Ley de Hardy-Weinberg
En genética de poblaciones, el principio de Hardy-Weinberg (PHW) (también equilibrio de Hardy-Weinberg, ley de Hardy-Weinberg o caso de Hardy-Weinberg ) establece que la composición genética de una población permanece en equilibrio mientras no actúe la selección natural ni ningún otro factor y no se produzca ninguna mutación. Es decir, la herencia mendeliana, por sí misma, no engendra cambio evolutivo. Recibe su nombre del matemático inglés G. H. Hardy y del médico alemán Wilhelm Weinberg, que establecieron el teorema independientemente en 1908.[1]
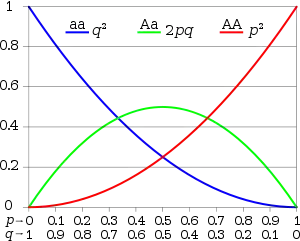
En el lenguaje de la genética de poblaciones, la ley de Hardy-Weinberg afirma que, bajo ciertas condiciones, tras una generación de apareamiento al azar, las frecuencias de los genotipos de un locus individual se fijarán en un valor de equilibrio particular. También especifica que esas frecuencias de equilibrio se pueden representar como una función sencilla de las frecuencias alélicas en ese locus. En el caso más sencillo, con un locus con dos alelos A y a, con frecuencias alélicas de p y q respectivamente, el PHW predice que la frecuencia genotípica para el homocigoto dominante AA es p2, la del heterocigoto Aa es 2pq y la del homocigoto recesivo aa es q2. El principio de Hardy-Weinberg es una expresión de la noción de una población que está en "equilibrio genético", y es un principio básico de la genética de poblaciones.
Suposiciones
Las suposiciones originales del equilibrio de Hardy-Weinberg (EHW) eran que el organismo en consideración:
- Sea diploide, y el carácter en consideración no esté en un cromosoma que tiene un número distinto de copias en cada sexo, como el cromosoma X en los humanos (es decir, que el carácter sea autosómico)
- Se reproduzca sexualmente, bien monoicamente o dioicamente
- Tenga generaciones discretas
Además, la población en consideración está idealizada, esto es:
- Existe apareamiento aleatorio en la población (a este tipo de población se le conoce como "población panmíctica")
- Tiene un tamaño infinito (o lo bastante grande para minimizar el efecto de la deriva genética)
y no experimenta:
El primer grupo de suposiciones son un requisito de las matemáticas implicadas. Es relativamente sencillo expandir la definición del EHW para que incluya modificaciones de estas suposiciones, por ejemplo las de los caracteres ligados al sexo. Las otras suposiciones son inherentes al principio de Hardy-Weinberg.
Cuando se discuten varios factores, se utiliza una población de Hardy-Weinberg como referencia. No es sorprendente que estas poblaciones sean estáticas.
Derivación
Una mejor, aunque equivalente, descripción probabilística del PHW es que los alelos de la siguiente generación para cualquier individuo se eligen aleatoria e independientemente unos de otros. Considérense dos alelos A y a con frecuencias en la población de p y q respectivamente. Las distintas maneras de formar nuevos genotipos se pueden derivar utilizando un cuadro de Punnett, por el que la fracción en cada celda es igual al producto de las probabilidades de la fila y la columna.
| Hembras | |||
|---|---|---|---|
| A (p) | a (q) | ||
| Machos | A (p) | AA (p2) | Aa (pq) |
| a (q) | Aa (pq) | aa (q2) | |
Las tres posibles frecuencias genotípicas finales de la descendencia son:



Estas frecuencias se llaman frecuencias de Hardy-Weinberg (o proporciones de Hardy-Weinberg). Esto se consigue en una generación, y solo hace falta suponer un apareamiento aleatorio en una población de tamaño infinito.
A veces una población se crea juntando machos y hembras con distintas frecuencias alélicas. En este caso, la suposición de una sola población queda violada hasta la siguiente generación, de manera que la primera generación no tendrá equilibrio de Hardy-Weinberg. Las generaciones sucesivas sí tendrán equilibrio de Hardy-Weinberg.
Ejemplo
A continuación se ejemplifica la ley de Hardy-Weinberg a partir de un ejemplo real: la enfermedad metabólica hereditaria fenilcetonuria.[2]
Los niños con fenilcetonuria no pueden procesar la fenilalanina, un aminoácido de las proteínas, por lo que la fenilalanina se acumula en la sangre causando daños cerebrales y retraso mental. Esta enfermedad es provocada por un gen recesivo cuando se da una situación de homocigosis aa. Siendo p la frecuencia del alelo sano y q la del alelo "defectuoso", se calculara la incidencia de los portadores de la combinación aa.
Si se realiza un cruzamiento de dos portadores Aa, en donde permanece oculto el gen recesivo, los genotipos obtenidos en la siguiente generación serán los siguientes:
| A (p) | a (q) | |
| A (p) | AA (p²) | Aa (pq) |
| a (q) | Aa (pq) | aa (q²) |
Los tres genotipos AA : Aa : aa aparecen en una relación p² : 2pq : q². Si se suman se obtiene la unidad:
- p² + 2pq + q² = (p + q)² = 1.
La frecuencia de los genotipos enfermos de fenilcetonuria es de un 0,0001, un valor correspondiente a q². La frecuencia q del gen a será la raíz cuadrada de 0,0001, es decir, 0,01. La enfermedad tiene una incidencia de 1 cada 10 000 individuos, pero la frecuencia del gen es 100 veces mayor, 1 cada 100. Los genes a se encuentran en el par Aa con una frecuencia
- 2pq = 2q(1 - q) = 2· 0,01·(1 - 0,01) = 0,0198.
Cerca de un 2% de los individuos de la población europea porta, por lo tanto, este gen en el par Aa, lo que da una idea de lo persistente que puede llegar a ser un gen recesivo manteniéndose "clandestino" en heterocigosidad.
Desviaciones del equilibrio de Hardy-Weinberg
Las violaciones de las suposiciones de Hardy-Weinberg pueden causar desviaciones de los valores esperados. Cómo afecta esto a la población depende de las suposiciones que son violadas.
- Apareamiento aleatorio. El PHW establece que la población tendrá las frecuencias genotípicas especificadas (llamadas proporciones de Hardy-Weinberg) tras una generación de apareamiento aleatorio dentro de la población. Cuando suceden violaciones de este requisito, la población no tendrá proporciones de Hardy-Weinberg. Tres de estas violaciones son:
- Endogamia, que provoca un aumento de la homocigosidad en todos los genes.
- Emparejamiento selectivo, que causa un aumento en la homocigosidad de los genes implicados en el carácter que se está seleccionando para el apareamiento (y de los genes que están en desequilibrio de ligamiento con ellos).
- Población de poco tamaño, que causa un cambio aleatorio en las frecuencias genotípicas, especialmente si la población es muy pequeña. Esto es debido al efecto de muestreo, y se llama deriva genética.
Las demás suposiciones afectan a las frecuencias alélicas, pero no afectan por sí mismas al apareamiento aleatorio. Si una población viola alguna de estas, la población seguirá teniendo proporciones de Hardy-Weinberg en cada generación, pero las frecuencias alélicas cambiarán con esa fuerza.
- La selección, en general, hace que cambien las frecuencias alélicas, a menudo con mucha rapidez. Aunque la selección direccional conduce finalmente a la pérdida de todos los alelos excepto el favorecido, algunas formas de selección, como la selección estabilizadora, conducen a un equilibrio sin pérdida de alelos.
- La mutación tendrá un efecto muy sutil en las frecuencias alélicas. Los ritmos de mutación son del orden de 10-4 a 10-8 y el cambio en las frecuencias alélicas será, como mucho, del mismo orden. Las mutaciones recurrentes mantendrán a los alelos en la población, aunque haya una fuerte selección en contra de ellos. La validez del principio de Hardy-Weinberg se basa en la asunción de que no se producen nuevas mutaciones. Si un locus en particular muestra una tasa elevada de mutaciones, entonces habrá un aumento regular en la proporción de alelos mutantes en una población. En la práctica, las mutaciones se producen en casi todos los loci, aunque a diferentes tasas, pero el efecto de su introducción está habitualmente equilibrado por la pérdida de alelos mutantes debido a la reducción de la eficacia biológica de los individuos afectados. Si se observa que una población está en equilibrio de Hardy-Weinberg, se asume generalmente que estos dos factores opuestos tienen efectos aproximadamente iguales.
- La migración enlaza genéticamente dos o más poblaciones. En general, las frecuencias alélicas se harán más homogéneas entre las dos poblaciones. Algunos modelos de migración incluyen inherentemente el apareamiento no aleatorio (el efecto Wahlund, por ejemplo). Para esos modelos, las proporciones de Hardy-Weinberg no serán válidas en general.
Más adelante se explica cómo afectan estas violaciones a las pruebas estadísticas formales del EHW.
Desafortunadamente, las violaciones de las suposiciones del principio de Hardy-Weinberg no significa que la población violará el EHW. Por ejemplo, la selección estabilizadora conduce a una población en equilibrio con proporciones de Hardy-Weinberg. Esta propiedad que enfrenta a la selección con la mutación es la base de muchas estimaciones del ritmo de mutación (equilibrio mutación-selección).
Si una población se junta con otra, con machos y hembras con distintas frecuencias alélicas, la frecuencia alélica de la población masculina seguirá a la de la población femenina, porque todos reciben su cromosoma X de su madre. La población converge hacia el equilibrio muy rápidamente.
Generalizaciones
La derivación simple de arriba puede ser generalizada por más de dos alelos y poliploidía.
Generalización para más de dos alelos
Considerar una frecuencia de extra alelo, . El caso de dos alelos es la expansión binomial de , y así el caso de tres alelos es la expresión trinómica de .




Más generalmente, considerar los alelos A1, ... An dado por la frecuencia de los alelos a ;



dado por todos los homozigotos:

y para todos los heterozigotos:

Generalización para la poliploidía
El principio de Hardy-Weinberg también se puede generalizar para sistemas poliploides, esto es, organismos que tienen más de dos copias de cada cromosoma. Considérese de nuevo solo dos alelos. El caso diploide es la expansión binomial de:
y por tanto el caso poliploide es la expansión binomial de:

donde c es la ploidía. Por ejemplo, con un tetraploide (c = 4):
| Genotipo | Frecuencia |
|---|---|










Dependiendo de si el organismo es un tetraploide 'verdadero' o un anfidiploide quedará determinado el tiempo necesario para que la población alcance el equilibrio de Hardy-Weinberg.
Generalización completa
La fórmula completamente generalizada es la expansión multinomial de :


Aplicaciones
El principio de Hardy-Weinberg se puede aplicar de dos maneras: o bien se considera que una población tiene proporciones de Hardy-Weinberg, en cuyo caso se pueden calcular las frecuencias genotípicas, o si las frecuencias genotípicas de los tres genotipos son conocidas, se pueden comprobar las desviaciones que sean estadísticamente significativas.
Aplicación a casos de dominancia completa
Supóngase que los fenotipos de AA y Aa son indistinguibles, es decir, existe una dominancia completa. Suponiendo que el principio de Hardy-Weinberg se aplica a la población, entonces todavía se puede calcular a partir de f(aa):


y se puede calcular a partir de . Y también una estimación de f(A) y f(Aa) derivadas de y respectivamente. Sin embargo, hay que tener en cuenta que no se puede comprobar el equilibrio de una población utilizando los tests de significancia siguientes, porque esta se asume a priori.



Tests de significancia de la desviación
La comprobación de la desviación del PHW se suele llevar a cabo utilizando la prueba χ² de Pearson, utilizando las frecuencias genotípicas observadas que se han obtenido de los datos y las frecuencias genotípicas esperadas obtenidas mediante el PHW. Para los sistemas en los que hay un gran número de alelos, esto puede ofrecer datos con muchos genotipos vacíos posibles y poca cantidad de genotipos, porque a menudo no hay suficientes individuos en la muestra para representar adecuadamente todas las clases genotípicas. Si este es el caso, entonces la suposición asintótica de la distribución chi-cuadrado no se sostendrá y puede ser necesario utilizar una forma de la prueba exacta de Fisher, que requiere de una computadora para su resolución.
Prueba de desviación χ2 de ejemplo
Estos datos son de E.B. Ford (1971) sobre la polilla Callimorpha dominula, de la que se registraron los fenotipos de una muestra de la población. La distinción entre el genotipo y el fenotipo se considera insignificante. La hipótesis nula es que la población tiene proporciones de Hardy-Weinberg, y la hipótesis alternativa es que la población no tiene proporciones de Hardy-Weinberg.
| Genotipo | Manchas blancas (AA) | Intermedia (Aa) | Pocas manchas (aa) | Total |
|---|---|---|---|---|
| Número | 1469 | 138 | 5 | 1612 |
De la que se pueden calcular las frecuencias alélicas:

y

Por lo que los valores esperados de Hardy-Weinberg son:



La prueba χ² de Pearson establece que:





Hay 1 grado de libertad (los grados de libertad para el test de las proporciones de Hardy-Weinberg son número de fenotipos - número de alelos). El nivel de significancia para 1 grado de libertad es 3,84, y como este valor de χ2 es menor, se acepta la hipótesis nula que decía que la población tiene proporciones de Hardy-Weinberg.
Prueba exacta de Fisher
La prueba exacta de Fisher se puede aplicar para comprobar si existen proporciones de Hardy-Weinberg. Como la prueba está condicionado por las frecuencias alélicas, p y q, el problema se puede entender como la comprobación del número adecuado de heterocigotos. De esta forma, la hipótesis de las proporciones de Hardy-Weinberg queda violada si el número de heterocigotos es muy grande o muy pequeño. Las probabilidades condicionadas para el heterocigoto, dadas las frecuencias alélicas, las proporciona Emigh (1980) de la forma
![{\displaystyle prob[n_{12}|n_{1}]={\frac {{n} \choose {n_{11},n_{12},n_{22}}}{{2n} \choose {n_{1}}}}2^{n_{12}},}](../I/e16bb5375178d25b53c7dc4db83a3b04ec5d36a6.svg)
donde n11, n12 y n22 son los números observados para los tres genotipos, AA, Aa y aa respectivamente, y n1 es el número de alelos A, donde .

Un ejemplo Utilizando uno de los ejemplos de Emigh (1980), se puede considerar el caso en el que n = 100 y p = 0,34. En la tabla 4 se muestran los heterocigotos observados posibles y su nivel de significancia exacto.
| Número de heterocigotos | Nivel de significancia |
|---|---|
| 0 | 0,000 |
| 2 | 0,000 |
| 4 | 0,000 |
| 6 | 0,000 |
| 8 | 0,000 |
| 10 | 0,000 |
| 12 | 0,000 |
| 14 | 0,000 |
| 16 | 0,000 |
| 18 | 0,001 |
| 20 | 0,007 |
| 22 | 0,034 |
| 34 | 0,067 |
| 24 | 0,151 |
| 32 | 0,291 |
| 26 | 0,474 |
| 30 | 0,730 |
| 28 | 1,000 |
Utilizando esta tabla, se busca el nivel de significancia de la prueba basándose en el número observado de heterocigotos. Por ejemplo, si se han observado 20 heterocigotos, el nivel de significancia de la prueba es 0,007. El gradiente de niveles de significancia es bastante basto, como es normal en la prueba exacta de Fisher con muestras pequeñas.
Desafortunadamente, es necesario crear una tabla así para cada experimento, ya que las tablas dependen tanto de n como de p.
Coeficiente de consanguinidad
El coeficiente de consanguinidad F es 1 menos la frecuencia observada de los heterocigotos por encima de la esperada a partir del equilibrio de Hardy-Weinberg.

donde el valor esperado a partir del equilibrio de Hardy-Weinberg se obtiene mediante

Por ejemplo, para los datos de Ford anteriores:


Para dos alelos, la prueba chi-cuadrado sobre la calidad del ajuste con las proporciones de Hardy-Weinberg es equivalente al test de consanguinidad, F = 0.
Historia
La genética mendeliana fue redescubierta en 1900. Sin embargo, se mantuvo su controversia durante varios años, ya que entonces no se sabía cómo podía producir caracteres continuos. Udny Yule (1902) argumentó contra el mendelismo porque pensaba que los alelos dominantes aumentarían en número en una población. El estadounidense William E. Castle (1903) demostró que, sin selección, las frecuencias genotípicas permanecerían estables. Karl Pearson (1903) halló una posición de equilibrio con los valores p = q = 0,5. Reginald Punnet, incapaz de responder al argumento de Yule, le presentó el problema a G. H. Hardy, un matemático británico con el que jugaba al cricket. Hardy era un matemático puro y mostraba cierto desprecio por las matemáticas aplicadas; su opinión sobre el uso que le daban los biólogos a las matemáticas quedó plasmada en un artículo de 1908 en el que lo describe como «muy simple»:
Al Editor de Science: soy reacio a entrometerme en una discusión que concierne a temas de los que no tengo un conocimiento experto, y debería haber esperado que el sencillo argumento que deseo aportar fuera familiar para los biólogos. Sin embargo, ciertas observaciones del señor Udny Yule sobre las que el señor R. C. Punnett ha llamado mi atención sugieren que todavía merece la pena hacerlo...
Supongamos que Aa es un par de caracteres mendelianos, siendo el A dominante, y que en una generación cualquiera el número de dominantes puros (AA), de heterocigotos (Aa) y de recesivos puros (aa) son p:2q:r. Finalmente, supongamos que los números son bastante grandes, de manera que se pueda considerar que el apareamiento es aleatorio, que los sexos están distribuidos uniformemente en las tres variedades y que todas son igualmente fértiles. Es suficiente un poco de matemática del nivel de las tablas de multiplicar para demostrar que en la siguiente generación los números serán (p+q)²:2(p+q)(q+r):(q+r)², o digamos p1:2q1:r1.
La cuestión interesante es: ¿en qué circunstancias será esta distribución la misma que en la generación anterior? Es fácil ver que la condición para esto es q² = pr. Y como q1²=p1r1, para cualquier valor de p, q y r, la distribución permanecerá en cualquier caso sin cambios tras la segunda generación.
Por aquel entonces, el principio se conocía como ley de Hardy en el mundo angloparlante, hasta que Curt Stern (1943) señaló que ya había sido formulado independientemente en 1908 por el físico alemán Wilhelm Weinberg (ver Crow 1999). Otros han intentado asociar el nombre de Castle con la ley por su trabajo de 1903, pero raramente se la alude como ley de Hardy-Weinberg-Castle.
Con la ley de Hardy-Weinberg se asentaron los cimientos de la genética de poblaciones, según la cual, la alteración genética de una población solo puede darse por factores como mutaciones, selección natural, influencias casuales, convergencias o divergencias individuales, de modo que el cambio genético implica la perturbación del equilibrio establecido por la ley de Hardy-Weinberg.
Notas y referencias
Referencias
- Castle, W. E. (1903). The laws of Galton and Mendel and some laws governing race improvement by selection. Proc. Amer. Acad. Arts Sci.. 35: 233–242.
- Crow, J.F. (1999). Hardy, Weinberg and language impediments. Genetics 152: 821-825. enlace
- Emigh, T.H. (1980). A comparison of tests for Hardy-Weinberg equilibrium. Biometrics 36: 627 – 642.
- Ford, E.B. (1971). Ecological Genetics, London.
- Hardy, G. H. (1908). "Mendelian proportions in a mixed population". Science 28: 49–50. copia de la ESP
- Pearson, K. (1903). Mathematical contributions to the theory of evolution. XI. On the influence of natural selection on the variability and correlation of organs. Philosophical Transactions of the Royal Society of London, Ser. A 200: 1–66.
- Stern, C. (1943). "The Hardy–Weinberg law". Science 97: 137–138. Enlace estable del JSTOR
- Weinberg, W. (1908). "Über den Nachweis der Vererbung beim Menschen". Jahreshefte des Vereins für vaterländische Naturkunde in Württemberg 64: 368–382.
- Yule, G. U. (1902). Mendel's laws and their probable relation to intra-racial heredity. New Phytol. 1: 193–207, 222–238.
Notas
- L. L. Cavalli-Sforza / W. F. Bodmer, Genética de poblaciones humanas, Ediciones Omega S. A. Barcelona 1981. ISBN 84-282-0660-0
- «Evolutionibus». Archivado desde el original el 5 de marzo de 2016. Consultado el 2 de julio de 2007.
- Datos de ejemplo de Emigh (1980).
Enlaces externos
- Descripción de poblaciones mendelianas: equilibrio de Hardy-Weinberg
- Calculadora del equilibrio de Hardy-Weinberg
- Simulador de genética de poblaciones
| Control de autoridades |
|
|---|
 Datos: Q49974
Datos: Q49974 Multimedia: Hardy–Weinberg law / Q49974
Multimedia: Hardy–Weinberg law / Q49974