Opisthocoelicaudia skarzynskii
Opisthocoelicaudia skarzynskii es la única especie conocida del género extinto Opisthocoelicaudia ("cola con excavaciones posteriores") de dinosaurio saurópodo saltasáurido, que vivió a finales del período Cretácico, hace aproximadamente 70 millones de años, en el Maastrichtiense, de lo que hoy es Asia.
| Opisthocoelicaudia skarzynskii | ||
|---|---|---|
| Rango temporal: 70 Ma Cretácico superior | ||
 | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Sauropsida | |
| Superorden: | Dinosauria | |
| Orden: | Saurischia | |
| Suborden: | Sauropodomorpha | |
| Infraorden: | Sauropoda | |
| (sin rango): | Titanosauria | |
| Familia: | Saltasauridae | |
| Subfamilia: | Opisthocoelicaudiinae | |
| Género: |
Opisthocoelicaudia Borsuk-Bialynicka, 1977 | |
| Especie: |
O. skarzynskii Borsuk-Białynicka, 1977 | |
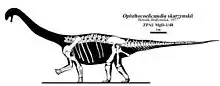
Se basa en un esqueleto bien conservado que carece solo de la cabeza y el cuello fue desenterrado en 1965 por científicos polacos y mongoles, lo que hizo que Opisthocoelicaudia uno de los saurópodos más conocidos del Cretácico tardío. Las marcas de dientes en este esqueleto indican que grandes dinosaurios carnívoros se habían alimentado de la carcasa y posiblemente se habían llevado las partes que ahora faltaban. Hasta la fecha, solo se conocen dos especímenes adicionales, mucho menos completos, incluida una parte de un hombro y una cola fragmentaria. Como saurópodo era relativamente pequeño, Opisthocoelicaudia midió aproximadamente 11,4 a 13 metros de longitud. Al igual que otros saurópodos, se habría caracterizado por una cabeza pequeña sentada sobre un cuello muy largo y un tronco en forma de barril con cuatro patas en forma de columna. El nombre Opisthocoelicaudia significa "cola de la cavidad posterior", aludiendo a la inusual condición opistocoelicas de las vértebras de la cola anterior que eran cóncavas en sus lados posteriores. Esta y otras características esqueléticas llevan a los investigadores a proponer que la Opisthocoelicaudia fue capaz de pararse sobre sus patas traseras.
Nombrado y descrita por la paleontóloga polaca Maria Magdalena Borsuk-Białynicka en 1977, se pensó en un principio que Opisthocoelicaudia era un nuevo miembro de Camarasauridae, pero actualmente se considera un miembro derivado de Titanosauria. Sus relaciones exactas dentro de Titanosauria son polémicas, pero puede haber estado cerca del Alamosaurus norteamericano . Todos los fósiles de Opisthocoelicaudia provienen de la Formación Nemegt. A pesar de ser rico en fósiles de dinosaurios, el único otro saurópodo de esta unidad de roca es Nemegtosaurus, que se conoce por un solo cráneo. Como el cráneo de Opisthocoelicaudia sigue siendo desconocida, varios investigadores han sugerido que Nemegtosaurus y Opisthocoelicaudia pueden representar la misma especie. Las huellas de saurópodos de la Formación Nemegt, que incluyen impresiones de la piel, probablemente pueden referirse a Nemegtosaurus u Opisthocoelicaudia, ya que estos son los únicos saurópodos conocidos de esta formación.
Descripción

Al igual que otros saurópodos, Opisthocoelicaudia tenía una cabeza pequeña en un cuello largo, un cuerpo en forma de barril en cuatro extremidades columnares y una cola larga. Era relativamente pequeño para un saurópodo, el espécimen tipo se estimó en 11,4 a 13 metros desde la cabeza hasta la punta de la cola.[1][2][3] La masa corporal se ha estimado en 8,4 toneladas,[4] 10,5 toneladas,[1] 22 toneladas,[5] 13 toneladas[6] y 25,4 toneladas[7] en estudios separados.

El cráneo y el cuello no se conservan, pero la reconstrucción del ligamento nucal indica la posesión de un cuello de longitud media de aproximadamente 5 metros.[8] Borsuk-Białynicka, en su descripción de 1977, notó la presencia de 11 vértebras dorsales. Gregory Paul, en 2019, argumentó que la parte conocida de la columna vertebral en realidad incluye la última cervical, dejando solo 10 dorsales, el recuento típico en los titanosaurianos.[9] Al igual que en otros titanosaurianos, la espalda era bastante flexible debido a la falta de articulaciones vertebrales accesorias, lasarticulaciones hiposfeno-hypantro,[10] mientras que la región pélvica se fortaleció con una sexta vértebra de cadera adicional.[8] Las vértebras anteriores de la cola eran opistócoelicas, lo que significa que eran convexas en sus lados anteriores y cóncavas en sus lados posteriores, formando articulaciones esféricas.[8] Estas vértebras de la cola opistocoelicas le dan a Opisthocoelicaudia su nombre y sirven para distinguir el género de todos los otros titanosaurianos.[11] Otros titanosaurios generalmente se caracterizaban por vértebras de la cola anterior fuertemente procoelicas, que eran cóncavas en sus lados anteriores y convexas en sus lados posteriores.[12] Otra característica única se puede encontrar en las vértebras de la espalda, que muestran procesos espinosos bifurcados, lo que resulta en una doble fila de proyecciones óseas a lo largo de la parte superior de la columna vertebral.[13] Si bien es único en los titanosaurianos, esta característica se puede encontrar en varios otros saurópodos no relacionados, incluidos Diplodocus y Euhelopus, donde evolucionó de forma independiente.[14]

Al igual que en las caderas de otros titanosaurianos, el isquion era relativamente corto y medía solo dos tercios de la longitud del pubis. Los huesos del isquion izquierdo y derecho, así como los huesos del pubis izquierdo y derecho se osificaron entre sí en la mayor parte de su longitud, cerrando la brecha que en otros saurópodos normalmente está presente entre estos huesos.[11] Las extremidades eran proporcionalmente cortas, como se ve en otros titanosaurianos.[15] Las extremidades anteriores midieron 1,87 metros de altura en la muestra casi completa, aproximadamente dos tercios de la longitud de las extremidades posteriores, que se reconstruyeron a 2,46 metros de altura.[8] Al igual que en otros titanosaurianos, las extremidades se extendieron ligeramente hacia afuera en lugar de pararse verticalmente debajo del cuerpo,[16] mientras que las extremidades anteriores eran más flexibles y móviles en comparación con otros saurópodos.[15]
La mano estaba compuesta simplemente por las cinco metacarpiales, que estaban orientadas verticalmente y dispuestas en semicírculo. Faltaban huesos del carpo, como en otros titanosaurianos.[8][17] Los huesos de los dedos y las garras también estaban completamente ausentes, en la mayoría de los otros titanosaurianos, estos huesos todavía estaban presentes, aunque de tamaño extremadamente reducido. En el pie, el hueso astrágalo se redujo fuertemente como en otros titanosaurianos, mientras que el calcáneo probablemente estaba completamente ausente en Opisthocoelicaudia.[8] En contraste con la mano, el pie mostraba dígitos y garras bien desarrollados. La fórmula falangeana, que indica el número de falanges, que comienzan con el dígito más interno, es 2-2-2-1-0. Raramente se encuentran esqueletos de pie de titanosaurianos y además de Opisthocoelicaudia, solo se conocen ejemplos completamente conservados de Epachthosaurus y el titanosauriano de La Invernada, cuyas fórmulas falangeales son 2-2-3-2-0 y 2-2-2-2-0, respectivamente. De estos tres titanosaurianos, Opisthocoelicaudia fue la más derivada al mostrar la menor cantidad de falanges, lo que indica una reducción progresiva en el recuento de falanges durante la evolución de los titanosaurianos.[18] La garra del primer dígito no era más grande que el segundo dígito, como en otros saurópodos, pero era aproximadamente igual en tamaño.[19]

Se han encontrado osteodermos, placas óseas formadas en la piel, con al menos 10 géneros de titanosaurianos. La falta de osteodermos en el esqueleto casi completo de Opisthocoelicaudia podría indicar que están ausentes en este género.[20] Sin embargo, se descubrió que el Alamosaurus estrechamente relacionado tenía osteodermos casi un siglo después de su descubrimiento, además de varios otros titanosaurianos estrechamente relacionados, lo que aumenta la posibilidad de que Opisthocoelicaudia también los tuviera.[21]
Descubrimiento e investigación
El espécimen tipo fue descubierto entre el 10 y el 23 de junio de 1965, durante una expedición paleontológica polaco-mongol dirigida por la paleontóloga polaca Zofia Kielan-Jaworowska.[22][23] La mayor de una serie de expediciones llevadas a cabo en 1963-1971, esta expedición involucró a 21 miembros, que a veces fueron apoyados por trabajadores mongoles adicionales contratados. El sitio de descubrimiento se encuentra en la provincia de Ömnögovi, en el sur de Mongolia, en el área de Altan Uul, que expone unos 100 km² de tierras baldías. Los sedimentos expuestos en Altan Uul pertenecen a la Formación Nemegt, la más joven de las tres formaciones geológicas de la Cuenca Nemegt. Opisthocoelicaudia fue el primero de varios descubrimientos importantes de dinosaurios realizados por la expedición de 1965. Los otros hallazgos, realizados en diferentes localidades, incluyen varios esqueletos del tiranosáurido Tarbosaurus, así como los especímenes tipo del ornitomimosauriano gigante Deinocheirus, el saurópodo Nemegtosaurus y el paquicefalosáurido Homalocephale.[23]

En el quinto día de trabajo de campo, Ryszard Gradziński, el geólogo de la expedición, encontró una concreción de huesos bien conservados que prometían pertenecer a un esqueleto bastante completo. La excavación comenzó al día siguiente, revelando un esqueleto casi completo que carece solo de la cabeza y el cuello. Hasta hoy, este espécimen sigue siendo, con mucho, el hallazgo más completo de este dinosaurio. El transporte del espécimen fuera del terreno accidentado causó importantes problemas técnicos. Los bloques de piedra que contenían los fósiles tuvieron que moverse unos 580 metros en trineos de metal improvisados, hechos de bidones de gasolina antes de poder cargarlos en camiones. Porque el esqueleto estaba incrustado en capas de arenisca muy dura, varios bloques pesaron más de una tonelada. El 9 de julio, el embalaje del esqueleto en 35 cajas comenzó para su transporte a Dalanzadgad, juntas, las cajas pesaban unas 12 toneladas.[22][23]
El espécimen tipo pertenecía a un individuo anciano.[8] Su tafonomía es inusual ya que se encontró acostada sobre su espalda; en contraste, la mayoría de los otros esqueletos de dinosaurios casi completos de la Formación Nemegt generalmente se encuentran acostados de lado.[24] El espécimen se encontró encerrado en una cama cruzada de arenisca depositada por un río. La mayoría de las vértebras descubiertas todavía estaban conectadas entre sí, formando una serie continua que constaba de 8 vértebras dorsales, 6 sacras y 34 caudales. Se encontraron 3 vértebras adicionales aisladas de la serie y pueden pertenecer al área de transición entre la espalda y el cuello. Las partes restantes del esqueleto se desplazaron ligeramente fuera de su posición anatómica original. Tanto la extremidad izquierda como los huesos de las costillas se encontraron en el lado derecho del cuerpo, mientras que, por el contrario, se encontraron la extremidad derecha y los huesos de las costillas en el lado izquierdo. Se han identificado marcas de mordida en el esqueleto, particularmente en la pelvis y el hueso del muslo, lo que muestra que los depredadores se habían alimentado de la carcasa. Faltan el cráneo y el cuello, lo que indica que los carnívoros podrían haberse llevado estas partes del cuerpo.[8] La integridad de los restos indica que el individuo había muerto cerca del sitio del descubrimiento.[24] Un evento de inundación podría haber transportado la carcasa a una corta distancia y posteriormente cubrirla con sedimento, incluso antes de que el tejido blando se hubiera descompuesto por completo.[8]

En 1977, la paleontóloga polaca Maria Magdalena Borsuk-Białynicka publicó su descripción completa del esqueleto y nombró a Opisthocoelicaudia skarzynskii como un nuevo género y especie. El nombre del género, que insinúa la condición inusual de opistocoelica de las vértebras de la cola, significa "cola de la cavidad posterior". Se deriva del griego ὄπισθεν, opisthen, "detrás, atrás", κοῖλος, koilos, "hueco" y del latín cauda, cola.[25] El nombre específico honra a Wojciech Skarżyński, la persona que preparó el espécimen tipo.[8] Opisthocoelicaudia fue el tercer saurópodo de Asia conocido por un esqueleto postcraneal, después de Euhelopus y Mamenchisaurus.[8] Después de su descubrimiento, el esqueleto holotipo se convirtió en parte de la colección del Instituto de Paleobiología en Varsovia, pero luego fue devuelto a su país de origen, ahora alojado en el Instituto de Geología de la Academia de Ciencias de Mongolia en Ulán Bator. bajo el número de catálogo MPC-D100/404.[26][19] Además del espécimen tipo, Borsuk-Białynicka describió un omóplato y un coracoide, ZPAL MgD-I/25c de la misma localidad. Estos huesos aún no estaban fusionados entre sí, lo que indica un individuo juvenil.[8]
Para 2017, los fósiles de saurópodos se habían recuperado de un total de 32 localidades dentro de la Formación Nemegt y posiblemente pertenezcan a Opisthocoelicaudia o Nemegtosaurus. Al menos dos hallazgos de la localidad de Nemegt, una cola fragmentaria, MPD 100/406 y un par de garras, muestran características de diagnóstico para Opisthocoelicaudia y se pueden derivar a este último.[27][19] Los equipos de campo liderados por Philip Currie intentaron reubicar la cantera holotipo Opisthocoelicaudia en 2006 y 2008, pero tuvieron éxito solo en 2009 gracias a los datos adicionales proporcionados por Gradziński.[19] Aunque no fue posible la prospección de material óseo adicional ya que la cantera había sido llenada por arena arrastrada por el viento, se podía determinar que la cantera caía dentro de la porción inferior de la Formación Nemegt.[19]
Clasificación

Originalmente, Opisthocoelicaudia fue clasificada como miembro de la familia Camarasauridae, junto con Camarasaurus y Euhelopus. Esta clasificación se basó en varias características compartidas del esqueleto, sobre todo las espinas neurales bifurcadas de las vértebras posteriores. En 1977, Borsuk-Białynicka consideró a Opisthocoelicaudia más cerca de Euhelopus que de Camarasaurus , colocándolo en la subfamilia Euhelopodinae.[8] Un estudio de 1981 por Walter Coombs y Ralph Molnar , por otro lado, lo consideró un miembro de la subfamilia Camarasaurinae y por lo tanto un pariente cercano de Camarasaurus.[28] Hoy, tanto Euhelopus como Opisthocoelicaudia están clasificados fuera de Camarasauridae. En 1993, Leonardo Salgado y Rodolfo Coria mostraron a Opisthocoelicaudia que representaba un titanosauriano y lo clasificaron dentro de la familia Titanosauridae.[12] El nombre Titanosauridae actualmente es considerado inválido por muchos científicos,[29] en cambio, el nombre Lithostrotia se usa a menudo como equivalente.[11][30]
Dentro de Lithostrotia, se ha encontrado que Opisthocoelicaudia está estrechamente relacionada con los géneros Alamosaurus, Neuquensaurus , Rocasaurus y Saltasaurus, formando juntos la familia Saltasauridae. Las interrelaciones de estos géneros son controvertidas. Muchos científicos consideraron que Opisthocoelicaudia está más estrechamente relacionada con Alamosaurus , con ambos géneros formando un grupo monofilético, el Opisthocoelicaudiinae. Otros científicos llegaron a la conclusión de que el Opisthocoelicaudiinae es parafilético, no forma un grupo natural.[31] Contradiciendo la mayoría de los otros estudios, Upchurch y sus colegas en 2004 argumentaron que Alamosaurus debe colocarse fuera de Saltasauridae como pariente cercano de Pellegrinisaurus y por lo tanto, no está relacionado con Opisthocoelicaudia y los otros.[11]
Filogenia
El siguiente cladograma se basa en Calvo y colegas de 2007, que muestra un Opisthocoelicaudiinae monofilético.[32]
| Saltasauridae |
| ||||||||||||||||||||||||
Relación con Nemegtosaurus
Otro saurópodo de la Formación Nemegt, Nemegtosaurus, solo se conoce por un cráneo. Opisthocoelicaudia, por otro lado, carece tanto del cráneo como del cuello, lo que impide una comparación directa y genera sospechas de que puede representar un sinónimo de Nemegtosaurus. De acuerdo con el Código Internacional de Nomenclatura Zoológica, el nombre más antiguo tiene prioridad sobre los sinónimos más jóvenes, si se demuestra que Opisthocoelicaudia es sinónimo de Nemegtosaurus, el nombre Nemegtosaurus seguirá siendo válido, mientras que Opisthocoelicaudia dejaría de ser válido.[27]

Tanto Opisthocoelicaudia como Nemegtosaurus fueron descubiertos durante la expedición conjunta de Polonia y Mongolia de 1965. Antes de que los restos fueran preparados y descritos, la tripulación de la expedición creía que ambos hallazgos pertenecían a la misma especie de saurópodo.[23] En 1977, Borsuk-Białynicka consideró que Opisthocoelicaudia y Nemegtosaurus representaban géneros separados porque Nemegtosaurus en ese momento se consideraba miembro de los Dicraeosauridae, mientras que Opisthocoelicaudia parecía ser un representante de un grupo diferente, los Camarasauridae.[8]
Actualmente, tanto Opisthocoelicaudia como Nemegtosaurus están clasificados dentro de los Titanosauria, y Jeffrey Wilson declaró en 2005 que no se puede descartar la sinonimia.[33] Currie y sus colegas, en 2003 y 2017, argumentaron que una sinonimia es muy probable a la luz de los nuevos descubrimientos fósiles en la Formación Nemegt.[27][19] Después de reubicar la cantera original de Nemegtosaurus, estos investigadores excavaron huesos postcraneales del holotipo de Nemegtosaurus, incluido el centro de una vértebra caudal y los huesos de las extremidades posteriores, lo que permitió, por primera vez, una comparación directa entre Nemegtosaurus y Opisthocoelicaudia, basadas en elementos superpuestos. Se descubrió que estos elementos postcraneales son muy similares a las partes correspondientes del holotipo de Opisthocoelicaudia. Lo que es más importante, el centro caudal descubierto es opistocélico, una característica de diagnóstico de Opisthocoelicaudia, lo que sugiere que ambos géneros están estrechamente relacionados o son sinónimos. Además, estos autores señalaron que ninguna de las 32 localidades conocidas de saurópodos de la Formación Nemegt reveló evidencia de la presencia de más de una especie de saurópodos.[19] En su primera descripción, Borsuk-Białynicka de 1977 argumentó que diferentes géneros de saurópodos que comparten el mismo hábitat no son nada inusual, como es evidente en la Formación Morrison de América del Norte.[8] Currie y sus colegas, sin embargo, enfatizaron que la fauna de dinosaurios de la Formación Nemegt era fundamentalmente diferente, ya que los dinosaurios más grandes estaban presentes con solo unas pocas especies por clado, lo que indica un hábitat duro y geográficamente restringido. Sin embargo, todavía faltan pruebas definitivas de la sinonimia sugerida y se requerirían elementos superpuestos adicionales antes de que Opisthocoelicaudia y Nemegtosaurus puedan ser declarados formalmente sinónimos.[19]
Paleoecología

La Formación Nemegt se depositó en el Cretácico Superior, aunque se desconoce su edad exacta ya que nunca se ha fechado radiométricamente. Según diferentes autores, la formación es del Campaniense tardío al Maastrichtiense temprano o medio.[34] Los sedimentos de la Formación Nemegt fueron depositados en una llanura atravesada por ríos.[24] El clima era cálido y subhúmedo con sequías estacionales,[34] y los suelos eran relativamente secos.[35] Sin embargo, la Formación Nemegt fue más húmeda que la subyacente y por lo tanto más antigua, Barun Goyot y Djadochta, que muestran un clima semiárido.[34]
La fauna de la Formación Nemegt incluye animales acuáticos o anfibios como peces, tortugas y cocodrilos, así como pájaros y los abundantes dinosaurios de tamaño mediano a grande, mientras que los vertebrados terrestres más pequeños como lagartos y mamíferos son raros.[34] Los dinosaurios terópodos son muy diversos en el Nemegt e incluyen el abundante tiranosáurido Tarbosaurus,[34] que podría haber cazado a Opisthocoelicaudia.[36] El único otro saurópodo conocido es Nemegtosaurus, que se conoce de un solo cráneo. Los ornitisquios están representados por los hadrosáuridos "pico de pato", incluidos los muy comunes Saurolophus, los paquicefalosaurianos de cráneos gruesos y los anquilosáuridos fuertemente blindados. Los neoceratopsios están ausentes, a pesar de estar presentes en las formaciones más antiguas de Barun Goyot y Djadochta.[34] Otros hallazgos importantes de dinosaurios de la misma localidad que Opisthocoelicaudia incluyen el trodóntido Borogovia[37] y el anquilosáurido Tarchia.[38]
Paleobiología
Postura

Originalmente, Borsuk-Białynicka asumió que en posición estándar el cuello era horizontal o inclinado ligeramente hacia abajo. Esto se basó en la reconstrucción del ligamento nucal , que se extiende sobre las vértebras cervicales y dorsales y sirve para soportar el peso de la cabeza y el cuello.[8] Aunque en varias reconstrucciones posteriores se previó un cuello ascendente curvo en forma de S, similar a un cisne, después de representaciones similares de saurópodos más conocidos, estudios recientes sostienen que los cuellos de saurópodos eran relativamente rectos y se transportaban más horizontalmente.[39]

La espalda también fue reconstruida en una orientación más o menos horizontal por Borsuk-Białynicka,[8] seguido de la mayoría de las representaciones posteriores. En un estudio de 2007, Daniela Schwarz y sus colegas sugirieron que la parte posterior se hundiera hacia la parte posterior. Según estos investigadores, el omóplato habría estado inclinado en un ángulo horizontal de 55-65°, mucho más pronunciado de lo que se pensaba anteriormente, lo que daría lugar a una región elevada del hombro.[40] Con la columna vertebral del tronco y el cuello en una línea relativamente recta, esto daría como resultado una posición elevada de la cabeza.[39]
Opisthocoelicaudia puede haber sido capaz de levantarse sobre sus extremidades posteriores para alimentarse, usando su cola como una tercera pata. En 1977, Borsuk-Białynicka citó varias características esqueléticas que podrían haber estado relacionadas con la cría, incluidas las vértebras opistocoelicas de la parte anterior de la cola, que, según este autor, habrían hecho que la cola fuera más flexible que en otros saurópodos. Las características de la pelvis, como el estante engrosado del acetábulo, el ilion y la sínfisis púbica fusionada pueden haber permitido que la pelvis resistiera el estrés de pararse en dos patas.[8]
Heinrich Mallison en 2011 argumentó que Opisthocoelicaudia pudo haber podido inclinar la parte anterior de la cola contra la parte posterior, produciendo una hebilla en la sección media. Por lo tanto, la parte anterior habría sido más recta cuando se levantaba que en otros saurópodos.[41] En 2005, Wilson asumió que la cría era una innovación no solo de Opisthocoelicaudia sino también de géneros relacionados dentro de la subfamilia Saltasaurinae. Las características comunes de estos géneros, como la cola acortada, pueden haber evolucionado como adaptaciones para alzarse.[31]
Huellas

Las huellas de la Formación Nemegt eran desconocidas hasta 2003, cuando Currie y sus colegas describieron varios ejemplos de la localidad de Nemegt. La mayoría de estas huellas pertenecían a hadrosáuridos, probablemente Saurolophus, mientras que dos fueron dejados por un gran terópodo, probablemente Tarbosaurus y otros dos por el retropié de un saurópodo. Las huellas de saurópodos fueron asignadas a Opisthocoelicaudia, que, según estos autores, mostró una morfología del pie trasero correspondiente y probablemente fue el único saurópodo conocido y, por lo tanto, la única especie potencial de creador de huellas de la Formación Nemegt ya que Nemegtosaurus es considerado un sinónimo. Las huellas se dejaron en el lodo suave y húmedo de puntos poco profundos o recién secos a lo largo de un río y posteriormente se llenaron de arena. Hoy solo queda el relleno de arena, y la piedra de lodo que la recubre ha sido erosionada.[27] La huella mejor conservada mide 63 centímetros de ancho, por lo que probablemente fue creada por un individuo más grande que el espécimen tipo. Aunque la superficie de la parte inferior es difícil de obtener, las superficies verticales están muy bien conservadas, lo que hace que esta pista sea una de las pistas de saurópodos mejor conservadas conocidas. Se pueden distinguir cuatro impresiones digitales, con dos o tres que muestran impresiones de garras. Los dedos de los pies eran casi perpendiculares. Incluso se ha conservado una impresión de la piel por encima de la impresión del primer dedo del pie, que muestra las escamas no superpuestas, cada una con un diámetro promedio de 14 milímetros. El pie del creador de la pista fue probablemente un poco más largo que ancho. La segunda pista es mucho menos profunda que la primera, pero muestra impresiones de dígitos medios bien conservadas con un alto grado de detalle.[27]
Aunque se informaron varias pistas de saurópodos adicionales en cuentas posteriores, continuaron siendo raras en relación con las pistas de hadrosáuridos y terópodos mucho más comunes. Brennan Stettner y sus colegas, en 2017, informaron sobre las huellas descubiertas durante una expedición de 2007 a la localidad de Nemegt. El mejor conservado de estos, una impresión muy grande de 76 centímetros de largo de un retropié, presenta una parte inferior muy bien conservada que muestra almohadillas digitales y cuatro dígitos dirigidos hacia afuera, los primeros tres muestran garras.[42] También en 2017, Judai Nakajima y sus colegas describieron una impresión en forma de riñón como la primera impresión de una mano de saurópodo descubierta en la formación.[43]
Referencias
- Seebacher, F. (2001). «A new method to calculate allometric length-mass relationships of dinosaurs». Journal of Vertebrate Paleontology 21 (1): 51-60. doi:10.1671/0272-4634(2001)021[0051:ANMTCA]2.0.CO;2. Parámetro desconocido
|citeseerx=ignorado (ayuda) - Holtz, Thomas R., Jr.; Rey, Luis V. (2007). Dinosaurs: the most complete, up-to-date encyclopedia for dinosaur lovers of all ages. New York: Random House. ISBN 978-0-375-82419-7. (requiere registro).
- Barsbold, R. (1997). «Mongolian dinosaurs». En Currie, P.J.; Padian, K., eds. Encyclopedia of Dinosaurs. Academic Press. pp. 447–450. ISBN 978-0-12-226810-6.
- Paul, Gregory S. (1997). «Dinosaur models: the good, the bad, and using them to estimate the mass of dinosaurs». En Wolberg, D.L.; Stump, E.; Rosenberg, G.D., eds. Dinofest International. Proceedings of a symposium held at Arizona State University. Academy of Natural Sciences, Philadelphia. pp. 129-154 (152).
- Anderson, J.F.; Hall-Martin, A.; Russell, D.A. (2009). «Long-bone circumference and weight in mammals, birds and dinosaurs». Journal of Zoology 207 (1): 53-61. doi:10.1111/j.1469-7998.1985.tb04915.x.
- Packard, G.C.; Boardman, T.J.; Birchard, G.F. (2009). «Allometric equations for predicting body mass of dinosaurs». Journal of Zoology 279 (1): 102-110. doi:10.1111/j.1469-7998.2009.00594.x.
- Benson, R. B. J.; Campione, N. S. E.; Carrano, M. T.; Mannion, P. D.; Sullivan, C.; Upchurch, P.; Evans, D. C. (2014). «Rates of Dinosaur Body Mass Evolution Indicate 170 Million Years of Sustained Ecological Innovation on the Avian Stem Lineage». PLoS Biology 12 (5): e1001853. PMC 4011683. PMID 24802911. doi:10.1371/journal.pbio.1001853.
- Borsuk-Białynicka, M.M. (1977). «A new camarasaurid sauropod Opisthocoelicaudia skarzynskii gen. n., sp. n. from the Upper Cretaceous of Mongolia». Palaeontologia Polonica 37 (5): 5-64.
- Paul, Gregory S. (2019). «Determining the largest known land animal: A critical comparison of differing methods for restoring the volume and mass of extinct animals». Annals of the Carnegie Museum 85 (4): 335-358.
- Apesteguía, S. (2005). «Evolution of the Hyposphene-Hypantrum Complex within Sauropoda». En Tidwell, V.; Carpenter, K., eds. Thunder-Lizards: The Sauropodomorph Dinosaurs. Indiana University Press. ISBN 978-0-253-34542-4.
- Upchurch, P.; Barret, P.M.; Dodson, P. (2004). «Sauropoda». En Weishampel, D.B.; Dodson, P.; Osmólska, H., eds. The Dinosauria (2nd edición). University of California Press. pp. 259–322. ISBN 978-0-520-25408-4.
- Salgado, L.; Coria, R.A. (1993). «Consideraciones sobre las relaciones filogeneticas de Opisthocoelicaudia skarzynskii (Sauropoda) del Cretácico superior de Mongolia». Jornadas Argentinas de Paleontologia de Vertebrados, Resumenes 10.
- Wilson, J. (2002). «Sauropod dinosaur phylogeny: critique and cladistic analysis». Zoological Journal of the Linnean Society 136 (2): 217-276. doi:10.1046/j.1096-3642.2002.00029.x. Parámetro desconocido
|doi-access=ignorado (ayuda) - Taylor, M.P.; Wedel; M.J. (2013). «Why sauropods had long necks; and why giraffes have short necks». PeerJ 1: e36. ISSN 2167-8359. PMC 3628838. PMID 23638372. doi:10.7717/peerj.36.
- Carrano, M.T. (2005). «The Evolution of Sauropod Locomotion: Morphological Diversity of a Secondarily Quadrupedal Radiation». En Rogers, K.C.; Wilson, J.A., eds. The Sauropods: Evolution and Paleobiology. University of California Press. ISBN 978-0-520-24623-2.
- Wilson, J.A.; Carrano, M.T. (1999). «Titanosaurs and the origin of "wide-gauge" trackways: a biomechanical and systematic perspective on sauropod locomotion». Paleobiology 52 (2): 252-267. doi:10.1017/S0094837300026543.
- Apesteguía, S. (2005). «Evolution of the Titanosaur Metacarpus». En Tidwell, V.; Carpenter, K., eds. Thunder-Lizards: The Sauropodomorph Dinosaurs. Indiana University Press. pp. 321–345. ISBN 978-0-253-34542-4.
- González Riga, B.J.; Calvo, J.O.; Porfiri, J. (2008). «An articulated titanosaur from Patagonia (Argentina): New evidence of neosauropod pedal evolution». Palaeoworld 17: 33-40. doi:10.1016/j.palwor.2007.08.003.
- Currie, P.J.; Wilson, J.A.; Fanti, F.; Mainbayar, B.; Tsogtbaatar, Khishigjav (in press). "Rediscovery of the type localities of the Late Cretaceous Mongolian sauropods Nemegtosaurus mongoliensis and Opisthocoelicaudia skarzynskii: Stratigraphic and taxonomic implications". Palaeogeography, Palaeoclimatology, Palaeoecology.
- d’Emic, M.D.; Wilson, J.A.; Chatterjee, S. (2009). «The titanosaur (Dinosauria: Sauropoda) osteoderm record: review and first definitive specimen from India». Journal of Vertebrate Paleontology 29: 165-177. doi:10.1671/039.029.0131.
- Carrano, M.T.; Michael, D.D. (2015). «Osteoderms of the titanosaur sauropod dinosaur Alamosaurus sanjuanensis Gilmore, 1922». Journal of Vertebrate Paleontology 35 (1): e901334. doi:10.1080/02724634.2014.901334.
- Kielan-Jaworowska, Z.; Dovchin, N. (1968). «Narrative of the Polish-Mongolian Expeditions 1963–1965». Palaeontologia Polonica 19: 7-40.
- Kielan-Jaworowska, Z. (2013). «The Polish-Mongolian paleontological expeditions, 1963–1971, and the nomadic expedition, 2002». In Pursuit of Early Mammals. Indiana University Press. pp. 35-70. ISBN 978-0-253-00817-6.
- Gradziński, Ryszard (1969). «Sedimentation of dinosaur-bearing Upper Cretaceous deposits of the Nemegt Basin, Gobi Desert». Palaeontologia Polonica 21: 147-249. Archivado desde el original el 15 de julio de 2020. Consultado el 21 de julio de 2020.
- Hentschel, E., & Wagner, G. (1990). Zoologisches Wörterbuch. Tiernamen, allgemeinbiologische, anatomische, physiologische Termini und biographische Daten (4rd edition). Stuttgart: Gustav Fischer.
- Maryańska, T. (2000). «Sauropods from Mongolia and the former Soviet Union». En Benton, M.J.; Shishkin, M.A.; Unwin, D.M. et al., eds. The Age of Dinosaurs in Russia and Mongolia. Cambridge University Press. pp. 457–458. ISBN 978-0-521-54582-2.
- Currie, P.J.; Badamgarav, D.; Koppelhus, E.B. (2003). «The first Late Cretaceous footprints from the locality in the Gobi of Mongolia». Ichnos 10: 1-12. doi:10.1080/10420940390235071.
- Coombs, W.P.; Molnar, R.E. (1981). «Sauropoda (Reptilia, Saurischia) from the Cretaceous of Queensland». Memoirs of the Queensland Museum 20 (2): 351-373.
- Wilson, J.A.; Upchurch, P. (2003). «A Revision of Titanosaurus Lydekker (Dinosauria – Sauropoda), the first dinosaur genus with a 'Gondwanan' distribution». Journal of Systematic Palaeontology 1 (3): 125-160. doi:10.1017/s1477201903001044.
- D’emic, M.D. (2012). «The early evolution of titanosauriform sauropod dinosaurs». Zoological Journal of the Linnean Society 166 (3): 624-671. doi:10.1111/j.1096-3642.2012.00853.x.
- Wilson, J. (2005). «Overview of Sauropod Phylogeny and Evolution». En Rogers, K.C.; Wilson, J.A., eds. The Sauropods: Evolution and Paleobiology. University of California Press. ISBN 978-0-520-24623-2.
- Calvo, J.O., Porfiri, J.D., González-Riga, B.J., Kellner, A.W. (2007). «A new Cretaceous terrestrial ecosystem from Gondwana with the description of a new sauropod dinosaur». Anais da Academia Brasileira de Ciências 79 (3): 529-541. PMID 17768539. doi:10.1590/S0001-37652007000300013. Parámetro desconocido
|doi-access=ignorado (ayuda) - Wilson, J.A. (2005). «Redescription of the Mongolian sauropod Nemegtosaurus mongoliensis Nowinski (Dinosauria: Saurischia) and comments on Late Cretaceous sauropod diversity». Journal of Systematic Palaeontology 3 (3): 283-318. doi:10.1017/S1477201905001628.
- Osmólska, H. (1997). «Nemegt Formation». Encyclopedia of Dinosaurs. Academic Press. pp. 471–472. ISBN 978-0-12-226810-6.
- Jerzykiewicz, Tomasz; Dale A. Russell (1991). «Late Mesozoic stratigraphy and vertebrates of the Gobi Basin». Cretaceous Research 12 (4): 345-377. ISSN 0195-6671. doi:10.1016/0195-6671(91)90015-5.
- Hurum, J.H.; Sabath, K. (2003). «Giant theropod dinosaurs from Asia and North America: Skulls of Tarbosaurus bataar and Tyrannosaurus rex compared». Acta Palaeontologica Polonica 48 (2): 161-190.
- Osmólska, H. (1987). «Borogovia gracilicrus gen. et sp. n., a new troodontid dinosaur from the Late Cretaceous of Mongolia». Acta Palaeontologica Polonica 32: 133-150.
- Maryańska, T. (1969). «Remains of armoured dinosaurs from the uppermost Cretaceous in Nemegt Basin, Gobi Desert». Palaeontologia Polonica 21: 23-32. Archivado desde el original el 12 de julio de 2020. Consultado el 21 de julio de 2020.
- Stevens, K.A. (2013). «The articulation of sauropod necks: methodology and mythology». PLoS ONE 8 (10): e78572. Bibcode:2013PLoSO...878572S. PMC 3812995. PMID 24205266. doi:10.1371/journal.pone.0078572.
- Schwarz, D.; Frey, E.; Meyer, C. (2007). «Novel reconstruction of the orientation of the pectoral girdle in sauropods». The Anatomical Record 290 (1): 32-47. PMID 17441196. doi:10.1002/ar.20405.
- Mallison, H. (2011). «Rearing Giants: Kinetic-Dynamic Modeling of Sauropod Bipedal and Tripodal Poses». En Klein, N.; Remes, K.; Gee, C.T. et al., eds. Biology of the Sauropod Dinosaurs: Understanding the Life of Giants. pp. 239-320. ISBN 978-0-253-35508-9.
- Stettner, B.; Persons, S.; Currie, P.J. (in press). "A giant sauropod footprint from the Nemegt Formation (Upper Cretaceous) of Mongolia". Palaeogeography, Palaeoclimatology, Palaeoecology.
- Nakajima, J.; Kobayashi, Y.; Chinzorig, T.; Tanaka, T.; Takasaki, R., Tsogtbaatar, K.; Currie, P.J.; Fiorillo, A.R. (in press). "Dinosaur tracks at the Nemegt locality: Paleobiological and paleoenvironmental implications". Palaeogeography, Palaeoclimatology, Palaeoecology.
| Control de autoridades |
|
|---|
 Datos: Q133088
Datos: Q133088 Multimedia: Opisthocoelicaudia / Q133088
Multimedia: Opisthocoelicaudia / Q133088