Purga genética
La purga genética es la reducción de la frecuencia de un alelo deletéreo, causada por una mayor eficiencia de la selección natural provocada por la endogamia.[1]
La purga se produce porque muchos alelos deletéreos solo expresan todo su efecto nocivo cuando son homocigotos, presentes en dos copias. Durante la endogamia, a medida que los individuos emparentados se aparean, producen descendientes que tienen más probabilidades de ser homocigotos. Los alelos deletéreos aparecen con más frecuencia, lo que hace que los individuos sean menos aptos genéticamente, es decir, transmiten menos copias de sus genes a las generaciones futuras. Dicho de otra manera, la selección natural purga los alelos deletéreos.
La purga reduce tanto el número total de alelos deletéreos recesivos como la disminución de la aptitud media causada por la endogamia (la depresión endogámica para la aptitud).
El término "purga" se usa a veces para la selección contra alelos deletéreos de una manera general. Evitaría la ambigüedad usar "selección purificadora" en ese contexto general, y reservaría purga a su significado más estricto definido anteriormente.
El mecanismo
Los alelos deletéreos que se segregan en poblaciones de organismos diploides tienen una tendencia notable a ser, al menos, parcialmente recesivos. Esto significa que, cuando ocurren en homocigosis (copias dobles), reducen la aptitud en más del doble que cuando ocurren en heterocigosis (copia única). En otras palabras, parte de su potencial efecto deletéreo está oculto en la heterocigosis, pero se expresa en la homocigosis, por lo que la selección es más eficaz contra ellos cuando ocurren en la homocigosis. Dado que la endogamia aumenta la probabilidad de ser homocigoto, aumenta la fracción del efecto deletéreo potencial que se expresa y, por lo tanto, se expone a la selección. Esto provoca cierto aumento de la presión selectiva contra los alelos deletéreos (parcialmente) recesivos, lo que se conoce como purga. Por supuesto, también causa cierta reducción en la aptitud, lo que se conoce como depresión endogámica.
La purga puede reducir la frecuencia promedio de alelos deletéreos en el genoma por debajo del valor esperado en una población no endogámica.[2] Aunque esta reducción no suele compensar todos los efectos negativos de la endogamia,[3] tiene varias consecuencias beneficiosas para la aptitud. Una consecuencia es la reducción de la llamada carga de consanguinidad. Esto significa que, después de la purga, se espera que la endogamia adicional sea menos dañina. Pero la consecuencia más inmediata es la reducción de la depresión endogámica de la aptitud: debido a la purga, la aptitud media disminuye menos de lo que se esperaría solo por la endogamia y, después de una disminución inicial, incluso puede recuperarse hasta casi su valor antes de la endogamia.[4]
El efecto conjunto de la endogamia y la purga sobre la aptitud
Tener en cuenta la purga cuando se predice la depresión por endogamia es importante en la genética evolutiva, porque la disminución de la aptitud causada por la endogamia puede ser determinante en la evolución de la diploidía, la reproducción sexual y otras características biológicas principales. También es importante en la cría de animales y, por supuesto, en la genética de la conservación, porque la depresión endogámica puede ser un factor relevante que determina el riesgo de extinción de las poblaciones en peligro, y porque los programas de conservación pueden permitir cierto manejo de la reproducción para controlar la endogamia.[5]
En resumen, debido a la purga, la depresión por consanguinidad no es proporcional a la medida estándar de consanguinidad (coeficiente de consanguinidad F de Wright), ya que esta medida solo se aplica a los alelos neutrales. En cambio, la disminución de la aptitud es proporcional a la "endogamia purgada" g, lo que da la probabilidad de ser homocigoto para los alelos deletéreos debido a la endogamia, teniendo en cuenta cómo se purgan.
La purga reduce la depresión endogámica de dos formas: primero, ralentiza su progreso; en segundo lugar, reduce la depresión endogámica general que se espera a largo plazo. Cuanto más lento es el progreso de la endogamia, más eficiente es la purga.
Una explicación más detallada
En ausencia de selección natural, se esperaría que la aptitud media disminuya exponencialmente a medida que aumenta la consanguinidad, donde la consanguinidad se mide utilizando el coeficiente de consanguinidad de Wright, F[6] (la razón por la que la disminución es exponencial en F lugar de lineal es solo que la aptitud generalmente se considera un rasgo multiplicativo). La tasa a la que la aptitud disminuye a medida que aumenta F (la tasa de depresión por endogamia δ) depende de las frecuencias y los efectos deletéreos de los alelos presentes en la población antes de la endogamia.
El coeficiente F anterior es la medida estándar de consanguinidad y da la probabilidad de que, en cualquier locus neutral dado, un individuo haya heredado dos copias de un mismo gen de un ancestro común (es decir, la probabilidad de ser homocigoto "por descendencia"). En condiciones simples, F se puede calcular fácilmente en términos de tamaño de la población o de información genealógica. F a menudo se denota usando minúsculas (f), pero no debe confundirse con el coeficiente de coancestría.
Sin embargo, la predicción anterior para la disminución de la aptitud rara vez se aplica, ya que se derivó asumiendo que no hay selección, y la aptitud es precisamente el rasgo objetivo de la selección natural. Por lo tanto, el coeficiente de consanguinidad F Wright para loci neutros no se aplica a los alelos deletéreos, a menos que la consanguinidad aumente tan rápido que el cambio en la frecuencia de los genes se rija solo por muestreo aleatorio (es decir, por deriva genética).
Por lo tanto, la disminución de la aptitud debe predecirse usando, en lugar del coeficiente de consanguinidad estándar F, un "coeficiente de consanguinidad purgado" (g) que da la probabilidad de ser homocigoto por descendencia para alelos deletéreos (parcialmente) recesivos, teniendo en cuenta cómo su la frecuencia se reduce mediante la purga.[4] Debido a la purga, la aptitud disminuye al mismo ritmo δ que en ausencia de selección, pero en función de g lugar de F
Este coeficiente consanguinidad purgado g también se puede calcular, con una buena aproximación, utilizando expresiones simples en términos del tamaño de la población o de la genealogía de los individuos. Sin embargo, esto requiere cierta información sobre la magnitud de los efectos deletéreos que están ocultos en la condición heterocigótica, pero que se expresan en la homocigosis. Cuanto mayor sea esta magnitud, denominada coeficiente de purga d, más eficiente será el purgado.
Una propiedad interesante de la purga es que, durante la consanguinidad, mientras F aumenta acercándose a un valor final F = 1, g puede acercarse a un valor final mucho menor. Por lo tanto, no se trata solo de que la purga ralentice el deterioro de la aptitud, sino también de que reduce la pérdida de aptitud general producida por la endogamia a largo plazo.
La purga es menos eficaz contra alelos levemente deletéreos que contra los letales, pero en general, cuanto más lento es el aumento de la endogamia F, menor se vuelve el valor final del coeficiente de endogamia purgado g, por lo tanto, la reducción final de la aptitud. Esto implica que, si la consanguinidad progresa con la suficiente lentitud, no se espera una depresión consanguínea relevante a largo plazo. Esto da como resultado que la aptitud de una población pequeña, que ha sido una población pequeña durante mucho tiempo, puede ser la misma que una población grande con más diversidad genética. En genética de conservación, sería muy útil determinar la tasa máxima de aumento de la endogamia que permite una purga tan eficiente.
Ejemplos
Ecuaciones predictivas cuando la endogamia se debe al pequeño tamaño de la población
Considere una gran población no consanguínea con aptitud media W . Entonces, el tamaño de la población se reduce a un nuevo valor más pequeño N (de hecho, el tamaño efectivo de la población debería usarse aquí), lo que lleva a un aumento progresivo de la endogamia.
Luego, la depresión por endogamia ocurre a una tasa δ, debido a alelos deletéreos (parcialmente) recesivos que estaban presentes a bajas frecuencias en diferentes loci. Esto significa que, en ausencia de selección, el valor esperado para la aptitud media después de t generaciones de consanguinidad sería:

donde es la media poblacional del coeficiente de endogamia de Wright después de t generaciones de endogamia.[6]

Sin embargo, dado que la selección opera sobre la aptitud, la aptitud media debe predecirse teniendo en cuenta tanto la endogamia como la purga, como

En la ecuación anterior, es el "coeficiente de consanguinidad purgado" medio después de t generaciones de consanguinidad.[4] Depende del "coeficiente de purga" d, que representa los efectos deletéreos ocultos en la heterocigosis pero expuestos en la homocigosis.

El "coeficiente de consanguinidad purgado" promedio puede aproximarse usando la expresión recurrente
![{\displaystyle g_{t}=\left[\left(1-{\frac {1}{2N}}\right)g_{t-1}+{\frac {1}{2N}}\right]\left[1-2dF_{t-1}\right]}](../I/79f5343c6d07102ddc3f1066b12a9880094ce9c7.svg)
También hay ecuaciones predictivas para usar con información genealógica.
El ejemplo de depresión endogámica debida a letales recesivos
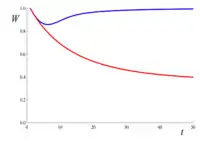
Como ejemplo de depuración genética, considérese una gran población en la que hay alelos letales recesivos que se segregan con una frecuencia muy baja en muchos loci, de modo que cada individuo porta en promedio uno de estos alelos. Aunque alrededor del 63% de los individuos portan al menos uno de estos alelos letales, casi ninguno lleva dos copias del mismo letal. Por lo tanto, dado que los letales se consideran completamente recesivos (es decir, son inofensivos en heterocigosis), casi no causan muertes. Ahora suponga que el tamaño de la población se reduce a un valor pequeño (digamos N = 10) y permanece así de pequeño durante muchas generaciones. A medida que aumenta la consanguinidad, la probabilidad de ser homocigoto para uno (o más) de estos alelos letales también aumenta, lo que hace que disminuya la aptitud. Sin embargo, a medida que esos letales comienzan a ocurrir en homocigosis, la selección natural comienza a purgarlos. La figura de la derecha da la disminución esperada de la aptitud contra el número de generaciones, teniendo en cuenta solo el aumento en la endogamia F (línea roja), o tanto la endogamia como la purga (línea azul, calculada utilizando el coeficiente de endogamia purgado g). Este ejemplo muestra que la purga puede ser muy eficaz para prevenir la depresión por endogamia. Sin embargo, para los alelos deletéreos no letales, la eficiencia de la purga sería menor y puede requerir poblaciones más grandes para superar la deriva genética.
Renovación del genoma
Las levaduras Saccharomyces cerevisiae y Saccharomyces paradoxus tienen un ciclo de vida que alterna entre períodos largos de reproducción asexual como un diploide, que terminan en la meiosis que por lo general es seguida inmediatamente autofecundación, con solo el raro cruzamiento.[7] Las mutaciones deletéreas recesivas se acumulan durante la fase de expansión diploide y se purgan durante la autofecundación: esta purga se ha denominado "renovación del genoma".[8][9]
Evidencia y problemas
Cuando una población previamente estable sufre endogamia, si nada más cambia, la selección natural debería consistir principalmente en purgas. Las consecuencias conjuntas de la endogamia y la depuración sobre la aptitud varían en función de muchos factores: la historia previa de la población, la tasa de aumento de la endogamia, la dureza del medio o de las condiciones competitivas, etc. Los efectos de la purga fueron notados por primera vez por Darwin[10] en plantas, y se han detectado en experimentos de laboratorio y en poblaciones de vertebrados que experimentan endogamia en zoológicos o en la naturaleza, así como en humanos.[11] La detección de purga a menudo se ve oscurecida por muchos factores, pero existe evidencia consistente de que, de acuerdo con las predicciones explicadas anteriormente, la endogamia lenta da como resultado una purga más eficiente, de modo que una F endogámica dada conduce a una menor amenaza para la viabilidad de la población si se ha producido más lentamente.[12]
Sin embargo, en situaciones prácticas, el cambio genético en la aptitud también depende de muchos otros factores, además de la endogamia y la purga. Por ejemplo, la adaptación a las condiciones ambientales cambiantes a menudo provoca cambios genéticos importantes durante la endogamia. Además, si la endogamia se debe a una reducción en el tamaño de la población, la selección contra nuevas mutaciones deletéreas puede volverse menos eficiente y esto puede inducir una disminución adicional de la aptitud a medio-largo plazo.
Además, parte de la depresión endogámica no podría deberse a alelos deletéreos, sino a una ventaja intrínseca de ser heterocigoto en comparación con ser homocigoto para cualquier alelo disponible, lo que se conoce como sobredominio. La depresión por consanguinidad causada por sobredominio no se puede purgar, pero parece ser una causa menor de depresión por consanguinidad general, aunque su importancia real sigue siendo un tema de debate.[13]
Por lo tanto, predecir la evolución real de la aptitud durante la endogamia es muy difícil de alcanzar. Sin embargo, el componente de disminución de la aptitud que se espera de la endogamia y la purga de alelos deletéreos podría predecirse usando g.
Referencias
- García-Dorado, A. (2015-09). «On the consequences of ignoring purging on genetic recommendations for minimum viable population rules». Heredity 115 (3): 185-187. ISSN 1365-2540. PMC 4814235. PMID 25873145. doi:10.1038/hdy.2015.28.
- Glémin, Sylvain (2003-12). «How are deleterious mutations purged? Drift versus nonrandom mating». Evolution; International Journal of Organic Evolution 57 (12): 2678-2687. ISSN 0014-3820. PMID 14761049. doi:10.1111/j.0014-3820.2003.tb01512.x.
- Hedrick, Philip W.; Kalinowski, Steven T. (1 de noviembre de 2000). «Inbreeding Depression in Conservation Biology». Annual Review of Ecology and Systematics 31 (1): 139-162. ISSN 0066-4162. doi:10.1146/annurev.ecolsys.31.1.139.
- García-Dorado, Aurora (2012-4). «Understanding and Predicting the Fitness Decline of Shrunk Populations: Inbreeding, Purging, Mutation, and Standard Selection». Genetics 190 (4): 1461-1476. ISSN 0016-6731. PMC 3316656. PMID 22298709. doi:10.1534/genetics.111.135541.
- Frankham, Richard (1 de noviembre de 2005). «Genetics and extinction». Biological Conservation (en inglés) 126 (2): 131-140. ISSN 0006-3207. doi:10.1016/j.biocon.2005.05.002.
- Crow, J.F.; Kimura, M. (1970). An Introduction to Population Genetics Theory. NY: Harper & Row.
- Tsai, Isheng J.; Bensasson, Douda; Burt, Austin; Koufopanou, Vassiliki (25 de marzo de 2008). «Population genomics of the wild yeast Saccharomyces paradoxus: Quantifying the life cycle». Proceedings of the National Academy of Sciences of the United States of America 105 (12): 4957-4962. ISSN 1091-6490. PMC 2290798. PMID 18344325. doi:10.1073/pnas.0707314105.
- Mortimer, R. K.; Romano, P.; Suzzi, G.; Polsinelli, M. (1994-12). «Genome renewal: a new phenomenon revealed from a genetic study of 43 strains of Saccharomyces cerevisiae derived from natural fermentation of grape musts». Yeast (Chichester, England) 10 (12): 1543-1552. ISSN 0749-503X. PMID 7725789. doi:10.1002/yea.320101203.
- Masel, Joanna; Lyttle, David N. (2011-12). «The consequences of rare sexual reproduction by means of selfing in an otherwise clonally reproducing species». Theoretical Population Biology 80 (4): 317-322. ISSN 1096-0325. PMC 3218209. PMID 21888925. doi:10.1016/j.tpb.2011.08.004.
- Darwin, C. R. (1876). The effects of cross and selffertilisation in the vegetable kingdom. London: John Murray.
- Crnokrak, Peter; Barrett, Spencer C. H. (2002-12). «Perspective: purging the genetic load: a review of the experimental evidence». Evolution; International Journal of Organic Evolution 56 (12): 2347-2358. ISSN 0014-3820. PMID 12583575. doi:10.1111/j.0014-3820.2002.tb00160.x.
- Leberg, Paul L.; Firmin, Brigette D. (2008-01). «Role of inbreeding depression and purging in captive breeding and restoration programmes». Molecular Ecology 17 (1): 334-343. ISSN 0962-1083. PMID 18173505. doi:10.1111/j.1365-294X.2007.03433.x.
- Crow, James F. (2008). «Mid-century controversies in population genetics». Annual Review of Genetics 42: 1-16. ISSN 0066-4197. PMID 18652542. doi:10.1146/annurev.genet.42.110807.091612.
| Control de autoridades |
|
|---|
 Datos: Q2118830
Datos: Q2118830