Nemegtomaia barsboldi
Nemegtomaia barsboldi es la única especie conocida del género extinto Nemegtomaia ("madre de Nemegt") de dinosaurio terópodo ovirraptórido, que vivió a finales del período Cretácico, hace aproximadamente 70 millones de años, en el Maastrichtiense, en lo que hoy es Asia. El primer espécimen se encontró en 1996 y se convirtió en la base del nuevo género y especie N. barsboldi en 2004. El nombre original del género era Nemegtia, pero se cambió a Nemegtomaia en 2005, ya que el nombre anterior estaba ocupado. La primera parte del nombre genérico se refiere a la cuenca de Nemegt , donde se encontró el animal, y la segunda parte significa "buena madre", en referencia al hecho de que se sabe que los oviraptoridos criaron sus huevos. El nombre específico honra al paleontólogo Rinchen Barsbold. Se encontraron dos especímenes más en 2007, uno de los cuales se encontró en la parte superior de un nido con huevos, pero el dinosaurio había recibido su nombre de género antes de que se encontrara asociado con los huevos.

| Nemegtomaia barsboldi | ||
|---|---|---|
| Rango temporal: 70 Ma Cretácico superior | ||
 | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Sauropsida | |
| Superorden: | Dinosauria | |
| Orden: | Saurischia | |
| Suborden: | Theropoda | |
| Infraorden: | Oviraptorosauria | |
| Superfamilia: | Caenagnathoidea | |
| Familia: | Oviraptoridae | |
| Subfamilia: | Oviraptorinae | |
| Género: |
Nemegtomaia Lü et al., 2004 [originalmente Nemegtia, preocupado] | |
| Especie: |
N. barsboldi Lü et al., 2004 | |
| Sinonimia | ||
| ||
Se estima que la Nemegtomaia medía alrededor de 2 metros de longitud y pesaba 40 kilogramos. Como un oviraptorosauriano, habría sido emplumado. Tenía un cráneo profundo, estrecho y corto, con una cresta arqueada. No tenía dientes, tenía un hocico corto con un pico similar a un loro y un par de proyecciones similares a dientes en el paladar. Tenía tres dedos, el primero era más grande y tenía una garra fuerte. Nemegtomaia está clasificada como miembro de la subfamilia de oviraptóridos Heyuanninae, y es el único miembro conocido de este grupo con una cresta craneal . Aunque Nemegtomaia se ha utilizado para sugerir que los oviraptorosaurianos fueron pájaros no voladores, el clado generalmente se considera un grupo de dinosaurios no aviarios.
El espécimen de Nemegtomaia anidado se colocó encima de lo que probablemente era un anillo de huevos, con los brazos cruzados sobre ellos. Ninguno de los huevos está completo, pero se estima que tienen 5 a 6 centímetros de ancho y 14 a 16 centímetros de largo cuando están intactos. El espécimen se encontró en un área estratigráfica que indica que Nemegtomaia prefirió anidar cerca de arroyos que proporcionarían sustrato blando, arenoso y alimento. Nemegtomaia puede haber protegido sus huevos cubriéndolos con sus plumas de cola y alas. El esqueleto del espécimen de anidación tiene daños que indican que fue comido por escarabajos de la piel.. La dieta de los oviraptóridos es incierta, pero sus cráneos son más similares a otros animales que se sabe o se cree que han sido herbívoros . Nemegtomaia es conocida por las formacionesNemegt y Baruun Goyot, que se cree que representan ambientes húmedos y áridos que coexistieron en la misma área.
Descripción

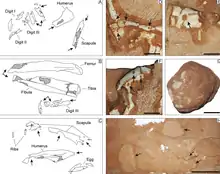
Se estima que Nemegtomaia medía alrededor de 2 metros de longitud y pesaba 40 kilogramos, un tamaño extrapolado de parientes más conocidos. Como un oviraptorosauriano, habría sido emplumado. Las espinas neurales de las vértebras del cuello eran cortas, y los arcos neurales tenían una apariencia en forma de x. Las tres del medio de estas vértebras eran las más grandes. La escápula parece haber sido de 185 milímetros de longitud total. El húmero tenía una fosa en una posición similar a las aves modernas, pero atípica entre los oviraptorosaurianos, y parece haber medido 152 milímetros de largo. El radio, de sección transversal ovalada, y puede haber medido 144 milímetros de largo. El primer dedo era relativamente grande y tenía una fuerte garra y era más macizo que los otros dos dedos. El segundo dedo era un poco más largo que el primero, y el tercer dedo era el más pequeño. El margen superior del ilion de la pelvis era recto, y aunque ambas iliones estaban cerca una de la otra, no estaban fusionadas. El eje púbico se volvió hacia atrás. Se estima que el fémur tenía 286 milímetros de largo y la tibia de la parte inferior de la pierna 317 milímetros de largo.[1][2]
El cráneo de Nemegtomaia era profundo, estrecho y corto, en comparación con el resto del cuerpo y alcanzaba 179 milímetros de longitud. Tenía una cresta bien desarrollada, formada por los huesos nasales y premaxilares, principalmente el último, del hocico. El margen frontal casi vertical de la cresta del holotipo formó un ángulo de casi 90 grados con el margen superior del cráneo. En comparación con otros oviraptóridos, los procesos nasales de las premaxilas apenas eran visibles cuando se observaban desde arriba, donde se conectaban con los huesos nasales en los puntos más altos de la cresta. La cresta se extendía hacia atrás y hacia abajo, formando un arco redondo en el punto más alto. El diámetro de la órbita ocular fue de 52 milímetros, los ojos se veían grandes debido a lo pequeño del cráneo. La cavidad antorbital frente al ojo consistía en dos fenestras, una gran fenestra antorbital en la parte posterior y una pequeña fenestra maxilar en la parte delantera.En Nementomaia era distinta de otros oviraptóridos en que el hueso frontal en la línea media del cráneo tenía aproximadamente un 25% de la longitud del hueso parietal de adelante hacia atrás. Las narinas eran relativamente pequeñas y se colocaban en lo alto del cráneo.[1][2]
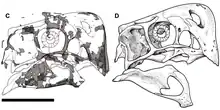
Las mandíbulas de Nemegtomaia no tenían dientes, y al igual que otros dinosaurios oviraptóridos, tenía un hocico corto con un pico profundo, robusto y algo parecido a un loro. Tenía un paladar duro formado por las premaxilas, vómitos y maxilares , como otros oviraptoridos. El paladar era fuertemente cóncavo hacia abajo y tenía una hendidura en la parte central. Al igual que en otros oviraptóridos, tenía un par de proyecciones en forma de diente en el paladar que se dirigían hacia abajo, una característica que se ha llamado "pseudo-dientes". Nemegtomaia tenía pequeños agujeros en los lados de la sutura entre los premaxilares en la parte frontal del hocico, que pueden haber sido aberturas de nutrientes y que indican la presencia de un pico queratinoso. La mandíbula inferior era corta y profunda, con una superficie inferior convexa y alcanzaba 153 milímetros de longitud. El hueso dentario de la mandíbula inferior alcanzó 50 milímetros en su punto más alto. La sínfisis mandibular donde se conectan las dos mitades de la mandíbula inferior fue corta, profunda y muy neumática, con espacios de aire. La fenestra mandibular era grande y estaba ubicada en la parte frontal de la mandíbula inferior.[1][2][3][4] Como en la mayoría de los otros oviraptoridos, la parte frontal de la mandíbula inferior estaba doblada hacia abajo.[5]
Descubrimiento e investigación
En 1996, el paleontólogo japonés Yoshitsugu Kobayashi, como parte del equipo del "Proyecto de Dinosaurio Internacional de las Tierras Altas de Mongolia", encontró un esqueleto incompleto de un dinosaurio oviraptórido en la formación Nemegt del Desierto de Gobi en el suroeste de Mongolia. El espécimen, MPC-D 100/2112 en el Centro Paleontológico de Mongolia, anteriormente PC y GIN 100/2112, consiste en un cráneo casi completo y un esqueleto parcial, que incluye vértebras cervicales, dorsales, sacras y caudales, una escápula izquierda, los extremos inferiores de ambos húmero, el radio derecho, ambas iliones, los extremos superiores de ambos huesos púbicos, ambos isquiones y el extremo superior de un fémur. El espécimen fue descrito como un nuevo espécimen del género Ingenia, conocida como Ingenia sp., de especie incierta, por el paleontólogo chino Lü Junchang y sus colegas en 2002, y solía resaltar las similitudes entre los oviraptorosaurianos y las aves.[1][2][4]
En 2004, Lü y sus colegas determinaron que el esqueleto pertenecía a un taxón nuevo y distinto, y lo convirtieron en el espécimen holotipo de Nemegtia barsboldi. El nombre del género se refiere a la cuenca de Nemegt y el nombre específico honra al paleontólogo mongol Rinchen Barsbold , el líder del equipo que encontró el espécimen.[1] En 2005, los descriptores descubrieron, después de ser notificado por un biólogo, que el nombre Nemegtia ya se había utilizado para un género de Ostracoda, de la misma formación en 1978 y por lo tanto estaba preocupado . Propusieron en cambio el nuevo nombre de género Nemegtomaia, " maia " significa "buena madre" en griego, y el nombre completo significa "buena madre del Nemegt", haciendo referencia al descubrimiento reciente de que los oviraptóridos criaron huevos en lugar de robarlos, aunque no hay rastro de un nido o todavía se habían encontrado huevos asociados con la propia Nemegtomaia.[6][7] El primer miembro conocido de la familia de los oviraptóridos fue encontrado con un nido de huevos que originalmente se creía que pertenecía al género ceratopsiano Protoceratops y por lo tanto fue nombrado Oviraptor en 1924, este nombre significa "ladrón de huevo". En la década de 1990, se descubrieron más especímenes de oviraptóridos asociados con nidos y huevos, en los que se encontraron embriones de oviraptóridos, lo que demuestra que los huevos pertenecían a los propios oviraptóridos.[8][9][10] Ingenia fue renombrada de manera similar como Ajancingenia en 2013, ya que el nombre del género anterior estaba ocupado por un Nematoda.[11]
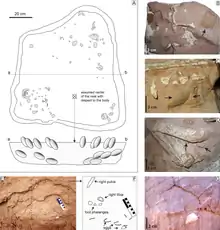
En 2007 , la expedición "Dinosaurios del Gobi" encontró dos nuevos especímenes de Nemegtomaia y fueron descritos por el paleontólogo italiano Federico Fanti y sus colegas en 2013. El primer espécimen, MPC-D 107/15, fue encontrado por Fanti, quien lo apodo "María", en la Formación Baruun Goyot y consiste en un nido con el presunto padre en la parte superior. Como el cuarto género de oviraptórido encontrado en la parte superior de un nido, después de Oviraptor , Citipati y Machairasaurus cf., Nemegtomaia por lo tanto, había recibido un nombre de género que se refería a esta característica antes de que fuera encontrada asociada con los huevos. El espécimen fue excavado desde un acantilado vertical en "circunstancias difíciles", incluyendo fuertes lluvias y colapso de bloques de arenisca.[2][7]
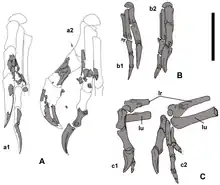
El esqueleto encotrado en el nido preserva partes del cráneo, ambas escápulas, el brazo y la mano izquierdos, el húmero derecho, los huesos púbicos, los isquiones, los fémures, las tibias, los peroné y las porciones inferiores de ambos pies. Este espécimen se encontró a menos de 500 metros del holotipo y era del mismo tamaño; fue asignado a Nemegtomaia debido a sus características anatómicas similares y su proximidad geográfica. Se recolectó en un solo bloque para preservar la relación espacial de los huesos y los huevos. El segundo espécimen, MPC-D 107/16, fue encontrado por el paleontólogo estadounidense Nicholas R. Longrich en la Formación Nemegt, y consta de las manos, un cúbito y radio izquierdos parciales, costillas, una pelvis parcial y ambos fémures. Este espécimen era 35% más pequeño que los otros, y fue asignado a Nemegtomaia debido a que sus manos tenían las mismas características que las del espécimen MPC-D 107/15. Es posible que las manos hayan pertenecido a un individuo diferente, ya que no se encontraron articuladas con el resto del esqueleto, se conocen otros oviraptoridos de canteras con múltiples esqueletos, pero esto no puede confirmarse.[2]
Clasificación
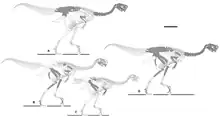
Aunque Nemegtomaia no posee ninguna característica única que lo distinga de otros oviraptóridos, autapomorfias, la combinación de una cresta, un primer dedo agrandado y una gran cantidad de vértebras sacras, ocho, es exclusiva de este taxón.[12] En su análisis filogenético de 2004 , Lü y sus colegas clasificaron a Nemegtomaia como un oviraptorosauriano derivado o "avanzado" y descubrieron que está más estrechamente relacionado con el género Citipati.[1] En 2010, Longrich y sus colegas determinaron que Nemegtomaia pertenecía a la familia Oviraptoridae, como parte de la subfamilia Ingeniinae, por lo que es el único miembro del último grupo con una cresta prominente. Los miembros de la otra subfamilia reconocida, Oviraptorinae, poseen crestas. Los miembros de esta subfamilia se distinguían por un tamaño más pequeño, extremidades anteriores cortas y robustas con garras débilmente curvadas, el número de vértebras en elsynsacrum, así como ciertas características de los pies y la pelvis. Longrich y sus colegas sugirieron que la presencia de una cresta en Nemegtomaia hace posible que esta característica evolucione o desaparezca varias veces entre los oviraptóridos, o que el animal no haya sido parte de Ingeniinae.[2][13] En 2010, el paleontólogo estadounidense Gregory S. Paul sugirió que los oviraptóridos sin cresta eran juveniles o hembras de especies con crestas, y que el número de géneros en el grupo era exagerado. Enumeró Nemegtomaia como " Citipati (=Nemegtomaia) barsboldi", considerándolo muy similar a ese género, pero en 2016 lo enumeró como "Conchoraptor (=Nemegtomaia) barsboldi".[3][14]
En 2012, Fanti y sus colegas también encontraron que Nemegtomaia era parte de Ingeniinae como miembro derivado, más cercano a Heyuannia, debido a las proporciones de las manos de los dos nuevos especímenes, relativamente cortas con un dedo índice robusto. Afirmaron que, aunque la presencia de crestas generalmente se asocia con Oviraptorinae en lugar de Ingeniinae, la característica puede estar correlacionada con el tamaño y la madurez. Señalaron que los huesos nasales y frontales del Ingeniine Conchoraptor eran neumáticos y podrían haber crecido hasta convertirse en una cresta a medida que el animal maduró, aunque todos los esqueletos conocidos de ese género son del mismo tamaño pequeño y una muestra parece haber sido completamente crecido.[2]
Filogenia
El nombre de la subfamilia Ingeniinae ha sido reemplazado por el nombre Heyuanninae, ya que Ingenia estaba ocupada.[15] El siguiente cladograma muestra la ubicación de Nemegtomaia dentro de Oviraptoridae, según Fanti et al., 2012.[2]

| Oviraptoridae |
| |||||||||||||||||||||||||||||||||||||||||||||
Tafonomía
El espécimen MPC-D 107/15 ha proporcionado mucha información sobre los procesos tafonómicos, cambios durante la descomposición y fosilización en la Formación Baruun Goyot. El espécimen se conserva en facies que se cree que se depositaron a través de una tormenta de arena o un cambio de dunas. No parece haber sido transportado después de la muerte, pero el cuerpo parece haberse desplazado ligeramente hacia la derecha, lo que indica que el sedimento en el que se depositó vino hacia él desde su lado izquierdo. El cuello está curvado hacia la izquierda, la mano izquierda está doblada hacia atrás y las piernas están dobladas. La columna vertebral, el cuello y las caderas se deterioraron durante el entierro, y se cree que gran parte del daño al esqueleto fue causado por la actividad de los invertebrados.[2]
Las perforaciones en los huesos, madrigueras y sedimentos reelaborados, tal vez causados por la construcción de cámaras pupales en el espécimen indican que fue capturado por colonias de escarabajos Dermestidae y quizás otros insectos carroñeros. Hay muchos rastros de alimentación en las articulaciones del esqueleto, y casi todas las superficies donde los huesos articulados han sido borrados. También hay túneles en el nido debajo del cuello y el cráneo, y no se han encontrado restos de huevos en partes con tales rastros. Los escarabajos modernos se alimentan principalmente de tejido muscular pero no de materiales húmedos y su actividad se previene mediante un entierro rápido. Por lo tanto, se cree que la muestra MPC-D 107/15 fue enterrado parcialmente al principio, con la parte superior expuesta lo suficiente como para desarrollar una colonia de escarabajos de la piel. Algunos daños en el esqueleto, especialmente en la columna vertebral, también pueden haber sido causados por el accionarn de pequeños mamíferos.[2]
Paleobiología
Evolución
El clado Oviraptorosauria se considera generalmente como un grupo de dinosaurios terópodos no aviarios, y su similitud con las aves se ha observado a menudo. Históricamente, los fósiles de oviraptorosaurios de la familia Caenagnathidae se han confundido con los de las aves y algunos investigadores han llegado a considerar que los oviraptorosaurios en general están más estrechamente relacionados con las aves que con otros dinosaurios no aviarios. En 2002, Lü y sus colegas utilizaron la entonces Nemegtomaia sin nombre para mostrar similitudes entre las aves y los oviraptorosaurios, y descubrieron que este último grupo estaba más cerca de las aves que de los dinosaurios similares a las aves, como los dromaeosaurios. Por lo tanto, concluyeron que los oviraptorosaurios eran aves no voladoras en lugar de dinosaurios no aviarios, y señalaron que el límite entre las aves y los dinosaurios se hacía cada vez más difícil de delinear.[4] Otros investigadores han encontrado en cambio que los dromaeosaurios y troodontidos están más estrechamente relacionados con las aves, formando juntos el grupo Paraves, los oviraptorosaurianos, los terizinosaurianos y los alvarezsaurios están fuera de este grupo. El grupo más amplio que incluye oviraptorosaurianos y Paraves se llama Pennaraptora y este grupo se define por la presencia de plumas penaceas.[16]
Los oviraptorosaurianos se conocen de Asia, donde pueden haberse originado y de América del Norte y se conocen principalmente de depósitos que datan de las edades de Campaniense y Maastrichtiense del período Cretácico superior. El grupo incluye miembros pequeños a grandes, y se caracterizan por sus cráneos y picos cortos, dedos alargados y colas cortas. Los miembros basales o "primitivos" tenían dientes, que desaparecieron en los miembros derivados del grupo, aquellos dentro de la superfamilia Caenagnathoidea, que incluye a Oviraptoridae. Eran al menos parcialmente herbívoros y criaban sus nidos en una postura de pájaro. Aunque se cree que todos tienen que estar emplumados, parecen no haber volado.Las crestas craneales parecen haber evolucionado de manera convergente en diferentes linajes dentro del grupo.[16] La familia Oviraptoridae, a la que pertenece Nemegtomaia, consistía en miembros generalmente pequeños, y se conoce exclusivamente del Cretácico Superior de Asia, con la mayoría de los géneros descubiertos en el desierto de Gobi de Mongolia y China.[13] Incluyendo Nemegtomaia, se han descubierto al menos nueve géneros de oviraptóridos en un área geográfica relativamente pequeña en el desierto de Gobi.[12][17]
Reproduction

El espécimen MPC-D 107/15 se encontró asociado con un nido con huevos, sus pies estaban colocados en el centro de lo que probablemente era un anillo de huevos, con los brazos cruzados sobre la parte superior de los huevos a cada lado del cuerpo, una postura similar a la que se ve en otros fósiles de oviraptóridos. La parte recolectada del nido mide aproximadamente 90 centímetros de ancho y 100 centímetros de largo. El esqueleto ocupa los 25 centímetros superiores del bloque, mientras que los 20 centímetros restantes están ocupados por huevos y cáscaras rotas. No hay evidencia de material vegetal en el nido, pero hay fragmentos de huesos indeterminados. El nido no conserva ningún huevo o embrión completo, lo que impide determinar el tamaño, la forma, el número y la disposición de los huevos en el nido. Es probable que originalmente hubiera dos capas de huevos debajo del cuerpo, y no parece haber huevos en el centro del nido. La mayoría de los huevos, se han identificado siete huevos distintos y los fragmentos de huevo se recuperaron en la capa inferior del nido o debajo del cráneo, el cuello y las extremidades de la muestra y los huesos descansaban directamente sobre los huevos o estaban dentro de 5 milímetros de sus superficies. El hecho de que el esqueleto se haya colocado directamente encima muestra que el nido no estaba completamente cubierto de arena. Aunque la colocación de los huevos no sugiere una disposición específica en el nido, la mayoría de los otros nidos de oviraptóridos muestran que los huevos se organizaron en pares en hasta tres niveles de círculos concéntricos. Los huevos de MPC-D 107/15 fueron, por lo tanto, muy probablemente desplazados durante el entierro, o por factores externos, como fuertes vientos, tormentas de arena o depredadores. Esto también respalda la idea de que la capa superior de los huevos no fue enterrada, ya que los huevos completamente enterrados habrían sido menos propensos a ser transportados por factores externos.[2]

Los huevos oviraptóridos parecen haber medido 17 centímetros de largo en promedio, y se cree que los huevos más completos encontrados con MPC-D 107/15 tenían 5 a 6 centímetros de ancho y 14 a 16 centímetros de largo cuando estaban intactos. Los huevos son casi idénticos a algunos que se han encontrado previamente en Mongolia y por lo tanto, han sido asignados a la oofamilia, familia de taxones de huevos Elongatoolithidae. Las cáscaras de huevo son relativamente delgadas, entre 1 y 1,2 milímetros, y su superficie externa está cubierta por crestas y nodos que se elevan aproximadamente 0.3 milímetros por encima de la cáscara. La microestructura de las cáscaras de huevo no se pudo estudiar adecuadamente, ya que la calcita ha sido muy alterada y recristalizada.[2]
El espécimen de anidación se encontró en un área estratigráfica que indica que los oviraptóridos preferían anidar cerca de arroyos que proporcionaban sustrato blando y arenoso y alimento en entornos que de otro modo serían xéricos, que recibían una pequeña cantidad de humedad. Se han encontrado muchos oviraptóridos en posiciones de cría, lo que indica que pueden haber criado durante períodos relativamente largos, similar a las aves modernas como el avestruz , el emú y el ratonero de pecho negro, que crían durante más de 40 días con un suministro limitado de sustento. La anidación en ambientes desérticos puede ser perjudicial para los adultos que permanecen en el nido durante gran parte del día, y para los huevos y los pichones, debido al estrés por calor. Por lo tanto, la elección del área de anidación puede haber sido un mecanismo para una incubación exitosa en calor extremo. También se ha sugerido que la evolución de las plumas de la cola en los oviraptorosaurios fue una adaptación para sombrear y proteger los huevos en sus nidos. Que el segundo dedo de los oviraptóridos de Heyuanninae se redujo de tamaño en comparación con el robusto primer dedo puede explicarse por un cambio en la función; puede estar relacionado con la presencia de plumas de ala larga que estaban unidos al segundo dedo. Estas plumas de ala probablemente se usaron para proteger los huevos durante la anidación. Cuando el segundo dedo comenzó a funcionar como un soporte de plumas, su capacidad de agarre se redujo, y esta función fue asumida por el primer dedo, que por lo tanto se hizo más robusto. El tercer dedo también se redujo de tamaño, probablemente porque se colocó detrás de las plumas de las alas de una manera donde no sería efectivo para agarrar.[2]

En 2018, el paleontólogo taiwanés Tzu-Ruei Yang y sus colegas identificaron capas de cutículas en cáscaras de huevo de dinosaurios maniraptores, incluidos los oviraptóridos. En las aves modernas, tales capas que consisten principalmente en lípidos e hidroxiapatita sirven para proteger los huevos de la deshidratación y la invasión de microbios. Los investigadores sugirieron que los huevos recubiertos con cutícula habrían sido un rasgo adaptado para mejorar su éxito reproductivo en los entornos variables donde anidaban Nemegtomaia y otros oviraptóridos.[18]
Varios estudios han sugerido que varios animales pondrían huevos en un solo nido y los organizarían para que pudieran ser protegidos por una persona, posiblemente un macho.[2] En 2010, el paleontólogo estadounidense David J. Varricchio y sus colegas encontraron que la relativamente grande del nido, el tamaño del de oviraptóridos y troodóntidos es más similar a las de los modernos arcosaurios, aves y los cocodrilos, los parientes vivos más cercanos de los dinosaurios, que la práctica polígama de apareamiento y cuidado parental masculino extenso, como se ve en paleognatos como avestruces y emus. Este sistema reproductivo es anterior al origen de las aves y por lo tanto, sería la condición ancestral para las aves modernas, con el cuidado biparental, donde ambos padres participan, como un desarrollo posterior. [19] Se sabe que muchos oviraptorosaurios tenían pigóstilo en el extremo de sus colas, lo que sugiere la presencia de abanicos de plumas. El paleontólogo estadounidense W. Scott Persons y sus colegas sugirieron en 2013 que estos podrían haber sido utilizados para la comunicación intraespecífica, como los rituales de cortejo.[19]
Dieta

La dieta de los oviraptóridos se ha interpretado de varias maneras desde que se pensó erróneamente que Oviraptor era un depredador de los huevos. Se ha sugerido que los oviraptorosaurianos en su conjunto eran herbívoros, que están respaldados por los gastrolitos, cálculos estomacales, que se encuentran en Caudipteryx , y las facetas de desgaste en los dientes de Incisivosaurus. En 2010 Longrich y colegas encontraron que las mandíbulas oviraptíridos tenían características similares a las observadas en herbívoros tetrápodos, animales de cuatro con extremidades, especialmente los de dicinodontes, un grupo extinto de sinápsido protomamíferos. Los oviraptóridos y los dicinodontos comparten características tales como mandíbulas cortas, profundas y sin dientes, sinfisias dentadas alargadas, fenestra mandibular alargada y una barra que se proyecta hacia abajo en el paladar. Los animales modernos con mandíbulas que se parecen a los de los oviraptóridos incluyen loros y tortugas. El último grupo también tiene proyecciones similares a dientes en sus premaxilares. Longrich y sus colegas concluyeron que debido a las similitudes entre los oviraptóridos y los animales herbívoros, la mayor parte de su dieta probablemente se habría formado por materia vegetal. Los oviraptóridos se encuentran a altas frecuencias en las formaciones de las que se conocen, similar al patrón visto en los dinosaurios que se sabe que son herbívoros. Estos animales eran más abundantes que los dinosaurios carnívoros, ya que había más energía disponible en su nivel trófico inferior en la cadena alimenticia. Las mandíbulas de los oviraptóridos pueden haberse especializado para procesar alimentos, como la vegetación xerofítica, adaptada para ambientes con poca agua, que habría crecido en su entorno, pero esto no es posible demostrarlo, ya que se sabe poco sobre la flora del área en el momento.[13] Un estudio de 2013 realizado por Lü y sus colegas encontró que los oviraptóridos parecen haber conservado sus proporciones de extremidades posteriores en toda la ontogenia, que también es un patrón que se observa principalmente en animales herbívoros.[20] En 2017, el paleontólogo canadiense Gregory F. Funston y sus colegas sugirieron que las mandíbulas de los oviraptóridos en forma de loro pueden indicar una dieta frugívoro que incorpora nueces y semillas.[12]
En 1977, Barsbold sugirió que los oviraptóridos se alimentaban de moluscos, pero Longrich y sus colegas rechazaron la idea de que practicaran la trituración de conchas, ya que estos animales tienden a tener dientes con amplias superficies de trituración. En cambio, la forma de los huesos dentarios en las mandíbulas inferiores de los oviraptóridos sugiere que tenían un pico afilado que se usaba para cortar alimentos duros, no para romper alimentos duros como bivalvos o huevos. El estante sinfisario en la parte frontal del dentario puede haber dado cierta capacidad de aplastamiento, pero como era un área relativamente pequeña, probablemente no era la función principal de las mandíbulas. El hecho de que la mayoría de los oviraptoridos se han encontrado en sedimentos que se interpretan como ambientes xéricos y áridos o semiáridos también argumenta en contra de que hayan sido comedores especializados de mariscos y huevos, ya que es poco probable que haya habido suficientes elementos en tales condiciones para soportarlos.[13]
Longrich y sus colegas señalaron que las extremidades anteriores robustas y el agrandamiento de un solo dedo en los oviraptóridos heyuaninos son similares a los que se ven en los animales modernos que comen hormigas y termitas, como los osos hormigueros y los pangolines, pero la morfología de las mandíbulas heyuaninas no respalda que sean insectívoros. Los investigadores encontraron que la función de las extremidades anteriores de heyuaninos no estaba clara, pero sugirieron que podrían haberse usado para rascarse, rasgarse o excavar, aunque no para capturar presas.[13]
En 2004, Lü y sus colegas propusieron que la articulación entre los huesos cuadrado y cuadratoyugal en el cráneo de Nemegtomaia sugirió que estos huesos eran móviles entre sí , lo que habría afectado el funcionamiento de las mandíbulas. En 2015, el paleontólogo belga Christophe Hendrickx y sus colegas encontraron que era poco probable que Nemegtomaia y otros oviraptóridos tuvieran cinesis parecida a un pájaro en sus cráneos, debido a que el hueso cuadrado está inmóvil.[21]
Paleoecología
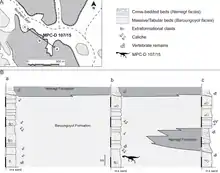
Nemegtomaia es conocido de las formaciones de Nemegt y Baruun Goyot, que datan de las edades de Campaniense superior a Maastrichtiense inferior del Cretácico tardío, hace unos 70 millones de años. Si bien este taxón solo se conoce en la localidad de Nemegt, pueden pertenecer restos oviraptóridos no identificados de otras localidades. El macizo de Nemegt tiene numerosos cañones o gargantas, hasta 45 metros de profundidad, que tienen algunas de las mejores exposiciones de estas formaciones. Se cree que las facies de rocas de la Formación Nemegt representan un ambiente húmedo y fluvial, asociado con ríos y arroyos, mientras que las de la Formación Baruun Goyot representan un ambiente árido o semiárido, con camas eólicas, afectadas por el viento. Históricamente se pensó que estas dos formaciones con sus diversos fósiles representaban períodos de tiempo secuenciales con diferentes entornos, pero en 2009 el paleontólogo canadiense David A. Eberth y sus colegas descubrieron que había una superposición parcial en la transición entre ellos. Las dos formaciones se intercalan a través de un intervalo estratigráfico que es de unos 25 metros de espesor, lo que sugiere que los entornos fluviales y eólicos coexistieron cuando el área se sedimentó.[2][12][22][23]

El entorno de la Formación Nemegt se ha comparado con el Delta del Okavango de Botsuana actual.[24] Los hábitats en y alrededor de los ríos Nemegt proporcionaron un hogar para una amplia gama de organismos. Los animales acuáticos incluyen moluscos, peces, tortugas y el crocodilomorfo Shamosuchus. Se han encontrado fósiles de mamíferos, como los multituberculados y se conocen aves como Gurilynia, Judinornis y Teviornis. Los dinosaurios herbívoros de la Formación Nemegt incluyen anquilosáuridos como Tarchia , el paquicefalosauriano Prenocephale, hadrosáuridos como Saurolophus y Barsboldia y sauropodos como Nemegtosaurus y Opisthocoelicaudia. Otros terópodos como los tiranosáuridos Tarbosaurus, Alioramus y Bagaraatan, trodóntidoss como Borogovia, Tochisaurus y Saurornithoides, tericinosáuridos Therizinosaurus y ornitomimosaurianos como Deinocheirus, Anserimimus y Gallimimus.[25][26][27]
Otros géneros de oviraptorosaurianos conocidos de la Formación Nemegt incluyen el basal Avimimus, los oviraptóridos Rinchenia, Nomingia, Conchoraptor y Ajancingenia , y el cenagnátido Elmisaurus. A pesar de la gran cantidad de taxones de oviraptóridos en estas formaciones, el Nemegt tiene la mayor diversidad conocida de todas, ninguno de ellos estaba estrechamente relacionado. La Formación Nemegt es única en tener oviraptóridos y caenagnátidos y en 1993 el paleontólogo canadiense Phillip J. Curriey sus colegas sugirieron que esta diversidad se debía a que los dos grupos preferían diferentes ambientes en el área. En 2016, el paleontólogo japonés Takanobu Tsuihiji y sus colegas sugirieron que los oviraptóridos podrían haber preferido ambientes más secos, mientras que los cenagnátidas preferían los ambientes fluviales, según el tipo de formaciones en las que se han encontrado.[2][28] Funston y sus colegas sugirieron que se encontraron oviraptóridos tanto en ambientes xéricos como mesímicos,pero fueron más abundantes en los primeros, mientras que los otros grupos de oviraptorosaurios evitaron los ambientes xéricos, y que la coexistencia de las familias puede explicarse mediante la división de nicho en la dieta. El ambiente de la Formación Nemegt puede haber actuado como un oasis y de ese modo atrajo oviraptóridos.[12]
Referencias
- Lü, J.; Tomida, Y.; Azuma, Y.; Dong, Z.; Lee, Y.-N. (2004). «New oviraptorid dinosaur (Dinosauria: Oviraptorosauria) from the Nemegt Formation of southwestern Mongolia». Bulletin of the National Science Museum, Tokyo, Series C 30: 95-130.
- Fanti, F.; Currie, P. J.; Badamgarav, D.; Lalueza-Fox, C. (2012). «New specimens of Nemegtomaia from the Baruungoyot and Nemegt Formations (Late Cretaceous) of Mongolia». PLoS ONE 7 (2): e31330. Bibcode:2012PLoSO...731330F. PMC 3275628. PMID 22347465. doi:10.1371/journal.pone.0031330.
- Paul, G. S. (2010). The Princeton Field Guide to Dinosaurs. Princeton University Press. pp. 152, 154. ISBN 978-0-691-13720-9. (requiere registro).
- Lü, J., Dong, Z., Azuma, Y., Barsbold, R. & Tomida, Y. (2002). "Oviraptorosaurs compared to birds." In Zhou, Z. & Zhang, F. (eds). Proceedings of the 5th Symposium of the Society of Avian Paleontology and Evolution. Beijing: Science Press. pp. 175–189.
- Lü, J.; Yi, L.; Zhong, H.; Wei, X.; Dodson, P. (2013). «A new oviraptorosaur (Dinosauria: Oviraptorosauria) from the Late Cretaceous of Southern China and its paleoecological implications». PLoS ONE 8 (11): e80557. Bibcode:2013PLoSO...880557L. PMC 3842309. PMID 24312233. doi:10.1371/journal.pone.0080557.
- Lü, J.; Tomida, Y.; Azuma, Y.; Dong, Z.; Lee, Y.-N. (2005). «Nemegtomaia gen. nov., a replacement name for the oviraptorosaurian dinosaur Nemegtia Lü et al. 2004, a preoccupied name». Bulletin of the National Science Museum, Tokyo, Series C 31: 51.
- Arbour, V.M. (2012). «Gobi Desert Diaries: Nemegtomaia Edition». pseudoplocephalus.blogspot.com. Consultado el 10 de enero de 2017.
- Norell, M.A.; Clark, J. M.; Chiappe, L.M.; Dashzeveg, D. (1995). «A nesting dinosaur». Nature 378 (6559): 774-776. Bibcode:1995Natur.378..774N. doi:10.1038/378774a0.
- Dong, Z.-M.; Currie, P.J. (1996). «On the discovery of an oviraptorid skeleton on a nest of eggs at Bayan Mandahu, Inner Mongolia, People's Republic of China». Canadian Journal of Earth Sciences 33 (4): 631-636. Bibcode:1996CaJES..33..631D. doi:10.1139/e96-046.
- Clark, J. M.; Norell, M.; Chiappe, L. M. (1999). «An oviraptorid skeleton from the late Cretaceous of Ukhaa Tolgod, Mongolia, preserved in an avianlike brooding position over an oviraptorid nest». American Museum Novitates (3265): 1-35. hdl:2246/3102.
- Easter, J. (2013). «A new name for the oviraptorid dinosaur "Ingenia" yanshini (Barsbold, 1981; preoccupied by Gerlach, 1957)». Zootaxa 3737 (2): 184-190. PMID 25112747. doi:10.11646/zootaxa.3737.2.6.
- Funston, G.F.; Mendonca, S.E.; Currie, P.J.; Barsbold, R. (2017). «Oviraptorosaur anatomy, diversity and ecology in the Nemegt Basin». Palaeogeography, Palaeoclimatology, Palaeoecology 494: 101-120. Bibcode:2018PPP...494..101F. doi:10.1016/j.palaeo.2017.10.023.
- Longrich, N.R.; Currie, P.J.; Zhi-Ming, D. (2010). «A new oviraptorid (Dinosauria: Theropoda) from the Upper Cretaceous of Bayan Mandahu, Inner Mongolia». Palaeontology 53 (5): 945-960. doi:10.1111/j.1475-4983.2010.00968.x.
- Paul, G. S. (2016). The Princeton Field Guide to Dinosaurs. Princeton University Press. p. 181. ISBN 978-0-691-16766-4.
- Yun, C.-G. (2019). «Heyuanninae clade nov., a replacement name for the oviraptorid subfamily “Ingeniinae” Barsbold, 1981». Zootaxa 4671 (2): 295-296. doi:10.11646/zootaxa.4671.2.11.
- Hendrickx, C.; Hartman, S.A.; Mateus, O. (2015). «An overview on non-avian theropod discoveries and classification». PalArch's Journal of Vertebrate Palaeontology 12 (1): 1-73. Archivado desde el original el 22 de junio de 2018. Consultado el 12 de junio de 2020.
- Wang, S.; Sun, C.; Sullivan, C.; Xu, X. (2013). «A new oviraptorid (Dinosauria: Theropoda) from the Upper Cretaceous of southern China». Zootaxa 3640 (2): 242. PMID 26000415. doi:10.11646/zootaxa.3640.2.7.
- Yang, T.-.; Chen, Y.-H.; Wiemann, J.; Spiering, B.; Sander, P. M. (2018). «Fossil eggshell cuticle elucidates dinosaur nesting ecology». PeerJ 6: e5144. PMC 6037156. PMID 30002976. doi:10.7717/peerj.5144.
- Persons, S. (2013). «Oviraptorosaur tail forms and functions». Acta Palaeontologica Polonica. doi:10.4202/app.2012.0093. Parámetro desconocido
|doi-access=ignorado (ayuda) - Lü, J.; Currie, P.J.; Xu, L.; Zhang, X.; Pu, H.; Jia, S. (2013). «Chicken-sized oviraptorid dinosaurs from central China and their ontogenetic implications». Naturwissenschaften 100 (2): 165-175. Bibcode:2013NW....100..165L. PMID 23314810. doi:10.1007/s00114-012-1007-0.
- Hendrickx, C.; Araújo, R.; Mateus, O. (2015). «The non-avian theropod quadrate I: standardized terminology with an overview of the anatomy and function». PeerJ 3: e1245. PMC 4579021. PMID 26401455. doi:10.7717/peerj.1245.
- Eberth, D. A.; Badamgarav, D.; Currie, P. J. (2009). «The Baruungoyot-Nemegt transition (Upper Cretaceous) at the Nemegt type area, Nemegt Basin, South Central Mongolia». 고생물학회지 25 (1). ISSN 1225-0929.
- Eberth, D.A. (2017). «Stratigraphy and paleoenvironmental evolution of the dinosaur-rich Baruungoyot-Nemegt succession (Upper Cretaceous), Nemegt Basin, southern Mongolia». Palaeogeography, Palaeoclimatology, Palaeoecology 494: 29-50. Bibcode:2018PPP...494...29E. doi:10.1016/j.palaeo.2017.11.018.
- Holtz, T.R. (2014). «Paleontology: Mystery of the horrible hands solved». Nature 515 (7526): 203-205. Bibcode:2014Natur.515..203H. PMID 25337885. doi:10.1038/nature13930.
- Lucas, S.G.; Estep, J.W. (1998). «Vertebrate biostratigraphy and biochronology of the Cretaceous of China». New Mexico Museum of Natural History and Science Bulletin 14: 1-20.
- Watabe, M.; Tsogtbaatar, K.; Suzuki, S.; Saneyoshi, M. (2010). «Geology of dinosaur-fossil-bearing localities (Jurassic and Cretaceous: Mesozoic) in the Gobi Desert: Results of the HMNS-MPC Joint Paleontological Expedition». Hayashibara Museum of Natural Sciences Research Bulletin 3: 11-18.
- Jerzykiewicz, T.; Russell, D.A. (1991). «Late Mesozoic stratigraphy and vertebrates of the Gobi Basin». Cretaceous Research 12 (4): 345-377. doi:10.1016/0195-6671(91)90015-5.
- Tsuihiji, T.; Watabe, M.; Tsogtbaatar, K.; Barsbold, R. (2016). «Dentaries of a caenagnathid (Dinosauria: Theropoda) from the Nemegt Formation of the Gobi Desert in Mongolia». Cretaceous Research 63: 148-153. doi:10.1016/j.cretres.2016.03.007.
| Control de autoridades |
|
|---|
 Datos: Q935296
Datos: Q935296 Multimedia: Nemegtomaia / Q935296
Multimedia: Nemegtomaia / Q935296 Especies: Nemegtomaia
Especies: Nemegtomaia