Bois (cervidé)
Les bois sont des organes osseux ramifiés de grande taille présents sur la tête d'animaux tels que le cerf et la plupart des représentants de sa famille, les cervidés.
Pour les articles homonymes, voir Bois (homonymie) et Panache.
Pour les articles homonymes, voir Bois-d’orignal.

Principales caractéristiques
Des 40 espèces existantes de cerfs, tous à l’exception du cerf d’eau de Chine, Hydropotes inermis, et les trois espèces de cerfs porte-musc, Moshus esp., développent des bois. Chez la plupart des cervidés, seul le mâle porte des bois. Chez le renne, le mâle et la femelle portent des bois ; toutefois, les bois du renne mâle sont beaucoup plus grands que ceux du renne femelle. Le cervidé ayant porté les bois de plus grande taille était le Megaloceros, maintenant éteint. L'espèce actuelle portant les bois les plus importants est l'Alces (élan ou orignal).

Les bois tombent et repoussent chaque année pour atteindre leur plein développement durant la période de rut. Ils jouent un rôle de caractère sexuel secondaire.
Des recherches conduites par George et Peter Bubenik tendent à prouver que les bois des orignaux ont une fonction d'amplification des sons. Dans un article publié en 2008 dans le European Journal of Wildlife [1], ces chercheurs ont montré que les orignaux portant des bois avaient une meilleure audition que ceux qui en étaient dépourvus. Les bois agiraient comme une sorte de réflecteur parabolique.

Du point de vue anatomique, les bois sont très différents des cornes : il s'agit d'organes osseux vascularisés et caducs, qui tombent et repoussent chaque année, et non de gaines cornées recouvrant une cheville osseuse. La croissance se déroule de façon continue sur un an. Au cours de leur croissance qui débute au printemps, les bois sont d'abord recouverts d'un tissu tégumentaire (le velours) qui assure la protection, la vascularisation et l'innervation de ces organes. Ce tissu se dessèche et tombe lorsque la croissance osseuse est achevée, vers la fin de l'été ; c'est la période de frayure, pendant laquelle l'animal se frotte à des éléments solides de son environnement naturel pour accélérer cette chute. Les bois resteront à nu pendant toute la période de rut et ne seront vascularisés qu'en interne[2], les velours étant tombés. Après la période de rut, à la fin de l'hiver, le bois se détache du crâne et son emplacement reste marqué par un pédicule jusqu'à la croissance des nouvelles pousses. Les nouveaux bois sont appelés des refaits[3]. Le produit de la mue (les bois morts délaissés) s'appelle la « mue »[4].
Terminologie
Bien que les bois repoussent chaque année, leur taille varie avec l’âge pour de nombreuses espèces, augmentant chaque année pendant plusieurs années avant d’atteindre leur taille maximale.
Situés sur la tête du cerf, sur la zone appelée pivot :
- les premiers bois sont des dagues, vers 1 an et demi ;
- à partir de 2 ans et demi, les bois seront des merrains, comportant des embranchements : les andouillers ou cors ;
- à partir de 3 ans et demi, les andouillers seront, dans l’ordre en partant du crâne, l’andouiller de massacre comme première ramification ; le surandouiller comme deuxième ramification ; la chevillure comme troisième ramification ; la trochure comme quatrième ramification ; et l’époi comme dernière ramification, qui fait partie de l’enfourchure si la perche se termine en forme de fourche à deux dents, ou bien de l’empaumure si la perche se termine en forme de main à plusieurs doigts.
En cynégétique, une paire de bois est appelée « trophée » (y compris sur l'animal vivant) ou « massacre » (uniquement tirée de l'animal mort, et généralement montée sur support).
Intérêt pour la science et la médecine
Les bois, trouvés seulement chez les cervidés, se composent d’os vivant et sont uniques parmi les appendices de mammifères dans leur capacité de se régénérer complètement et périodiquement après une abscission complète chez les adultes.
L’étude du développement des bois est utile à la compréhension des interactions peau-os lors de la cicatrisation, la régénération et le contrôle neuronal au développement des os. Cette compréhension peut être couplée à la médecine humaine avec une application à l’accélération de la régénération de la peau chez les grands brûlés et au retardement du développement de l’ostéoporose[5].
Classification
Structurellement, les bois diffèrent de tous les autres appendices crâniens chez les ruminants [6] et peuvent être considérés comme une automorphie de la famille des Cervidae.
La famille des cervidés est caractérisée par la séparation des espèces selon leurs appendices crâniens, les bois.
La présente taxonomie morphologique sépare la famille des Cervidae sur la base d’un seul critère : l’absence (Hydropotinae) ou la présence (Odocoileinae + Cervinae) des bois. Toutefois, la valeur taxonomique des bois a souvent été questionnée[7] et la réduction drastique des épois chez les genres sud-américains Pudu et Mazama suggère l’inversion de la tendance morphologique est possible en conséquence d’une sélection corrélée avec une petite taille d’un organisme[8].
La classification des Cervidae assume l’homoplasie de certains traits morphologiques et compte sur les bois comme une synapomorphie. Cependant, on a démontré que les appendices crâniens ont évolué indépendamment au moins trois fois chez les ruminants[6].
Grâce aux analyses cladistiques qui ont détecté plusieurs cas de parallélisme et de Convergence évolutive des traits morphologiques chez les Cervidae, la compréhension des patrons de développement des caractères nécessite un portrait phylogénétique indépendant de ces traits. L’approche moléculaire est essentielle pour résoudre la phylogénie incertaine des Cervidae. Les protéines codantes de l’ADN mitochondriale et, en particulier, le gène mitochondrial complet du cytochrome b a prouvé son utilité à résoudre la phylogénie de plusieurs artiodactyles avec un temps évolutionnaire plus bas que 20 millions d’années[9].
La comparaison des séquences de nucléotides des cytochrome b mitochondriale des 11 cervidés confirme la précédente observation et apporte une nouvelle information phylogénétique significative sur les relations des Cervidae.
L’association d'Hydropotinae avec Odocoileinae implique que les bois ont été perdus chez Hydropotinae et nous venons à la conclusion que les bois ont été probablement acquis une fois avant la radiation chez les Hydropotinae modernes[9].
Origine
Des six familles actuelles de ruminants, quatre familles possèdent des coiffes (cervidés, bovidés, girafés et les antilocapridés), chacun avec différent type (bois, cornes, et ossicônes).
Les trois formes de coiffes existantes ont un développement et un positionnement semblable ce qui inspire l’hypothèse de l’homologie. Ils sont en paires et habituellement symétriques, situés sur l’os frontal, formées à partir d’un cornillon couvert par du tissu et leur présence est étroitement relié au mâle.
En contraste, les fortes différences de leur développement et leur physiologie ont mené à l’hypothèse d’origines multiples.
L’origine des structures des bois de cervidés est encore inconnue.
Un précurseur probable des bois de cerf peut être les bosses frontales.

Des bosses frontales ont été découvertes sur des fossiles nord-américains tel que Aletomeryx[10].
Face à une collision imminente, l’animal aura avantage à réduire les dommages en présentant ses bosses frontales à la menace se dirigeant dans sa direction. Si les bosses sont plus longues que l’arme présentée, l’attaquant peut échouer d’atteindre des parties vulnérables de son opposant. Les bosses frontales ne pouvant devenir trop longues sans risques de fracturation ou d'entailler la peau qui recouvre les bosses, ces désavantages favorisent plutôt la croissance de prolongement osseux « mort » pour recouvrir les bosses frontales[pas clair][10].
Des chercheurs ont émis l’hypothèse que le pédicule des bois était homologue aux ossicônes ou aux cornes. Conséquemment, la portion d’os exposée des bois a été interprétée comme une nouveauté évolutionnaire, se développant au bout d’un ossicône[5].
Tendance évolutive
La grosseur et la complexité des bois (avec plus de branches) réduisent les chances de blessures à la tête. En démonstration, le mâle Axis axis a des bois plus longs mais plus simples avec seulement deux épois. Lors du combat, leurs bois entrent en contact étroits et sont très près des yeux de leur opposant ce qui peut occasionner des blessures importantes aux yeux et à la tête. En opposition, la plupart des cervidés (Odocoileus, Cervus, Dama, Alces, Rangifer) ont des bois plus complexes avec deux ou plusieurs épois de distances. Lorsque deux mâles entrechoquent leurs bois lors d’un combat, leurs têtes sont si éloignées, qu’aussi longtemps que le contact bois-à-bois est maintenu, aucune blessure ne peut être infligée[10].
.jpg.webp)
Il est plus avantageux d’avoir des plus grands bois qui sont plus efficaces comme bouclier contre les attaques en plus d’utiliser les bois de son adversaire comme levier pour le déstabiliser.
On peut imaginer que l’évolution des armes des cervidés mènera ultimement à des bois larges et palmés avec des épois courts et peu pointus. Ces bois « idéaux » n’infligeraient aucune blessure et instaureraient une hiérarchie par l’apparence plutôt qu’avec les combats, et il en résulterait des combats qui seraient gagnés par une démonstration de force et d’endurance à la place de provocation de blessures handicapantes ou fatales[10].
Fonction

Les appendices crâniens sont très communs, voyants et diversifiés dans le règne animal. Ils ont diverses fonctions souvent pour la sélection des partenaires sexuels, mais aussi pour la recherche de nourriture, les sens et même aider au déplacement[5].
Trois raisons principales pour expliquer l’origine des bois chez les mâles : défense contre les prédateurs, structure appréciée par les femelles et une arme qui peut servir entre les mâles combattants pour le territoire et/ou un accès aux femelles[11].
Des bois plus évidents favorise la valeur communicative de l’affichage de la dominance et donc réduit la fréquence des combats.La grandeur des bois est un bon indicateur de la force de l’animal[10].
La sélection sexuelle a joué un rôle dans l’évolution de la complexité des bois. Ce sont les plus grandes espèces de cerfs qui ont les bois les plus complexes mais ce sont aussi ceux qui ont un plus fort dimorphisme sexuel et une forte sélection sexuelle intraspécifique[12].
La comparaison entre les espèces révèle qu’il y a une relation allométrique positive entre la taille des bois et la taille de l’animal[8]. Les espèces de petites tailles ont des bois simples et petits, tandis que les plus grandes espèces ont proportionnellement des bois ramifiés plus gros.
Les espèces ayant un harem plus grand ont, relativement à leur taille, de plus grand bois comparé avec les espèces qui ont un petit groupe. Les espèces polygames ont augmenté la compétition entre mâles pour l’accès aux femelles durant le rut ce qui favorise l’évolution de plus grandes armes de combats[13].
Retirer la majeure partie des bois d’un animal vivant dans un groupe de mâle mène à une rapide régression de son rang hiérarchique dû à son incapacité de combattre. Le succès reproducteur durant le rut est également altéré, mais revient quand l’animal développe de nouveaux bois et reprend son rang social dominant l’année suivante[13].
Évolution et développement : l'asymétrie fluctuante des bois de Cervidés
Des études ont été menées sur des élans (Alces alces)[14] et rennes (Rangifer tarandus)[15] en Norvège afin de déterminer si les bois étaient de bons indicateurs de qualité individuelle. Les bois sont des traits dits ornementaux. Chez les mâles de ces espèces, comme pour tous les Cervidés, les ornements symétriques sont importants pour attirer l’attention des femelles. Les bois semblent être sujets à la sélection directionnelle du choix des femelles[15]. Si un mâle exprime des traits visuels asymétriques, qu’est-ce que cela révèle aux femelles sur sa qualité individuelle ? Seuls les traits ornementaux peuvent être sujets à l’asymétrie ? Souvent, les mâles plus âgés portent de plus gros ornements[14], et semblent être en meilleure santé[15]. Ces études tentent de prouver que les bois des rennes et orignaux sont de bons indicateurs de la qualité, de la condition et de l’âge d’un individu.

Photo de George Shiras III.
Les animaux sont programmés pour se développer de façon symétrique. Cependant, il est possible que la symétrie d’un trait dévie vers une asymétrie, et ce phénomène se nomme l’asymétrie fluctuante. L’asymétrie fluctuante est causée par un stress environnemental lors du développement de l’animal[15]. Il a été observé que les traits ornementaux plus imposants présentent moins d’asymétries[15]. Durant la période de rut des Cervidés, les bois sont pleinement développés[14]. La taille du mâle est souvent corrélée à la taille des bois. Sachant que les femelles préfèrent les mâles avec des bois plus gros, ces ornements sont soumis à une sélection directionnelle du choix des femelles. Les traits ornementaux sont plus sensibles au stress lors du développement que les traits non-ornementaux, ce pourquoi des bois symétriques et de bonne taille pourraient indiquer que l’individu a une meilleure résistance au stress environnemental[15]. Chez les rennes, les individus dont la charge parasitaire est élevée pourraient alors être démasqués par la présence d’asymétries sur les bois[15]. Les mâles orignaux plus âgés ont été remarqués pour avoir des bois plus importants[14]. Ces mâles pourraient avoir une meilleure capacité à résister aux stress environnementaux, puisqu’ils ont acquis beaucoup d’expérience. Il est alors important pour le mâle d’indiquer son âge aux femelles, ainsi qu’aux autres mâles pour les combats[14].
L’étude menée sur les rennes en Norvège a été effectuée sur 59 mâles semi-domestiques[15]. Tous ces mâles étaient entrés en période de rut, et leurs bois étaient dépourvus de velours. Des données morphométriques ont été récoltées sur ces animaux, incluant le poids, la taille du renne et de ses bois, ainsi que la longueur des mâchoires (trait non-ornemental). De plus, le nombre de pointes présentes sur les bois a été compté, pour ensuite calculer l’asymétrie fluctuante.Pour évaluer la charge parasitaire, les nématodes retrouvés dans l’estomac des rennes ont été recensés[15]. L’étude sur les orignaux a été effectuée sur 2395 mâles sur une période de 25 ans[14]. Les animaux ont été abattus lors de périodes de chasse. Tous les orignaux mâles étudiés portaient des bois dont la croissance était terminée. Comme pour les rennes, les pointes sur les bois ont été dénombrées et l’asymétrie fluctuante a été calculée. L’âge de ces mâles a été déterminé par l’analyse au laboratoire des dents[14].
Chez le renne, les résultats démontrent que la charge parasitaire n’a pas de lien avec le poids de l’animal, mais qu’elle est fortement liée à l’asymétrie fluctuante des bois[15]. Il est intéressant de noter que lorsque la longueur des bois augmente, l’asymétrie fluctuante diminue. De plus, la taille du mâle était positivement corrélée à la taille des bois. L’asymétrie fluctuante n’est pas liée à la longueur de la mâchoire, qui est un trait non-ornemental[15]. Chez l’orignal, le nombre de pointes sur les bois augmente en fonction de l’âge du mâle[14]. Contrairement au renne, l’asymétrie fluctuante augmente avec la taille des bois. Cependant, plus le mâle est âgé, plus la corrélation entre la taille des bois et l’asymétrie fluctuante s’estompe. Pour une taille de bois donnée, les mâles plus âgés présentent moins d’asymétrie fluctuante vis-à-vis leurs ornements[14].
La relation négative entre l’asymétrie fluctuante observée sur les bois de rennes et leur taille montre que les bois sont bel et bien des ornements dépendants des conditions extérieures, contrairement aux traits non-ornementaux comme les mâchoires[15]. L’effet de la sélection directionnelle du choix des femelles maximise l’élaboration des bois et en diminue par le fait même l’homéostasie, ce qui rend les ornements plus vulnérables aux stress environnementaux, et donc à l’asymétrie fluctuante. Les mâles de plus grande taille ont des ornements plus imposants, et présentent alors moins d’asymétries[15]. Les rennes ont besoin de minéraux pour développer leurs bois. Les individus ayant des ornements plus gros et symétriques indiquent leur excellente capacité à acquérir les ressources alimentaires nécessaires. De plus, cette étude a prouvé que la charge parasitaire d’un individu peut être détectée par l’asymétrie qu’affiche cet individu[15]. À première vue, la relation positive entre la taille des bois des orignaux et l’asymétrie fluctuante contredit les suppositions émises antérieurement. Cependant, il a tout de même été prouvé que cette relation diminuait plus l’âge de l’individu augmentait[14]. Les orignaux mâles plus âgés indiquent une meilleure qualité génétique, car ils ont réussi à survivre malgré tous les stress environnementaux. Il est donc possible d’affirmer que la combinaison de la taille des bois et de l’asymétrie signale l’âge de l’individu, en plus de sa qualité individuelle[14]. En bref, les traits ornementaux des Cervidés, tels les bois, sont de bons indicateurs visuels de leur qualité individuelle. Ils sont utilisés comme signaux intrasexuels et intersexuels.
Histogenèse
Durant la première année d’élongation, la croissance passe par trois étapes d’ossification : endoconjonctive, transitionnelle et enchondrale[16].
Durant l’étape d’ossification endoconjonctive, la crête latérale de l’os frontal se forme par croissance périostique de l’os spongieux. Le périoste à cet endroit est inusité car il contient des cellules simili-embryonnaire et la transplantation expérimentale a démontré l’habilité de ce périoste de former des bois ectopiques. De plus, cette capacité intrinsèque est fournie par induction expérimentale qui montre que le signalement entre la peau et les os n’est pas nécessaire pour former la crête latérale. La déposition d’os spongieux en continu sur le dessus de la crête produit une bosse palpable appelée pédicule[16].
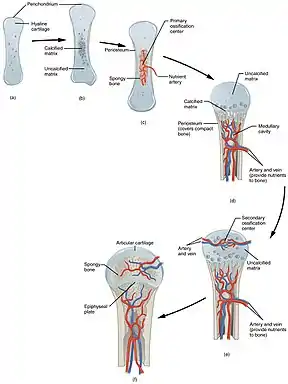
Durant l’étape de l’ossification transitionnel, l’élongation s’épaissit et produit un court mais visible pédicule. Le périoste apical subit une transformation partielle en périchondre, résultant en une déposition apicale d’os et de tissus cartilagineux hautement vascularisés. Bien que l’intensification de la croissance du pédicule coïncide typiquement avec la puberté du mâle et l’augmentation de testostérone, d’autre molécule de signalement entre périoste et le tégument sont nécessaires pour initier la formation du pédicule. Les facteurs de croissance jouent un rôle majeur dans le développement des bois. Le facteur de croissance 1 ressemblant à l’insuline (IGF1) est un facteur de croissance crucial pour la stimulation de la croissance des bois. Il existe une forte corrélation positive entre le taux de croissance des bois et la concentration circulatoire de IGF1. La localisation de IGF1 dans les chondrocytes et ostéoblastes suggère que IGF1 peut jouer un rôle important dans la formation du cartilage et des os[16].
La mélatonine (MLT), une hormone multifonction, joue un rôle important dans la régulation de l’équilibre d’oxydoréduction, le cycle circadien, l’activité métabolique et l’immunité. La voie de signalisation du récepteur à la mélatonine joue également un rôle important dans la prolifération et la différenciation cellulaire. Il a été démontré que la mélatonine favorise la croissance des bois. L’administration orale de mélatonine raccourcit le cycle des bois du Cerf de Virginie, le nombre de cycles par année augmente d’un à deux[16]. Une étude rapporte que le niveau d’IGF1 durant le cycle de croissance des bois est affecté par le traitement à la mélatonine[17].
Durant la régénération des bois, les cellules souches du périoste se différencient constamment pour former les cellules mésenchymateuses, puis se transforme en cellules cartilagineuses, ce qui favorise la croissance des bois. Aussi, la concentration de mélatonine sanguin modifie la régularité, ce qui indique que la mélatonine agit dans ce processus.
Il a été démontré que la voie de signalisation IGF1 fonctionne en aval de la voie de signalement de la mélatonine pour réguler la prolifération des cellules mésenchymateuses des bois[16].
La croissance du pédicule est complétée et la genèse des bois débute durant l’étape d’ossification enchondrale. Pendant la croissance des bois, le périoste se transforme en tissu mésenchyme et les cellules mésenchymateuses se développent en cellules cartilagineuses. C’est la prolifération du mésenchyme qui détermine la vitesse de croissance des bois. Le périoste apical est complètement transformé en périchondre et commence à déposer du tissu cartilagineux hautement vascularisé, qui s’ossifie par la voie endochondrique, comme les longs os. Le tégument des bois se transforme en une forme glandulaire appelée velours. La terminaison de la croissance des bois coïncide avec un fort relâchement d’androgène dans la circulation sanguine et le début du rut. C’est seulement le tégument qui recouvre les bois qui mue, le pédicule reste recouvert, laissant l’os nu. Le tissu osseux en périphérie meurt, cependant, le tissu osseux plus profond reste vivant et continue de former l’os compact. Les bois tombe quand le niveau d’androgène diminue et de nouveaux bois croient de nouveau par voie endochondrale sur le pédicule quand le niveau d’androgène est à son plus bas[16].
Exception

Des habitats fermés ont été colonisés par Capreolini, Muntiacini, Mazama americana, Mazama gouazoubira et Pudu et ces cervidés ont développé une morphologie adaptative semblable, incluant une réduction et une simplification des bois chez les mâles et l’acquisition d’une petite taille accompagnée par un monomorphisme sexuel. Ces réductions ont été positivement sélectionnées car les grands mâles avec des bois longs et ramifiés se déplacent plus lentement dans des habitats fermés et que la fonction d’exposition des bois est moins efficace lorsqu’il est situé dans un environnement où la visibilité est réduite[11].
Parallèlement à ces réductions morphologiques, les mâles Hydropotes et Muntiacini ont acquis des canines qui leur servent de défense durant le rut qui remplacent les bois dans les comportements sexuels de ces espèces[11].
Ceci supporte l’hypothèse que le rôle primaire des bois est utilisé dans la compétition sexuelle durant la saison de reproduction. Cette importante fonction peut expliquer la raison pour laquelle les bois sont maintenus chez tous les cervidés à l’exception des Hydropotes[11].
Le succès reproductif des mâles est corrélé avec la force et la taille des bois, c’est pourquoi les grands mâles avec un important armement sont positivement sélectionnés durant l’évolution des Cervidae. Ceci explique pourquoi les bois sont particulièrement développés et ramifiés chez les mâles des espèces vivant dans des habitats ouverts[11].
La seule espèce dans laquelle les femelles ont des bois est Rangifer tarandus. Comme les bois peuvent servir d’arme et parce qu'un haut pourcentage de mortalité chez les caribous est dû aux loups et au ours, la prédation peut avoir constitué une force de sélection suffisante pour l’apparition des bois chez les femelles. Cependant, cette pression de prédation est également appliquée pour Alces et existe également pour Odocoileus, mais ces femelles ne possèdent pas de bois[13].

L’hypothèse alternative suggère que des femelles avec des bois ont plutôt été sélectionnées positivement par la grégarisme. En fait, la compétition intraspécifique pour la nourriture est importante chez les hardes de sexe confondu, spécifiquement durant l’hiver lorsque la quantité de neige est élevée sur de longue période[13].
Les bois chez les femelles caribous peuvent avoir évolué comme une adaptation découlant de la compétition intraspécifique pour la nourriture durant l’hiver plutôt que la défense contre les prédateurs.
Chez les caribous la croissance des bois chez les jeunes débute bien avant la puberté. Ceci suggère que l’initiation de la croissance des bois n’est pas contrôlée par la sécrétion des hormones sexuelles. Il a été démontré qu’en retirant des gonades des faons de caribous d’une à deux semaines suivant la naissance on n’empêche pas la croissance des bois chez les deux sexes[18]. De plus, le renouvellement annuel des bois est maintenu même suite la castration chez un caribou mâle adulte, ce qui n’est pas le cas chez les autres espèces de cerfs[13].
Utilisation
Le bois de cerf a été un matériau important dès le Paléolithique supérieur. Du fait de sa dureté, il a d'abord servi à la fabrication d'outils tranchants et perforants (pointe de sagaies, harpons, aiguilles à chas…). Au Néolithique, on s'en servait pour fabriquer des pics, des marteaux, des manches pour les lames en silex, des éléments intermédiaires pour emmancher les haches polies, etc. On continua de l'utiliser jusqu'au Moyen Âge[19], notamment pour la fabrication de peignes.
- Objets en bois de cervidés
 Peigne.
Peigne. Gobelets.
Gobelets. Décapsuleur.
Décapsuleur. Lustre.
Lustre. Porte-manteau.
Porte-manteau.
Représentations
_CoA.png.webp)
Les bois sont un meuble en héraldique.
Cernunnos est une divinité gauloise dont la tête est ornée de bois de cerf.
Références
- (en) Moose's sharp hearing is attributed to antlers
- Crigel Marie-Hélène, « Les bois de cerf : revue de littérature scientifique », Article de synthèse, , p. 29 (lire en ligne)
- Encyclopédie pratique de la chasse, Georges Cortay, Pascal Durantel, De Borée Éditions, 2006
- Photos de mues de chevreuil sur Le Solitaire Ardennais, forum de la chasse en Wallonie.
- (en) Edward Byrd Davis, Katherine A. Brakora et Andrew H. Lee, « Evolution of ruminant headgear: a review », Proceedings of the Royal Society of London B: Biological Sciences, vol. 278, no 1720, , p. 2857–2865 (ISSN 0962-8452 et 1471-2954, PMID 21733893, DOI 10.1098/rspb.2011.0938, lire en ligne, consulté le )
- (en-US) Christine M. Janis et Kathleen M. Scott, « The interrelationships of higher ruminant families : with special emphasis on the members of the Cervoidea. American Museum novitates ; no. 2893 », smithsonian institution, (lire en ligne, consulté le )
- (en) Scott, K. M. & Janis, C. M., « Relationships of the Ruminantia (Artiodactyla) and an analysis of the characters used in ruminant taxonomy. In Mammal phylogeny: placentals », spinger,
- (en) Gould, S.J., « The origin and function of `bizarre' structures: antler size and skull size in the `irish elk', Megaloceros giganteus. », Evolution, , p. 191-220
- (en) Ettore Randi, Nadia Mucci, Massimo Pierpaoli et Emmanuel Douzery, « New phylogenetic perspectives on the Cervidae (Artiodactyla) are provided by the mitochondrial cytochrome b gene », Proceedings of the Royal Society of London B: Biological Sciences, vol. 265, no 1398, , p. 793–801 (ISSN 0962-8452 et 1471-2954, PMID 9628037, DOI 10.1098/rspb.1998.0362, lire en ligne, consulté le )
- Cyrille Barrette, « Fighting Behavior of Muntjac and the Evolution of Antlers », Evolution, vol. 31, no 1, , p. 169–176 (DOI 10.2307/2407555, lire en ligne, consulté le )
- Clément Gilbert, Anne Ropiquet et Alexandre Hassanin, « Mitochondrial and nuclear phylogenies of Cervidae (Mammalia, Ruminantia): Systematics, morphology, and biogeography », Molecular Phylogenetics and Evolution, vol. 40, no 1, , p. 101–117 (DOI 10.1016/j.ympev.2006.02.017, lire en ligne, consulté le )
- (en) « Ornament Complexity Is Correlated with Sexual Selection: (A Comment on Raia et al., “Cope’s Rule and the Universal Scaling Law of Ornament Complexity”) », sur ResearchGate (consulté le )
- (en) G. A. Lincoln, « Biology of antlers », Journal of Zoology, vol. 226, no 3, , p. 517–528 (ISSN 1469-7998, DOI 10.1111/j.1469-7998.1992.tb07495.x, lire en ligne, consulté le )
- Solberg, E. J., & Saether, B. E. (1993). Fluctuating asymmetry in the antlers of moose (Alces alces): does it signal male quality?. Proceedings of the Royal Society of London. Series B: Biological Sciences, 254(1341), 251-255
- Markusson, E., & Folstad, I. (1997). Reindeer antlers: visual indicators of individual quality?. Oecologia, 110(4), 501-507.
- (en) Feifei Yang, Changjiu He, Xuyang Sun et Jing Wang, « The Regulatory Mechanism of MLT/MT1 Signaling on the Growth of Antler Mesenchymal Cells », Molecules, vol. 22, no 10, , p. 1793 (DOI 10.3390/molecules22101793, lire en ligne, consulté le )
- (en) JAMES M. SUTTIE, PETER D. GLUCKMAN, JOHN H. BUTLER et PETER F. FENNESSY, « INSULIN-LIKE GROWTH FACTOR 1 (IGF-1) ANTLER-STIMULATING HORMONE? », Endocrinology, vol. 116, no 2, , p. 846–848 (ISSN 0013-7227, DOI 10.1210/endo-116-2-846, lire en ligne, consulté le )
- (en) Gerald A. Lincoln et Nicholas J. C. Tyler, The Biology of Deer, Springer, New York, NY, (ISBN 9781461276678 et 9781461227823, DOI 10.1007/978-1-4612-2782-3_118, lire en ligne), p. 493–498
- Henrique Sarmento-Pedro, « Le bois de cervidé, une matière première de l’artisanat du Moyen-Âge », Microscoop, CNRS - Délégation Centre Limousin Poitou-Charentes, vol. 83, , p. 16-17 (ISSN 1291-8083, lire en ligne). Numéro complet.
Voir aussi
Articles connexes
Liens externes
 Portail de la zoologie
Portail de la zoologie  Portail de l’anatomie
Portail de l’anatomie