Embryophyta
Les Embryophytes, rassemblent ce qu'on appelle les plantes terrestres. Elles constituent la super-division des Embryophyta, aussi dénommées Cormophytes (« plantes à tige », du latin cormus, tige ; du grec kormos, tronc) ou Archégoniates (végétaux dont le gamétange femelle est un archégone). Elles forment un groupe monophylétique, qui s'est développé parmi les chlorobiontes durant l'ère du Paléozoïque et s'est adapté avec succès à la vie sur la terre ferme, qu'elles ont progressivement colonisée, la sortie des eaux étant d'abord le fait des bryophytes avec leur gamétophyte aérien et durable [réf. souhaitée]. Ce terme de plantes terrestres n'interdit pas que certaines d'entre elles se soient par la suite adaptées aux milieux aquatiques (à la manière des cétacés chez les animaux) ou que leur fécondation reste en milieu aquatique[2].
Cet article concerne l'ensemble des plantes terrestres correspondant à la classe Equisetopsida dans la classification phylogénétique APG III (2009)[1]. Pour la classe Equisetopsida de la nomenclature botanique correspondant aux prêles, voir Equisetidae.

| Règne | Plantae |
|---|---|
| Sous-règne | Viridiplantae |
| Infra-règne | Streptophyta |
Avec les algues vertes, ou chlorophytes lato sensu (groupe paraphylétique), elles constituent le règne des plantes vertes et le clade des chlorobiontes caractérisé par l'association des chlorophylles a et b et l'amidon stocké dans les plastes[3].
Les embryophytes sont définies par la présence d'un embryon végétal, structure qui se développe après la fécondation à partir du zygote et qui contient un pied ou suçoir, un suspenseur et une tête. Ce zygote ou œuf dont le développement est polarisé, au moins des premiers stades, dépend des réserves d'origine maternelle. De même, l'embryon dépend au moins pour les premiers stades de développement de réserves d'origine maternelle. C'est la tête de l'embryon qui se développe en sporophyte avec racine, tige et feuille : le port plus ou moins érigé dans l'air doit en effet vaincre le poids et la verticalité sépare les territoires éclairés des zones d'ancrage et d'absorption. Ainsi, la conquête des terres implique la différenciation en organes et tissus spécialisés, parfois renforcés par de la lignine et recouverts de cuticule, revêtement hydrophobe[4].
Étymologie
Le terme Embryophytes vient du grec ancien έμβρυόν (embrión, « embryon »), et de φυτόν (phytón, « végétal »). Faisant référence à l'embryon diploïde (2n) pluricellulaire que les Embryophytes portent à un moment de leur cycle de vie.
Synapomorphies

Les principales synapomorphies (caractères dérivés propres) à ce groupe sont :
- la présence de stomates qui sont des orifices présents sur les feuilles et permettant de faciliter les échanges gazeux[5] (sauf chez les Marchantiophytes qui les auraient perdus secondairement au profit de pores aériens) ;
- la présence d'une cuticule recouvrant l'épiderme[5] ;
- le cycle digénétique (alternance de générations) qui « devient de plus en plus hétéromorphe avec une réduction de plus en plus importante du gamétophyte (aussi bien en taille qu'en durée de vie)[6] ». L'émergence de la phase diploïde du cycle (sporophyte) comme phase dominante pourrait être que la diploïdie autorise le masquage de l'expression de mutations délétères par complémentation génétique[7],[8] ;
- les gamètes sont formées dans des gamétanges mâles (anthéridies) et femelles (archégones où se produit la fécondation interne (en)). La réduction progressive du gamétophyte s'accompagne ainsi d'une protection de plus en plus grande de la phase haploïde[5].
Cycle de reproduction des Bryophytes
Marchantia polymorpha
Exemple du cycle de vie de Marchantia polymorpha, appartenant au taxon des marchantiophytes.
Phase gamétophytique[9]
Les spores sont tous identiques (homosporie). Ils sont donc, par définition, indifférenciés. Grâce à la germination, un gamétophyte se met en place. Cet individu est haploïde (à n chromosome). Ce dernier, met en place des gamétanges (structures à l’origine des gamètes) qui, par la suite, vont se différencier. À l'issue de la différenciation, il y a un thalle mâle et un thalle femelle.
Le thalle mâle met en place des anthéridiophores sur lequel on retrouve les anthéridies (gamétanges mâles) regroupés sous forme de corbeilles. Chaque anthéridie possède une assise cellulaire et à l’intérieur, se situe le tissu spermatogène. Parmi ces cellules, certaines sont nourricières et d’autres se différencient en spermatozoïdes (gamètes mâles).
Quant au thalle femelle, les archégones (gamétanges femelles) sont regroupés dans des structures appelées : archégoniophores. Chaque archégone est composé d’un oosphère, d’une paroi (assise cellulaire) et des cellules du col. Ces dernières se dégradent et permettent ainsi aux spermatozoïdes d’aller féconder l’oosphère (le gamète femelle). Il s’agit d’une fécondation par zoïdogamie (c’est-à-dire nécessitant la présence d’eau).
Phase sporophytique[10]
À l’issue de la fécondation, il y a un zygote (à 2n chromosomes) présent à l’intérieur de l’archégone du cycle précédent. Il s’ensuit plusieurs divisions cellulaires aboutissant sur le sporophyte. C’est un individu parasite très petit avec un suçoir. En effet, il est non chlorophyllien et puise ses ressources dans l’archégone du cycle précédent. Il dispose d’un tissu sporogène. Différentes cellules de ce tissu vont être à l’origine des élatères (elles restent diploïdes) et ont le rôle d’aider à l’expulsion des spores. D’autres sont des cellules “mères” qui vont subir la méiose et être à l’origine d’une tétrade de spores (cellules haploïdes). Les spores germent afin de donner le gamétophyte. Le cycle est ainsi terminé et une nouvelle génération débute.
Caractéristiques
Les caractéristiques de ce cycle sont donc :
- Phase gamétophytique est dominante
- Fécondation par zoïdogamie
- Dissémination par les spores
Évolution et développement

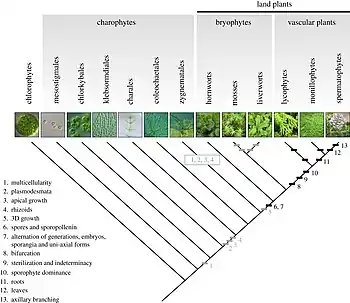
Dans la classification phénétique classique, les botanistes classent les Embryophytes en trois groupes :
- les Bryophytes (lato sensu : mousses, hépatiques, anthocérotes) ;
- les Ptéridophytes (fougères) ;
- les Spermatophytes (plantes à graines).
Cependant cette classification, toujours valide pour les évolutionnistes[réf. souhaitée], n'est pas acceptée par les cladistes puisque mousses et fougères forment des ensembles paraphylétiques[14].
La phylogénétique moléculaire suggère que les plantes terrestres forment un groupe monophylétique qui a divergé des algues vertes il y a près de 700 millions d'années[15]. Ces premières plantes sont caractérisées par un gamétophyte dominant colonisateur et un sporophyte dressé pour la dispersion. À cette période, la couche d’ozone est bien installée. La filtration des UV est suffisante et elle permet la vie hors de l’eau. Il y a donc une corrélation entre la composition de la couche d’ozone et la colonisation des plantes vertes sur Terre (Embryophytes). L’ancêtre commun des Embryophytes est donc apparu il y a 480 millions d’années (il y a ensuite une recolonisation des eaux douces par certains Embryophytes au fil du temps).
Pour pouvoir survivre en milieu terrestre, de nombreuses adaptations des plantes ont été réalisées au niveau de l'appareil végétatif et reproducteur, visant essentiellement à faire face à l'absence d'eau et à croître dans le milieu aérien en luttant contre l'attraction terrestre. Chez les plantes terrestres, on remarque que le sporophyte pousse verticalement pour maximiser la dispersion en milieu aérien (plus haut plus loin) : le thalle s'est progressivement transformé en cormus possédant des tissus spécialisés et bien différenciés. La forme cylindrique de ces axes dressés permet de diminuer la surface du végétal en contact avec l'atmosphère terrestre desséchante. Afin de permettre la croissance de végétaux de plus en plus hauts la mise en place d'un système racinaire efficace et d'un système de double circulation (xylème (sève brute) et phloème (sève élaborée)) permettant d'irriguer toutes les parties de la plante est nécessaire pour assurer des apports d'eau suffisants. Les axes se sont ensuite différenciés pour former différents organes végétatifs : tiges, feuilles, racines mais également reproducteurs à enveloppe pluricellulaire. On trouve ainsi les sporanges et gamétanges qui protègent les cellules reproductrices (les spores et gamètes). Il existe également plusieurs autres structures de protection afin de lutter contre les conditions hostiles du milieu terrestre : les spores disséminées par le vent sont protégées de la dessiccation par une épaisse paroi imprégnée de sporopollénine, le zygote, issu de la fécondation reste enfermé et protégé au début de son développement dans le gamétange femelle (ou archégone) où se développera le jeune embryon[16].
Aux alentours de 420-430 jusqu’à 400 millions d’années, on observe l’apparition d’organismes avec polysporangie. Les polysporangiés produisent plusieurs sporanges par sporophyte. Le sporophyte devient donc génération dominante avec la présence d’axes rampants et d’axes dressés : c’est l’apparition des Ptéridophytes au Silurien. Les Ptéridophytes sont des plantes vasculaires (Trachéophytes). Cependant, les premiers Ptéridophytes n’étaient pas vascularisés. Aglaophyton est un Ptéridophyte avec des hyphes de champignons, ce sont les premières plantes mycorhizées qui permettent une meilleure colonisation terrestre.
Les Trachéophytes ont des tissus conducteurs. Les plus vieux sont les Lycopodes, apparus à la fin du Silurien, avec des formes herbacées (axes rampants et axes dressés) et des formes arborescentes au carbonifère. Les plantes vasculaires à spores libres se diversifient au Dévonien. Au Dévonien, on trouve l’apparition des plantes à ovules aux alentours de 360 millions d’années. Ces Angiospermes sont issues des Progymnospermes qui se reproduisaient par des spores libres et avaient un tronc avec du bois. Le tronc est apparu avant l’ovule. Archaeopteris forme les premières grandes forêts de la fin du Dévonien. Les plantes à ovules apparaissent dans les sous bois des forêts.
Les plantes vascularisées produisent des spores qui germent pour donner un gamétophyte bisexué qui après fécondation conduira à la formation d’un embryon. Les Progymnospermes hétérosporés et hétérosporangie donnent les spermatozoïdes. Le microsporange, c’est-à-dire le sac pollinique, produit le grain de pollen et le mégasporange produit 4 spores. Dans le sporange, il y a un mégaprothalle (nucelle) dont la périphérie constitue le tégument.
Les Euphyllophytes possèdent de vraies feuilles, que l’on appelle des mégaphylles (= rameaux latéraux transformés avec du limbe). On y trouve les fougères, les prêles et les plantes à ovules. Les fougères et les plantes à ovules sont apparues à peu près au même moment mais de manière séparée. Les plantes à ovules viennent des Progymnospermes. Les Angiospermes arrivent au crétacé. Les Gymnospermes sont paraphylétiques[17].
Les Trachéophytes sont tous polysporangiophytes. Quant aux Mousses et Hépatiques, ce sont des unisporangiés. Les Euphyllophytes, les fougères et les prèles, sont des monilophytes. Les Gymnospermes sont paraphylétiques et les Angiospermes sont monophylétiques. Les Cycas sont les Gymnospermes les plus vieux (Carbonifères), puis sont apparus les Ginkgos (Permien secondaire), les Géophytes et les Angiospermes[18] (Mésozoïque).
Phylogénie
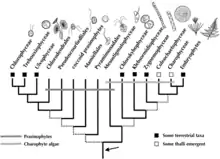
Jusque vers 2010, les données phylogénétiques semblaient indiquer que le groupe le plus proche des embryophytes était un groupe d'algues vertes, les charophytes, et plus particulièrement les coleochaetales[19]. Comme ces algues ne présentent aucun signe d'adaptation à la vie hors des océans, on pensait alors que la conquête du milieu terrestre s'était réalisée au sein du groupe des embryophytes[20].
Entre 2012 et 2018, ce sont les zygnématophycées, un groupe d'algues relativement peu étudié, qui se sont révélées les plus proches des embryophytes[20]. Fin 2019, le séquençage des génomes de différentes espèces des deux groupes (ainsi que d'autres végétaux) a montré que les embryophytes et les zygnématophycées ont notamment en commun deux gènes qui interviennent dans la résistance à la dessication, dénommés PYL et GRAS. On en déduit que le dernier ancêtre commun des deux groupes était déjà adapté à un mode de vie terrestre. Comme PYL et GRAS sont très fréquents dans les bactéries du sol, il est probable que cet ancêtre commun (ou l'un de ses ascendants) les a acquis par transfert horizontal depuis certaines de ces bactéries[20],[21].
Systématique
Classification
Liste des divisions selon ITIS (25 juin 2018)[22] :
Liste des sous-classes selon la classification phylogénétique APG III (2009)[1] :
- classe Equisetopsida C.Agardh, 1825 :
- Anthocerotidae Engl., 1893
- Bryidae Engl., 1892
- Marchantiidae Engl., 1893
- Lycopodiidae Beketov, 1863
- (Monilophytes) :
- Equisetidae Warm., 1883
- Marattiidae Klinge, 1882
- Ophioglossidae Klinge, 1882
- Polypodiidae Cronquist, 1966
- Psilotidae Reveal, 1996
- (Gymnospermes) :
- Ginkgooidae Engl., 1897
- Cycadidae Pax, 1894
- Pinidae Cronquist, Takht. & Zimmerm., 1966
- Gnetidae Pax, 1894
- (Angiospermes) :
- Magnoliidae Novák ex Takht., 1967
Selon Paleobiology Database (25 juin 2018)[23] :
- Angiospermae
- Antherocerotophyta
- Bryophyta
- Coniferophyta
- Cycadophyta
- Gigantopteridales
- Ginkgophyta
- Hermanophytales
- Iraniales
- Lycophyta
- Progymnospermophyta
- Pteridophyta
- Pteridospermophyta
- Sphenophyta
- Zosterophyllophyta
Notes et références
- (en) Angiosperm Phylogeny Group, « An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III », Botanical Journal of the Linnean Society, Wiley-Blackwell, Linnean Society of London et OUP, vol. 161, no 2, , p. 105–121 (ISSN 0024-4074 et 1095-8339, DOI 10.1111/J.1095-8339.2009.00996.X)
- Michel Godron, Écologie et évolution du monde vivant. Volume 1, L'Harmattan, , p. 170
- (en) Michael G. Simpson, Plant Systematics, Academic Press, , p. 5
- Jean-Claude Roland, Hayat El Maarouf Bouteau, François Bouteau, Atlas de biologie végétale, Dunod, , p. 122.
- Guillaume Lecointre et Hervé Le Guyader, Classification phylogénétique du vivant, t. 1, 4e éd., Paris, Belin, , 583 p. (ISBN 978-2-7011-8294-0)
- Yves Tourte, Michel Bordonneau, Max Henry et Catherine Tourte, Le monde des végétaux. Organisation, physiologie et génomique, Dunod, (lire en ligne), p. 48
- (en) H. Bernstein, G.S. Byers et R.E. Michod, « Evolution of sexual reproduction: Importance of DNA repair, complementation, and variation », The American Naturalist, vol. 117, no 4, , p. 537–549 (DOI 10.1086/283734).
- (en) R.E. Michod et T.W. Gayley, « Masking of mutations and the evolution of sex », The American Naturalist, vol. 139, no 4, , p. 706–734 (DOI 10.1086/285354).
- Phase gamétophytique
- Phase sporophytique
- Le milieu terrestre étant beaucoup moins tamponné thermiquement que la mer (écarts de température rapides et de grande amplitude), il impose des contraintes hydriques et thermiques supplémentaires.
- Les Trachéophytes sont des plantes homéohydres grâce à la présence de racines et de vaisseaux conducteurs. Les Bryophytes ne possèdent pas ces caractères et adoptent une autre stratégie de résistance au stress hydrique, la poïkilohydrie : comme les lichens, leur teneur en eau varie suivant les variations d'hygrométrie de l'environnement extérieur, de 10 à 90 % de leur poids frais, car ces végétaux sont incapables de contrôler la perte en eau de leurs cellules et se déshydratent facilement par toute leur surface. Elles restent pour beaucoup inféodées aux milieux humides, mais de nombreuses espèces sont aussi pionnières des milieux secs grâce à une stratégie d'évitement, l'anhydrobiose. Robert Gorenflot, Biologie végétale, Masson, , p. 213
- (en) Zoe Popper et al., « Evolution and diversity of plant cell walls: from algae to flowering plants », Annual Review of Plant Biology, vol. 62, no 1, , p. 567-590 (DOI 10.1146/annurev-arplant-042110-103809)
- Futura, « Embryophyte », sur Futura (consulté le )
- (en) YL Qiu, JD Palmer, « Phylogeny of early land plants: insights from genes and genomes », Trends Plant Sci., vol. 4, no 1, , p. 26-30 (DOI 10.1016/s1360-1385(98)01361-2.).
- « Évolution et caractéristiques-Origine et adaptation au milieu terrestre », sur uel.unisciel.fr (consulté le )
- Gerard Cusset, Botanique : Les Embryophytes, Elsevier Masson, , 512 p.
- « Classification des végétaux — Site des ressources d'ACCES pour enseigner la Science de la Vie et de la Terre », sur acces.ens-lyon.fr (consulté le )
- (en) Douglas E. Soltis, Pamela Soltis, J.J. Doyle, Molecular Systematics of Plants II : DNA Sequencing, Springer, , p. 530
- Coraline Madec, « Sortie des eaux des végétaux : un scénario revisité », Pour la science, no 507, , p. 8.
- (en) Shifeng Cheng, Wenfei Xian, Yuan Fu, Birger Marin, Jean Keller et al., « Genomes of Subaerial Zygnematophyceae Provide Insights into Land Plant Evolution », Cell, vol. 179, no 5, , p. 1057-1067, article no e14 (DOI 10.1016/j.cell.2019.10.019).
- Integrated Taxonomic Information System (ITIS), www.itis.gov, CC0 https://doi.org/10.5066/F7KH0KBK, consulté le 25 juin 2018
- Fossilworks Paleobiology Database, consulté le 25 juin 2018
Voir aussi
Références taxinomiques
- (en) Référence BioLib : Embryophyta (consulté le )
- (fr+en) Référence ITIS : Embryophyta (consulté le )
- (en) Référence NCBI : Embryophyta (taxons inclus) (consulté le )
- (en) Référence Tree of Life Web Project : Embryophyta (consulté le )
- (en) Référence Paleobiology Database : Embryophyta Crane et al. 2004 (consulté le )
Bibliographie
- Gérard Cusset, Botanique: les embryophytes, Masson, .
Articles connexes
Liens externes
- Ressources relatives au vivant :
- (cs + en) BioLib
- (sv) Dyntaxa
- (en) Paleobiology Database
- (en) Système d'information taxonomique intégré
- Ressource relative à la santé :
- Jean-Yves Dubuission (professeur des Universités, Sorbonne université) et Sabine Hennequin (maître de conférences, Sorbonne université), « Embryophytes ou cormophytes ou archégoniates », sur Universalis.fr
- Pascal Gantet, Vincent Tandart et Alain Verger, « Les bryophytes : reproduction sexuée », sur le site de l'Université catholique de Lille
 Portail de la botanique
Portail de la botanique  Portail origine et évolution du vivant
Portail origine et évolution du vivant