Aelurosaurus
Aelurosaurus (nom signifiant "lézard chat", du grec ancien αἴλουρος, "chat", et σαῦρος, "lézard") est un genre éteint de petits thérapsides gorgonopsiens carnivores ayant vécu du milieu à la fin du Permien dans l'actuelle Afrique du Sud. Il a été découvert dans le Bassin du Karoo en Afrique du Sud et décrit pour la première fois par le paléontologue Richard Owen en 1881. Owen l'a nommé Aelurosaurus car l'animal lui paraissait être un ancêtre des marsupiaux aux allures de chats, mais sans être encore un mammifère lui-même. Le genre Aelurosaurus contient cinq espèces : A. felinus, A. whaitsi, A. polyodon, A. wilmanae, et A. ? watermeyeri[1]. L'espèce type, A. felinus, est généralement décrite avec des traits bien établis, tandis que les quatre autres ne le sont pas en raison du mauvais état de conservation de leurs fossiles holotypes.
| Règne | Animalia |
|---|---|
| Embranchement | Chordata |
| Sous-embr. | Vertebrata |
| Classe | Synapsida |
| Ordre | Therapsida |
| Sous-ordre | † Gorgonopsia |
Découverte
Aelurosaurus a été découvert en 1881 dans le Bassin du Karoo en Afrique du Sud. Le crâne holotype n'est pas complet : il lui manque la région postorbitaire, ce qui conduisit Richard Owen à en décrire seulement la partie antérieure. Après avoir examiné les sutures et les dents, et avoir observé qu'elles n'étaient pas en bon état, il en conclut que le crâne appartenait à un spécimen adulte, voire âgé[1].
Espèces
Aelurosaurus felinus

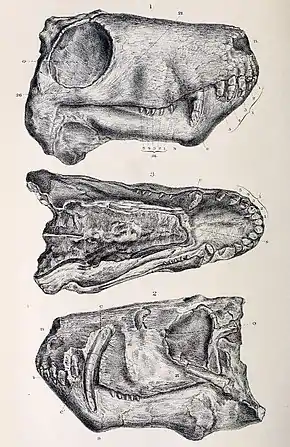
Le crâne holotype de l'espèce Aelurosaurus felinus, décrit par Owen en 1881, possède une orbite en forme d'ellipse mesurant 2,5 cm de largeur de 3 cm de hauteur ; la narine mesure 1,3 cm de large et 0,9 cm de long. La mâchoire supérieure a une largeur maximale d'environ 3,5 cm juste en-dessous des orbites. La mandibule, longue d'un peu plus de 7,5 cm et large de 2 cm, fait pratiquement longueur de tout le crâne. Ce spécimen en particulier a été préservé avec la gueule fermée, les dents du haut couvrant entièrement les dents de la mandibule. Sur la mâchoire supérieure, il y a cinq incisives, un espace vide d'environ 0,8 cm, une canine (1,2 cm exposés, sur une longueur totale d'environ 3,6 cm), un autre espace vide d'environ 0,8 cm, et cinq molaires. Une fois qu'Owen a eu retiré le mur alvéolaire de la canine maxillaire supérieure, il a découvert la canine mandibulaire, qu'il a trouvée à peu près de la même taille que la maxillaire[1]. Le canal semi-circulaire qui s'observe chez A. felinus et chez d'autres gorgonopsiens suggère que leurs têtes étaient inclinées vers le ventre : c'est une caractéristique des prédateurs[2].
Owen avait supposé à l'origine qu'il avait affaire à un crâne d'animal adulte, mais d'autres paléontologues ont estimé depuis qu'il pouvait s'agir du crâne d'un juvénile. Owen ne déduisait son âge qu'à partir de l'état de ses sutures et de ses dents, mais d'autres chercheurs ont remarqué d'autres indices pouvant indiquer un jeune âge, notamment ses « museau court, orbites larges, dents postcanines élancées, remplacement dentaire, nombreuses petites dents postcanines, arches crâniennes élancées, vomer étroit, tubérosités palatales bien développées, dents sur les apophyses transversales, grandes ectoptérygoïdes, mandibule élancée, [et] symphysis ouverte »[3],[4].
A. whaitsi
A. whaitsi a été décrit pour la première fois par Robert Broom en 1911. L'holotype de A. whaitsi est un autre crâne, mais dont est surtout conservé le museau, son indicateur principal consistant en un symphysis plus plat. La classification de ce crâne a changé plusieurs fois au fil du XXe siècle, car il avait très peu de traits distinctifs. et tous pouvaient n'être que des signes que l'on avait affaire à un individu juvénile ou au contraire à un spécimen appartenant à un autre genre qu'Aelurosaurus[3].
A. polyodon
A. Polyodon a été décrit pour la première fois par Broom en 1935. L'holotype de cette espèce est un museau écrasé. Nommé à l'origine Galerhinus polyodon, il a été renommé Aelurosaurus polyodon en 1970[3].
A. wilmanae
A. wilmanae a été décrit pour la première fois par Broom en 1940. L'holotype de A. wilmanae est un crâne, le mieux préservé dont on dispose pour le genre Aelurosaurus à l'heure actuelle. Le trait définitoire de l'espèce A. wilmanae est un museau plus large comparé à celui d'A. felinus, mais globalement le crâne reste petit : seulement 11 cm de long[3].
A.? watermeyeri
A.? watermeyeri a été décrit pour la première fois par Broom en 1940. L'holotype est un crâne. Il avait été nommé à l'origine Scylacocephalus watermeyeri. Le genre auquel appartient ce spéciment fait toujours débat parmi les paléontologues, car il partage des traits avec Aelurosaurus (une orbite large et un museau court, notamment) et avec Aloposaurus (un prépariétal et un postfrontal plus grands)[3].
Géologie et paléoenvironnement
Aelurosaurus felinus a été découvert à l'origine dans le Gough, qui fait partie du Bassin du Karoo en Afrique du Sud, qui inclut la Zone d'assemblage Pristerognathus et la Zone d'assemblage Tapinocephalus (deux aires définies à la fois par leurs caractéristiques géologiques et par les types de fossiles qu'on y a retrouvés)[5],[6]. Ces Zones d'assemblage font partie du groupe de Beaufort et s'étendent depuis le milieu jusqu'à la fin du Permien[7]. La composition stratigraphique du groupe de Beaufort inclut de la siltstone, de la mudstone, et des lits de grès, contenant des preuves que des cours d'eau (rivières ou fleuves) ont eu leur lit sur ce terrain, dans des conditions climatiques semi-arides. Des coprolithes denses contenant des os ont été découverts dans ces zones, ce qui indique que des prédateurs ont vécu près de ces cours d'eau[8].
Bien que la plupart des gorgonopsiens comme Aelurosaurus aient été découverts dans le Bassin du Karoo en Afrique du Sud, d'autres gorgonopsiens ont été découverts plus tard dans la Zone d'assemblage de Vyazniki, en Russie[9].
Notes et références
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Aelurosaurus » (voir la liste des auteurs).
- Owen (1881).
- Araújo, R., V. Fernandez, M. J. Polcyn, J. Fröbisch, R. M. S. Martins (2017).
- Gebauer (2007).
- « ...short snout, large orbits, slender postcanine teeth, tooth replacement, numerous small postcanine teeth, well developed foramina, large supraorbital portion of the frontal, anteriorly situated preparietal, slender skull arches, narrow vomer, well developed palatal tuberosities, teeth on transverse apophyses, large ectopterygoids, slender mandible, [and] open symphysis. »
- Boonstra (1935).
- Kammerer, C.F., R.M.H. Smith, M. O. Day, B.S. Rubidge (2015).
- Angielczyk, K. D., B. S. Rubidge, M. O. Day, F. Lin. (2016).
- Smith, R. M. H., J. Botha-Brink (2011).
- Kammerer, C.F. (2016).
Bibliographie
- (en) Angielczyk, K. D., B. S. Rubidge, M. O. Day, F. Lin, « A reevaluation of Brachyprosopus broomi and Chelydontops altidentalis, dicynodonts (Therapsida, Anomodontia) from the middle Permian Tapinocephalus Assemblage Zone of the Karoo Basin, South Africa », Journal of Vertebrate Paleontology, 2016, vol. 36, no 2. DOI:10.1080/02724634.2016.1078342
- (en) Araújo, R., V. Fernandez, M. J. Polcyn, J. Fröbisch, R. M. S. Martins, « Aspects of gorgonopsian paleobiology and evolution: insights from the basicranium, occiput, osseous labyrinth, vasculature, and neuroanatomy », PeerJ., 2017, vol. 5, p. 1-45.
- (en) L. D. Boonstra, « On the South African Gorgonopsian reptiles preserved in the American Museum of Natural History », American Museum Novitates, 1935, no 772, p. 1-14.
- (en) Gebauer, E.V.I., Phylogeny and evolution of the Gorgonopsia with a special reference to the skull and skeleton of GPIT/RE/7113 ("Aelurognathus"? parringtoni), thèse, Dissertation Universität Tübingen, 2007. [lire en ligne]
- (en) Kammerer, C.F., R.M.H. Smith, M. O. Day, B.S. Rubidge, « New information on the morphology and stratigraphic range of the mid-Permian gorgonopsian Eriphostoma microdon », Papers in Palaeontology, 2015, vol. 1, no 2, p. 201-221.
- (en) Kammerer, C.F., « On the evolutionary history of the Gorgonospia, with new information on their global first and last appearances », International Geological Congress, vol. 36, 2016. [lire en ligne]
- (en) Richard Owen, On the order Theriodontia with a description of a new genus and species (Aelurognathus fel. Ow.), Quat. Jour. Geol. Soc. London, 1881, vol. 37, p. 261-265.
- (en) Smith, R. M. H., J. Botha-Brink, « Morphology and composition of bone-bearing coprolites from the Late Permian Beaufort Group, Karoo Basin, South Africa », Palaeogeography, Palaeoclimatology, Palaeoecology, 2011, vol. 312, p. 40-53.
Liens externes
 Portail de la paléontologie
Portail de la paléontologie  Portail de l’herpétologie
Portail de l’herpétologie