se refiere ampliamente a los macrófagos que funcionan en procesos constructivos como la cicatrización de heridas y la reparación de tejidos, y aquellos que desactivan la activación dañina del sistema inmunológico al producir citocinas antiinflamatorias como IL-10. M2 es el fenotipo de los macrófagos tisulares residentes y puede elevarse aún más mediante IL-4. Los macrófagos M2 producen altos niveles de IL-10, TGF-beta y niveles bajos de IL-12. Los macrófagos asociados a tumores son principalmente del fenotipo M2 y parecen promover activamente el crecimiento tumoral.[6]
Los macrófagos existen en una variedad de fenotipos que están determinados por el papel que desempeñan en la maduración de la herida. Los fenotipos se pueden separar predominantemente en dos categorías principales; M1 y M2. Los macrófagos M1 son el fenotipo dominante observado en las primeras etapas de la inflamación y son activados por cuatro mediadores clave: interferón-γ (IFN-γ), factor de necrosis tumoral (TNF) y patrones moleculares asociados al daño (DAMP). Estas moléculas mediadoras crean una respuesta proinflamatoria que, a cambio, produce citocinas proinflamatorias como la IL-6 y el TNF. A diferencia de los macrófagos M1, los macrófagos M2 secretan una respuesta antiinflamatoria mediante la adición de interleucina-4 o interleucina-13. También juegan un papel en la cicatrización de heridas y son necesarios para la revascularización y reepitelización. Los macrófagos M2 se dividen en cuatro tipos principales según sus funciones: M2a, M2b, M2c y M2d. La forma en que se determinan los fenotipos M2 aún está en discusión, pero los estudios han demostrado que su entorno les permite adaptarse al fenotipo más apropiado para curar la herida de manera eficiente.[5]
Los macrófagos M2 son necesarios para la estabilidad vascular. Producen factor de crecimiento epitelial vascular A y TGF-β1.[5] Hay un cambio de fenotipo de macrófagos M1 a M2 en las heridas agudas, sin embargo, este cambio se ve afectado en las heridas crónicas. Esta desregulación da como resultado macrófagos M2 insuficientes y sus factores de crecimiento correspondientes que ayudan en la reparación de heridas. Con la falta de estos factores de crecimiento/citocinas antiinflamatorias y una sobreabundancia de citocinas proinflamatorias de los macrófagos M1, las heridas crónicas no pueden sanar de manera oportuna. Normalmente, después de que los neutrófilos comen desechos/patógenos, realizan apoptosis y se eliminan. En este punto, la inflamación no es necesaria y M1 se somete a un cambio a M2 (antiinflamatorio). Sin embargo, la desregulación ocurre cuando los macrófagos M1 no pueden/no fagocitan los neutrófilos que han sufrido apoptosis, lo que lleva a una mayor migración e inflamación de los macrófagos.[5]
Tanto los macrófagos M1 como M2 juegan un papel en la promoción de aterosclerosis. Los macrófagos M1 promueven la aterosclerosis por inflamación. Los macrófagos M2 pueden eliminar el colesterol de los vasos sanguíneos, pero cuando el colesterol se oxida, los macrófagos M2 se vuelven apoptóticos contribuyendo a la placa ateromatosa de la aterosclerosis.[7][8]
Función en la regeneración del músculo
El primer paso para comprender la importancia de los macrófagos en la reparación, el crecimiento y la regeneración de los músculos es que hay dos "ondas" de macrófagos con el inicio del uso de músculos dañinos subpoblaciones que tienen y no tienen una influencia directa en la reparación del músculo. La onda inicial es una población fagocítica que aparece durante los períodos de mayor uso de los músculos que son suficientes para causar la lisis de la membrana muscular y la inflamación de la membrana, que pueden ingresar y degradar el contenido de las fibras musculares lesionadas.[9][10][11] Estos macrófagos fagocíticos invasores tempranos alcanzan su concentración más alta aproximadamente 24 horas después del inicio de alguna forma de lesión o recarga de las células musculares.[12] Su concentración disminuye rápidamente después de 48 horas.[10] El segundo grupo son los tipos no fagocíticos que se distribuyen cerca de las fibras regenerativas. Estos alcanzan su punto máximo entre dos y cuatro días y permanecen elevados durante varios días durante la esperanzadora reconstrucción muscular.[10] La primera subpoblación no tiene ningún beneficio directo para la reparación del músculo, mientras que el segundo grupo no fagocítico sí.
Se cree que los macrófagos liberan sustancias solubles que influyen en la proliferación, diferenciación, crecimiento, reparación y regeneración del músculo, pero en este momento se desconoce el factor que se produce para mediar estos efectos.[12] Se sabe que la participación de los macrófagos en la promoción de la reparación de tejidos no es específica del músculo; se acumulan en numerosos tejidos durante la fase del proceso de curación posterior a la lesión.[13]
Cicatrización de la herida
Los macrófagos son esenciales para la cicatrización de heridas.[14] Reemplazan a los neutrófilos polimorfonucleares como las células predominantes en la herida al segundo día después de la lesión.[15] Atraídos al sitio de la herida por factores de crecimiento liberados por las plaquetas y otras células, los monocitos del torrente sanguíneo ingresan al área a través de las paredes de los vasos sanguíneos.[16] La cantidad de monocitos en la herida alcanza su punto máximo entre un día y un día y medio después de que ocurre la lesión. Una vez que están en el sitio de la herida, los monocitos maduran y se convierten en macrófagos. El bazo contiene la mitad de los monocitos del cuerpo en reserva listos para ser desplegados en el tejido lesionado.[17][18]
La función principal de los macrófagos es fagocitar las bacterias y el tejido dañado,[14] y también desbridamiento tejido dañado al liberar proteasas.[19] y también desbridan el tejido dañado mediante la liberación de proteasas. Los macrófagos también secretan una serie de factores como factores de crecimiento y otras citocinas, especialmente durante el tercer y cuarto días posteriores a la herida. Estos factores atraen las células involucradas en la etapa de proliferación de la curación al área.Los macrófagos también pueden restringir la fase de contracción.Los macrófagos son estimulados por el bajo contenido de oxígeno de su entorno para producir factores que inducen y aceleran la angiogénesis y también estimulan las células que reepitelizan la herida, crean tejido de granulación y establecen una nueva matriz extracelular. Al secretar estos factores, los macrófagos contribuyen a impulsar el proceso de curación de heridas a la siguiente fase.
Referencias
- Diccionario de biología. Editorial Norma S.A. 1982. p. 206.
- González, Ma. Isabel Crespo (2016). Fisiopatología general. Paraninfo. ISBN 9788428337984. Consultado el 11 de septiembre de 2017.
- Cruz-Reyes, Alejandro; Camargo-Camargo, Blanca (2000). Glosario de términos en parasitología y ciencias afines. Plaza y Valdes. ISBN 9789688568781. Consultado el 11 de septiembre de 2017.
- «The lymphocyte story». New Scientist (1605). Consultado el 13 de septiembre de 2007.
- Hesketh M, Sahin KB, West ZE, Murray RZ (July 2017). «Macrophage Phenotypes Regulate Scar Formation and Chronic Wound Healing». International Journal of Molecular Sciences 18 (7): 1545. PMC 5536033. PMID 28714933. doi:10.3390/ijms18071545.
- Galdiero MR, Garlanda C, Jaillon S, Marone G, Mantovani A (July 2013). «Tumor associated macrophages and neutrophils in tumor progression». Journal of Cellular Physiology 228 (7): 1404-12. PMID 23065796. doi:10.1002/jcp.24260.
- Hotamisligil GS (April 2010). «Endoplasmic reticulum stress and atherosclerosis». Nature Medicine 16 (4): 396-9. PMC 2897068. PMID 20376052. doi:10.1038/nm0410-396.
- Oh J, Riek AE, Weng S, Petty M, Kim D, Colonna M, Cella M, Bernal-Mizrachi C (April 2012). «Endoplasmic reticulum stress controls M2 macrophage differentiation and foam cell formation». The Journal of Biological Chemistry 287 (15): 11629-41. PMC 3320912. PMID 22356914. doi:10.1074/jbc.M111.338673.
- Krippendorf BB, Riley DA (January 1993). «Distinguishing unloading- versus reloading-induced changes in rat soleus muscle». Muscle & Nerve 16 (1): 99-108. PMID 8423838. doi:10.1002/mus.880160116.
- St Pierre BA, Tidball JG (July 1994). «Differential response of macrophage subpopulations to soleus muscle reloading after rat hindlimb suspension». Journal of Applied Physiology 77 (1): 290-7. PMID 7961247. doi:10.1152/jappl.1994.77.1.290.
- Tidball JG, Berchenko E, Frenette J (April 1999). «Macrophage invasion does not contribute to muscle membrane injury during inflammation». Journal of Leukocyte Biology 65 (4): 492-8. PMID 10204578. doi:10.1002/jlb.65.4.492.
- Schiaffino S, Partridge T (2008). Skeletal Muscle Repair and Regeneration. Advances in Muscle Research 3.
- Bréchot N, Gomez E, Bignon M, Khallou-Laschet J, Dussiot M, Cazes A, Alanio-Bréchot C, Durand M, Philippe J, Silvestre JS, Van Rooijen N, Corvol P, Nicoletti A, Chazaud B, Germain S (2008). «Modulation of macrophage activation state protects tissue from necrosis during critical limb ischemia in thrombospondin-1-deficient mice». PLOS ONE 3 (12): e3950. Bibcode:2008PLoSO...3.3950B. PMC 2597179. PMID 19079608. doi:10.1371/journal.pone.0003950.
- de la Torre J., Sholar A. (2006). Wound healing: Chronic wounds. Emedicine.com. Accessed 20 January 2008.
- Expert Reviews in Molecular Medicine. (2003). The phases of cutaneous wound healing Archivado el 17 de diciembre de 2008 en Wayback Machine.. 5: 1. Cambridge University Press. Accessed 20 January 2008.
- Lorenz H.P. and Longaker M.T. (2003). Wounds: Biology, Pathology, and Management Archivado el 24 de agosto de 2014 en Wayback Machine.. Stanford University Medical Center. Accessed 20 January 2008.
- Swirski FK, Nahrendorf M, Etzrodt M, Wildgruber M, Cortez-Retamozo V, Panizzi P, Figueiredo JL, Kohler RH, Chudnovskiy A, Waterman P, Aikawa E, Mempel TR, Libby P, Weissleder R, Pittet MJ (July 2009). «Identification of splenic reservoir monocytes and their deployment to inflammatory sites». Science 325 (5940): 612-6. Bibcode:2009Sci...325..612S. PMC 2803111. PMID 19644120. doi:10.1126/science.1175202.
- Jia T, Pamer EG (July 2009). «Immunology. Dispensable but not irrelevant». Science 325 (5940): 549-50. Bibcode:2009Sci...325..549J. PMC 2917045. PMID 19644100. doi:10.1126/science.1178329.
- Deodhar AK, Rana RE (1997). «Surgical physiology of wound healing: a review». Journal of Postgraduate Medicine 43 (2): 52-6. PMID 10740722.
Enlaces externos
- Esta obra contiene una traducción derivada de «Macrophage» de Wikipedia en inglés, publicada por sus editores bajo la Licencia de documentación libre de GNU y la Licencia Creative Commons Atribución-CompartirIgual 4.0 Internacional.
Véase también
- Granulocito
- Pidotimod
- Polarización de los macrófagos
| Control de autoridades |
|
|---|
 Datos: Q184204
Datos: Q184204 Multimedia: Macrophages / Q184204
Multimedia: Macrophages / Q184204