Bilateria
Los bilaterales (Bilateria) son los animales con simetría bilateral, por la cual el organismo es simétrico respecto a un plano (plano sagital) que divide el cuerpo en dos mitades especularmente idénticas. Este tipo de simetría es el más extendido en el reino animal. La simetría bilateral se relaciona con la motilidad. Todos los filos triblásticos presentan simetría bilateral, aunque secundariamente puedan perderla en estado adulto, como sucede en los equinodermos.
| Bilaterales | ||
|---|---|---|
| Rango temporal: Ediacárico – Reciente | ||
 | ||
 | ||
| Taxonomía | ||
| Superreino: | Eukaryota | |
| Reino: | Animalia | |
| Subreino: | Eumetazoa | |
| (sin rango) | ParaHoxozoa | |
| (sin rango): |
Bilateria Haeckel, 1874 | |
| Linajes | ||
| ||
| Sinonimia | ||
| ||
Características
La parte del cuerpo que toca primero el entorno (extremo anterior) lleva la mayor parte de los órganos de los sentidos y es diferente del extremo posterior; los dos lados del cuerpo, que se relacionan con el entorno de la misma forma, son idénticos, pero las superficies superior e inferior, por el contrario, son diferentes. Por tanto, un extremo está dirigido hacia delante y una superficie está orientada hacia el substrato. Este fenómeno comporta la cefalización y representa la aparición del eje antero-posterior del cuerpo.
Solo los acelomados (platelmintos, nemertinos y grupos menores) tienen una organización corporal de tipo macizo en la que el espacio comprendido entre la capa muscular de la pared corporal y el intestino está lleno de mesénquima y fibras musculares. Todos los demás animales bilaterales poseen un espacio repleto de líquido llamado cavidad general entre la pared el cuerpo y los órganos internos o, si falta, derivan claramente de formas que sí lo poseen. Dicho líquido puede actuar como esqueleto hidrostático y puede intervenir también en la translocación de gases, nutrientes y sustancias residuales.
En la inmensa mayoría de animales tal cavidad es de tipo celomado, es decir, poseen un celoma como cavidad corporal (anélidos, moluscos, artrópodos, cordados, etc.); De estos, pueden divergir dos ramas, según sean enterocelomados o esquizocelomados. Un pequeño número posee, por el contrario, un pseudoceloma (o blastoceloma) y reciben el nombre de pseudocelomados (o blastocelomados) (nematodos, rotíferos, etc.). El pseudoceloma representa un blastocele embrionario persistente.
Origen y filogenia
Se considera que el primer bilateral es también el primer gusano; de tal modo que el antiguo taxón Vermes puede considerarse parafilético, ya que de un gusano como último común ancestro descienden todos los animales complejos tales como moluscos, artrópodos, equinodermos o vertebrados. Se presume que este ancestro adquiere la simetría bilateral, la condición triblástica del embrión y posee una sola abertura corporal.
Por mucho tiempo se creyó que el grupo basal de Bilateria fue Acoelomata y que de él habrían derivado celomados y pseudocelomados. Sin embargo, el análisis filogenómico moderno demostró que todos estos grupos son artificiales (polifiléticos). Actualmente según estudios moleculares de especies de evolución lenta y con métodos estadísticos mejorados para poder evitar el error sistemático han dado la siguiente filogenia posible:[1][2][3][4][5][6][7]















Un árbol filogenético alternativo
Un árbol filogenético alternativo publicado en el libro "The Invertebrate Tree Of Life" (2020) es el propuesto por Gonzalo Giribet. Los nodos marcados son defendidos por el autor:[8]
| Bilateria |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Teorías anteriores
No hay muchas evidencias de la transición del nivel de organización diblástico al triblástico. Dos teorías han polarizado las ideas de los zoólogos en los últimos años; a dichas hipótesis hay que añadir una tercera de reciente publicación.
Teoría planuloide/aceloide
La mayoría de los autores son partidarios de la teoría planuloide/aceloide que sostiene un origen acelomado de todos los bilaterales, con lo que los gusanos planos (platelmintos) aparecerían antes que los celomados. Entre los trabajos recientes en este sentido cabe destacar el de Zrzavý et al.,[9] el primero basado en un estudio simultáneo de datos morfológicos y moleculares ("total evidence"). Esta teoría vincula el incremento de la complejidad estructural con el aumento de las dimensiones corporales y con la colonización de nuevos hábitats. Las relaciones entre los grandes grupos de metazoos, bajo el prisma de esta teoría, tienen dos interpretaciones:
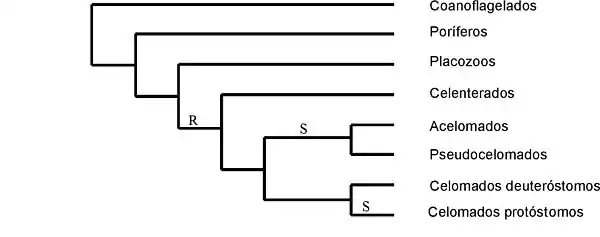
- a) suponiendo que la segmentación espiral del huevo (S) evolucionó una sola vez, pero la segmentación radial (R) es difilética, es decir, no es homóloga en Diblásticos y Deuteróstomos; este esquema evolutivo refleja en gran medida las ideas de Hyman[10] y está ampliamente aceptado por los zoólogos y se ve apoyado por resultados de tipo molecular; indica que las cavidades corporales evolucionaron siguiendo el orden acelomados, pseudocelomados, celomados:

- Análisis genéticos llevados a cabo en 2009 por un equipo de investigación internacional sobre 350 especies de acelomorfos parece confirmar esta hipótesis.[11][12]
- b) considerando la segmentación radial como monofilética y que la segmentación espiral evolucionó dos veces de modo independiente, una en el clado acelomados y pseudocelomados, que formarían un grupo monofilético, y otra en celomados protósomos; aunque lógicamente posible, este árbol no está difundido entre los zoólogos:
Teoría del arquicelomado
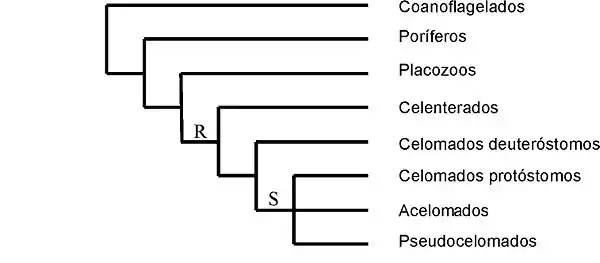
Este punto de vista se aproxima al de la escuela de Remane.[13] La teoría postula que todos los triblásticos son descendientes de un antecesor celomado. En los gusanos planos se habría producido, pues, una pérdida de la complejidad estructural, es decir, pérdida secundaria del celoma y del ano. Esta hipótesis no es tan popular como la anterior, pero está ganando soporte procedente de la ultraestructura comparada y de ciertos estudios recientes basados en datos moleculares, como por ejemplo el de Balavoine.[14] No obstante, se hace difícil explicar desde un punto de vista funcional el desarrollo tan precoz de las cavidades celomáticas en el curso de la evolución de los metazoos. Las interrelaciones de los metazoos según esta teoría serían las mostradas en el cladograma siguiente; es la hipótesis más parsimoniosa en cuanto a la segmentación del huevo:
Teoría colonial
Dewel[15] ha propuesto una novedosa hipótesis sobre el origen de los triblásticos que suscita vivas polémicas. Dicha teoría admite la transición colonial del nivel unicelular al nivel porífero, y postula dos nuevas transiciones, también de tipo colonial, la de porífero a cnidario y la de cnidario a bilateral. En el primer caso, una colonia de organismos con un nivel de organización porífero sufrió un proceso de individuación para originar una nueva entidad con nivel de organización cnidario; en esta transición se consideran dos eventos cruciales, (1) la pérdida de la circulación del agua a través del cuerpo en las Esponjas, con el consiguiente establecimiento de una sola abertura y (2) la aparición de epitelio auténtico en cnidarios. En el modelo para la transición de cnidario a bilateral, el último ancestro común de los eumetazoos se considera que tuvo una organización tipo cnidario colonial (similar al de los antozoos pennatuláceos); dicha colonia originó por duplicación de sus componente y posterior individuación una colonia con nivel de organización bilateral. Así, la fauna de Ediacara, los arqueociatos y otros grupos problemáticos serían el reflejo de que la complejidad se originó temprano en la evolución de los metazoos.
Urbilateria
Cabe destacar que una de las paradojas más notables sobre el origen de los bilaterales es que, según los datos morfológicos y ontogénicos, el último antecesor común de protóstomos y deuteróstomos, el llamado Urbilateria por De Robertis y Sasai,[16] era ya complejo (cabeza con fotorreceptores, tronco segmentado con «apéndices», cola, cerebro dorsal, cavidades celomáticas y gónadas seriadas con gonoducto, sistema circulatorio, corazón), lo cual no tiene una explicación clara a la luz de las hipótesis actuales. La hipótesis colonial proporcionaría una explicación a la complejidad del primer bilateral (ya que procedería de una colonia de Cnidarios formada por zooides con diversificación de funciones) y de la rápida diversificación de los bilaterales en el Cámbrico (explosión cámbrica). Durante dicha radiación evolutiva, pérdidas y simplificaciones selectivas pudieron ser una fuerza creativa de innovación. Finalmente, la individuación de la colonia pudo ser un evento único. La autora de la teoría deja implícito que los celomados son anteriores al resto de bilaterales, con lo que su punto de vista se acerca al de Remane, aunque por una vía totalmente diferente.
Véase también
- Simetría bilateral
- Anexo:Animales bilaterales
- Centroneuralia: Clado propuesto que reúne a cordados y protostomados como hermanos de Xenambulacraria.
Referencias
- Marlétaz, Ferdinand; Peijnenburg, Katja T. C. A.; Goto, Taichiro; Satoh, Noriyuki; Rokhsar, Daniel S. (2019). «A new spiralian phylogeny places the enigmatic arrow worms among gnathiferans». Current Biology 29 (2): 312-318.e3. doi:10.1016/j.cub.2018.11.042.
- Ecdysozoa description
- Lahcen I. Campbell, Omar Rota-Stabelli, Gregory D. Edgecombe, Trevor Marchioro, Stuart J. Longhorn, Maximilian J. Telford, Hervé Philippe, Lorena Rebecchi, Kevin J. Peterson, and Davide Pisani (2011). MicroRNAs and phylogenomics resolve the relationships of Tardigrada and suggest that velvet worms are the sister group of Arthropoda Archivado el 19 de febrero de 2020 en Wayback Machine.. PNAS. https://doi.org/10.1073/pnas.1105499108.
- Philippe, Hervé; Poustka, Albert J.; Chiodin, Marta; Hoff, Katharina J.; Dessimoz, Christophe; Tomiczek, Bartlomiej; Schiffer, Philipp H.; Müller, Steven; Domman, Daryl; Horn, Matthias; Kuhl, Heiner; Timmermann, Bernd; Satoh, Noriyuki; Hikosaka-Katayama, Tomoe; Nakano, Hiroaki; Rowe, Matthew L.; Elphick, Maurice R.; Thomas-Chollier, Morgane; Hankeln, Thomas; Mertes, Florian; Wallberg, Andreas; Rast, Jonathan P.; Copley, Richard R.; Martinez, Pedro; Telford, Maximilian J. (2019). «Mitigating Anticipated Effects of Systematic Errors Supports Sister-Group Relationship between Xenacoelomorpha and Ambulacraria». Current Biology 29 (11): 1818-1826.e6. ISSN 0960-9822. PMID 31104936. doi:10.1016/j.cub.2019.04.009. hdl:21.11116/0000-0004-DC4B-1.
- Marlétaz, Ferdinand (17 de junio de 2019). «Zoology: Worming into the Origin of Bilaterians». Current Biology (en inglés) 29 (12): R577-R579. ISSN 0960-9822. PMID 31211978. doi:10.1016/j.cub.2019.05.006.
- Telford, Maximilian J.; Robertson, Helen E.; Schiffer, Philipp H. (18 de junio de 2018). «Orthonectids Are Highly Degenerate Annelid Worms». Current Biology (en inglés) 28 (12): 1970-1974.e3. ISSN 0960-9822. PMID 29861137. doi:10.1016/j.cub.2018.04.088.
- Helen E. Robertson, Philippe Hervé, Maximilian; J. Telford, Poustka, Albert J.; Chiodin, Marta; Hoff, Katharina J.; Dessimoz, Christophe; Tomiczek (2019) The mitochondrial genomes of the mesozoans Intoshia linei, Dicyema sp. and Dicyema japonicum. PDF Cambridge University.
- Giribet et Edgecombe.The Invertebrate Tree Of Life. Princeton University Press, 2020. ISBN 06911702589780691170251. pag 21.
- Zrzavý, J., Mihulka, S., Kepka, P., Bezdék, A. y Tietz, D. (1998). «Phylogeny of Metazoa based on morphological and 18S ribosomal DNA evidence». Cladistics, 14(3): 249-285
- Hyman, L. H. (1949-67). The Invertebrates. Vols. 1-6. McGraw-Hill, New York.
- Andreas Hejnol et alii. (16 de septiembre de 2009). «Assessing the root of bilaterian animals with scalable phylogenomic methods». Proceedings of the Royal Society (en inglés). Royal Society Publishing. Consultado el 11 de julio de 2010.
- CORDIS noticias (28 de septiembre de 2009). «Unos gusanos esclarecen etapas primitivas de la evolución animal» (periodístico). Consultado el 11 de julio de 2010.
- Remane, A., Storcu, V. & Welsch, U., 1980. Zoología sistemática. Clasificación del Reino Animal. Omega, Barcelona.
- Balavoine, G. (1998). «Are Platyhelminthes coelomates without a coelom? An argument based on the evolution of Hox genes». Amer. Zool., 38(6): 843-858
- Dewel, R. A. (2000). «Colonial origin of Eumetazoa: Major morphological transi-tions and the origin of bilaterian complexity». J. Morphol., 243: 35-74
- Robertis, E. M. de y Sasai, Y. (1996). «A common plan for dorsoventral patterning in Bilateria». Nature, 380: 37-40 doi 10.1038/380037a0
Obras generales
- Altaba, C. R. et al. (1991). Invertebrats no artròpodes. Història Natural dels Països Catalans, 8. Enciclopèdia Catalana, S. A., Barcelona, 598 pp. ISBN 84-7739-177-7
- Hickman, C. P.; Ober, W. C. y Garrison, C. W. (2006). Principios integrales de zoología, 13.ª. edición. McGraw-Hill-Interamericana, Madrid (etc.), XVIII+1022 pp. ISBN 84-481-4528-3
Enlaces externos
- Para más información y cladogramas alternativos y/o más detallados véase Tree of Life