Allostérie
L'allostérie (du grec ἄλλoς, allos : autre et στερεός, stereós : solide) est un mode de régulation de l'activité d'une protéine oligomérique par lequel la fixation d'une molécule effectrice en un site modifie les conditions de fixation d'une autre molécule, en un autre site distant de la même protéine.
Ce concept a été formalisé par Jacques Monod, Jean-Pierre Changeux et Jeffries Wyman (en) dans une série d'articles, dont le plus important a été publié en 1965 dans Journal of Molecular Biology[1].
Principe de l'allostérie
La fixation de la molécule effectrice induit un changement de conformation spatiale de la protéine enzymatique. Autrement dit, la disposition spatiale de ses atomes constitutifs est modifiée. Cela a pour conséquence de modifier le site de liaison et ses réactifs impliqués dans le processus de catalyse. Dans le modèle de Monod-Wyman-Changeux (MWC), les enzymes allostériques doivent présenter plusieurs propriétés :
- elles sont multimériques, chaque protomère (ou monomère) fixe une molécule de ligand ;
- elles possèdent au moins un axe de symétrie ;
- elles existent sous deux conformations différentes : l'une appelée T, pour tendue, désignant conventionnellement la forme de faible affinité pour le substrat, l'autre R, pour relaxée, de forte affinité pour le substrat ;
- au sein d'une protéine, les protomères adoptent tous la même configuration, R ou T (transition concertée). En d'autres termes, il n'existe pas d'hybrides R/T dans le modèle MWC[2].
Il existe une allostérie dite positive où la fixation d'un effecteur augmente l'affinité de liaison du ligand. On parle de fixation coopérative. La molécule effectrice peut être le ligand lui-même, qui modifie dans ce cas l'affinité des autres sites de fixation (effet homotrope, par opposition à l'effet hétérotrope qui concerne des molécules de nature différente). Ce système est le système K (K est le symbole de la constante d'affinité), dont la courbe de la vitesse de réaction en fonction de la concentration en substrat est de type sigmoïde. Le cas inverse existe aussi : dans une allostérie négative la fixation de l'effecteur diminue l'affinité du ligand. Ce système est le système V (pour vitesse), dont la courbe représentative est de type hyperbolique.
Exemple de l'hémoglobine
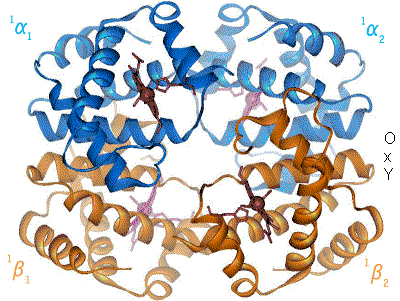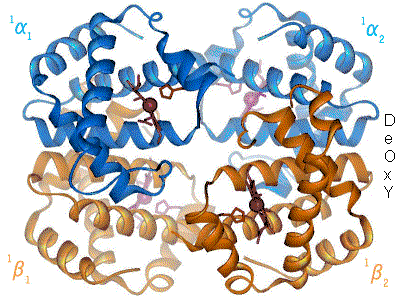
L'hémoglobine constitue un exemple important de protéine allostérique, bien qu'elle ne soit pas une enzyme stricto sensu mais plutôt une molécule de transport. Chaque monomère de l'hémoglobine, qui en comporte quatre, peut fixer une molécule de dioxygène. La fixation de la première molécule de dioxygène augmente l'affinité de liaison de la seconde, la fixation de la seconde augmente l'affinité pour la troisième et ainsi de suite (coopération positive par effet homotrope). Les effecteurs hétérotropes modifiant l'affinité de l'hémoglobine pour le dioxygène sont : le proton H+, la température, le dioxyde de carbone et le 2,3-diphosphoglycérate (un sous-produit de la glycolyse anaérobie ayant lieu dans les érythrocytes).
Importance physiologique de l'allostérie
L'allostérie est une forme de régulation de l'activité d'une enzyme en fonction des conditions extérieures changeantes. On peut distinguer les effets de senseurs et de rétrocontrôle négatif :
- lorsque la glycolyse est soutenue, le 2,3 biphosphoglycérate (2,3 BPG) s'accumule dans le globule rouge. Cette accumulation est un signal signifiant que l'utilisation d'énergie est en augmentation et que l'hémoglobine doit délivrer plus de dioxygène aux tissus. L'effecteur allostérique 2,3 biphosphoglycérate diminuant l'affinité du dioxygène pour l'hémoglobine, ce dernier se détache plus facilement et peut être délivré en plus grande quantité aux tissus. De la même manière, un pH acide, reflet d'un rejet accru de dioxyde de carbone, lui-même synonyme d'augmentation de l'activité physiologique, va provoquer une diminution de l'affinité de l'hémoglobine pour le dioxygène et donc un relargage au niveau des tissus, augmentant ainsi leur oxygénation. Cet effet (pH acide, par exemple) s'appelle l'effet Bohr, du nom de son découvreur ;
- l'effet de rétrocontrôle négatif est très souvent rencontré dans les voies métaboliques. Le produit final de la voie est souvent un inhibiteur allostérique d'un enzyme catalysant une étape initiale. Plus le produit final s'accumule, plus la réaction initiale est lente (par diminution de l'affinité à l'un des réactifs de la réaction) et donc moins de produit sera formé. Ce type de régulation évite d'accumuler le produit final, qui peut être toxique dans certaines circonstances et évite à l'organisme de produire en excès une molécule, ce qui est coûteux en énergie. L'effecteur allostérique participe à l'autorégulation de la voie métabolique :
- par exemple, dans la glycolyse, la phosphofructokinase-1 (PFK1) est une enzyme allostérique qui transforme le fructose 6-P en fructose 1,6-biP au cours de l'étape qui peut être considérée comme la première qui soit propre à cette voie. Cette enzyme réagit différemment en fonction de la quantité d'ATP, qui est le produit final « utile » de la glycolyse. À faible concentration, l'ATP se fixe sur le site « coenzyme » de l'enzyme, entraînant de fait toute l'enzyme sous sa forme relâchée. À forte concentration, l'ATP se fixe sur le site « inhibiteur » de l'enzyme, entraînant de fait toute l'enzyme sous sa forme tendue. Or l'ATP est le produit de la respiration cellulaire, phénomène qui suit la glycolyse et le cycle de Krebs. L'effet rétrocontrôle négatif de l'ATP est donc : inhibition de la voie lorsqu'il est en concentration suffisante, prévenant ainsi la dégradation de glucose non nécessaire. Un autre exemple intéressant et historique est celui de l'aspartate transcarbamylase, enzyme clé dans la voie de biosynthèse des pyrimidines, où l'ATP est un activateur et le CTP (cytosine triphosphate) et l'UTP (uracile triphosphate) sont inhibiteurs.
Extension de la notion aux canaux ioniques
Par extension, la notion d'allostérie s'applique au cas des canaux ioniques bien qu'il n'y ait pas de réaction chimique catalysée mais plutôt un transfert de matière (ion) à travers une membrane. Dans ce cas, la fixation d'une molécule effectrice positive favorisera l'ouverture du canal, quel que soit le mode d'ouverture du canal et, inversement, pour un effecteur négatif, le processus d'ouverture sera défavorisé. On parle d'agoniste et d'antagoniste des récepteurs allostérique respectivement.
Conséquence sur le repliement des protéines
Jusqu'à la découverte de l'allostérie, les mécanismes de repliement des protéines s'appuyaient sur un modèle uniquement thermodynamique. Le modèle de l'allostérie permet d'introduire une composante cinétique dans l'étude de ces repliements, qui sera démontrée en France par les travaux parallèles de Jeannine Yon et de Michel Goldberg[3].
Notes et références
- (en) Jacque Monod, Jeffries Wyman et Jean-Pierre Changeux, « On the nature of allosteric transitions: A plausible model », Journal of molecular biology, , Pages 88-118 (DOI https://doi.org/10.1016/S0022-2836(65)80285-6, lire en ligne)
- Cette limitation n'existe pas dans le modèle de Daniel Koshland, dit modèle séquentiel.
- Jérôme Segal, « Les premiers « replieurs » français : Michel Goldberg à l’Institut Pasteur et Jeannine Yon à Orsay », La revue pour l’histoire du CNRS, nos 7 /2002, (DOI https://doi.org/10.4000/histoire-cnrs.540, lire en ligne)
Voir aussi
 Portail de la biochimie
Portail de la biochimie