Pinocitosis
La pinocitosis es un tipo de endocitosis que consiste en la captación de material del espacio extracelular por invaginación de la membrana citoplasmática eucariota.[1] Esta membrana se repliega creando un endosoma llamado vesícula pinocítica o pinozoma que se desprende hacia el interior celular con las moléculas disueltas o en suspensión en su interior.
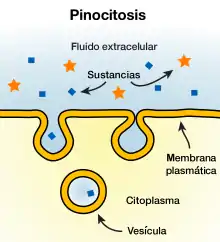
Junto con la fagocitosis, constituyen los dos tipos principales de endocitosis. La pinocitosis se diferencia de la fagocitosis principalmente en que la pinocitosis se da en todas las células y afecta a las moléculas (como la mayor parte de las proteínas) y la fagocitosis solo se da en ciertas células especializadas de los mamíferos y procesa partículas mayores (como las bacterias).[2] Las vesículas pinocíticas suele medir entre 100 y 200 nanómetros de diámetro.[3] Existen cuatro tipos de mecanismos pinocíticos básicos: la macropinocitosis, la endocitosis mediada por la clatrina, la endocitosis mediada por cavéolas y la endocitosis independiente de la clatrina y de las cavéolas.[2]
En el ser humano, este fenómeno se observa en células de la mucosa intestinal, cuando estas permiten el ingreso de vesículas de grasa durante la absorción de nutrientes. Otro tipo de célula en la cual se la ha observado frecuentemente es el óvulo humano. Cuando el óvulo madura en el ovario de la mujer, se rodea de "células nodrizas". Aparentemente, estas células ceden alimentos disueltos al óvulo, que los incorpora por pinocitosis.[cita requerida]
Proceso de pinocitosis
El proceso de la pinocitosis se da de forma continua en la membrana de la mayoría de las células; y la velocidad a la que se da es variable, siendo especialmente rápida por ejemplo en los macrófagos ya que cada minuto aproximadamente un 3% de su membrana es absorbida en forma vesícula. La velocidad a la que se forman las vesículas puede aumentar cuando las moléculas entran en contacto con la pared celular.[3] La pinocitosis se puede medir a través de la acumulación de marcadores presentes en el líquido intracelular (como enzimas u otros compuestos), y la cantidad de partículas internalizadas por la célula depende tanto de la cantidad de los marcadores presentes en el medio extracelular como del volumen de las vesículas.[2] La energía necesaria para realizar este proceso es bastante considerable, y es suministrada por el adenosín trifosfato, así como por la presencia de iones calcio.[3]
Las vesículas se originan en la superficie de la membrana celular, generalmente cuando una molécula se une con su receptor específico. Estos receptores están situados en unas zonas de la membrana celular llamadas depresiones revestidas. En la parte interior de estas depresiones se encuentra la proteína fibrilar llamada clatrina, así como otros filamentos contráctiles como la actina y la miosina. Una vez la molécula se une a los receptores la vesícula revestida de proteínas fibrilares pasa al citoplasma mediante invaginación, englobando la molécula y una parte del fluido extracelular.[3] Las paredes de la vesícula pinocítica se fusionan en lo que se denomina un endosoma temprano, proceso en el que intervienen las proteínas trans-SNARE (v-SNARE y t-SNARE).
En la fase intermedia la vesícula se denomina vesícula digestiva. El proceso de hidrólisis se inicia cuando se unen a la vesícula uno o varios lisosomas que vacían las enzimas llamadas hidrolasas ácidas en su interior. Estas hidrolasas digieren las sustancias que se encuentran dentro de la vesícula como las proteínas, los hidratos de carbono o los lípidos, y los transforman en moléculas más pequeñas de aminoácidos, glucosa, o fosfatos entre otras, que atraviesan la membrana de la vesícula hacia el citoplasma.[3]
En la última fase la vesícula recibe el nombre de cuerpo residual, y en la mayoría de los casos es excretada a través de la membrana celular debido a que está formada por sustancias no digeribles, en un proceso que se llama exocitosis.[3]
En la pinocitosis mediada por la clatrina una vez que la vesícula entra en el citoplasma el revestimiento de clatrina se degrada y los trisqueliones (formados por moléculas de clatrina) quedan libres en el citoplasma.
Macropinocitosis
La macropinocitosis es un mecanismo endocítico independiente de la clatrina que puede activarse en prácticamente todas las células animales. El diámetro de una vesícula macropinocítica, o macropinosoma, es mayor a un micrómetro (1 µm). En la mayoría de los tipos de células no ocurre de forma continua, sino que se induce durante un tiempo limitado en respuesta a la activación del receptor de la superficie celular por cargas específicas, incluidos factores de crecimiento, ligandos de integrinas, restos de células apoptóticas y algunos virus. Otros procesos relacionados con la macropinocitosis son la migración celular dirigida, la activación de células dendríticas presentadoras de antígenos.[2]
Estos ligandos activan mecanismos de señalización compleja relacionados con las enzimas GTPasa, lo que da como resultado un cambio en la dinámica de la actina y la formación de protuberancias en la superficie celular, llamadas volantes.[2] Cuando los volantes colapsan de nuevo sobre la membrana, se forman grandes vesículas endocíticas llenas de líquido, llamadas macropinosomas, que pueden aumentar la captación de líquido hasta diez veces de forma momentánea. Los macropinosomas se acidifican y luego se fusionan con endosomas tardíos o endolisosomas, y se terminan degradando.[4]
Algunas bacterias inducen la macropinocitosis mediante la inyección de toxinas en la célula para aumentar la producción de macropinosomas, que ayudan a su propia proliferación.[5]
Endocitosis mediada por clatrina
La endocitosis mediada por clatrina se da en todas las células de los mamíferos, y genera un aporte continuo de nutrientes esenciales para la célula como la lipoproteína de baja densidad que se adhiere al receptor LDL o la transferrina que se adhiere a los receptores Tfn. Se denomina así porque la clatrina es la principal proteína que estructura las cavidades que en la invaginación llegan a formar las vesículas pinocíticas.[2]
Este tipo de pinocitosis cumple diversas funciones importantes en distintas células. Una de ellas es la modulación de la transducción de señales en el desarrollo de tejidos y de órganos en el organismo controlando los niveles de receptores de señales en la superficie celular.[6] También regulan los niveles homeostáticos de la célula y del plasma controlando la cantidad de bombas de sodio-potasio presentes en la membrana celular, o capturando las proteínas plasmáticas después del filtrado de los riñones.[2] También ayudan a regular el potencial de membrana modulando la cantidad de canales de calcio presentes en las sinapsis o intervienen en el reciclaje de la vesícula sináptica una vez realizada la neurotransmisión.[7]
La endocitosis mediada por clatrina se denominaba anteriormente endocitosis mediada por receptores, pero se dejó de usar al ir descubriendo que la mayor parte de los canales pinocíticos involucran un receptor-ligando específico.[2]
Endocitosis mediada por cavéolas
Este tipo de pinocitosis acostumbra a ser más lenta que el resto y las vesículas generadas son más pequeñas (entre 50 y 60 nm de diámetro), por lo que se considera que en general no es un mecanismo que contribuya de forma significativa a la captación de fluido. Pese a esto en cierto tipo de células como las endotelias o los adipocitos, las cavéolas constituyen entre un 10 y un 20% de la superficie de la membrana celular.[2]
Endocitosis independiente de la clatrina y de las cavéolas
El hecho de que este mecanismo se defina de forma negativa es un indicador de que todavía no se conoce el mecanismo de formación. Pese a esto existen algunos ejemplos de cavidades independientes de la clatrina y de las cavéolas. Estos, más generalmente conocidos como "balsas" (rafts en inglés), son estructuras lipídicas pequeñas, de 40 a 50 nm de diámetro, que se difunden libremente en la superficie celular; y cumplen funciones específicas como la clasificación de proteínas o glucolípidos de membrana. Estas balsas pueden ser captadas dentro de otras vesículas, e internalizadas en el proceso endocítico.[2]
Por ejemplo tanto la toxina shiga como la toxina colérica se unen a los glucolípidos de este tipo de balsas para ser internalizadas posteriormente dentro de vesículas mediadas por la clatrina.[2]
Etimología
El término pinocitosis fue propuesto por W. H. Lewis en 1931, y proviene de πίνω (pinó-, "beber") y κύτος (kútos, "contenedor, receptáculo") en griego antiguo.[8]
Referencias
- Guyton, Arthur C.; Hall, John Edward (2006). Textbook of Medical Physiology. Elsevier España. ISBN 9788481749267. Consultado el 30 de octubre de 2019.
- Conner, Sean D.; Schmid, Sandra L. (2003-03). «Regulated portals of entry into the cell». Nature (en inglés) 422 (6927): 37-44. ISSN 1476-4687. doi:10.1038/nature01451. Consultado el 4 de octubre de 2021.
- Hall, John E. (2001). Tratado de fisiología médica (10. ed. en español edición). McGraw-Hill Interamericana. p. 20. ISBN 970-10-3599-2. OCLC 49608187. Consultado el 28 de septiembre de 2021.
- Alberts, Bruce (2015). Molecular biology of the cell (Sixth edition edición). p. 732. ISBN 978-0-8153-4432-2. OCLC 887605755. Consultado el 28 de septiembre de 2021.
- Steele-Mortimer, Olivia; Knodler, Leigh A.; Finlay, B. Brett (2000-02). «Poisons, Ruffles and Rockets: Bacterial Pathogens and the Host Cell Cytoskeleton: Bacterial Pathogens and the Host Cell Cytoskeleton». Traffic (en inglés) 1 (2): 107-118. doi:10.1034/j.1600-0854.2000.010203.x. Consultado el 4 de octubre de 2021.
- Seto, E. S. (1 de junio de 2002). «When cell biology meets development: endocytic regulation of signaling pathways». Genes & Development 16 (11): 1314-1336. ISSN 0890-9369. doi:10.1101/gad.989602. Consultado el 6 de octubre de 2021.
- De Camilli, Pietro; Takei, Kohji (1996-03). «Molecular Mechanisms in Synaptic Vesicle Endocytosis and Recycling». Neuron 16 (3): 481-486. ISSN 0896-6273. doi:10.1016/s0896-6273(00)80068-9. Consultado el 6 de octubre de 2021.
- Michaelis, A.; Green, M. M. (1991). Glossary of Genetics : Classical and Molecular (Fifth edition edición). Springer Berlin Heidelberg. ISBN 978-3-642-75333-6. OCLC 840295988. Consultado el 28 de septiembre de 2021.
 Datos: Q321362
Datos: Q321362