Paraceratherium
Paraceratherium est un genre éteint de rhinocéros sans cornes figurant parmi les plus grands mammifères terrestres jamais découverts, ayant vécu durant la totalité de l'Oligocène, c'est-à-dire de 34 à 23 millions d'années avant notre ère, dans ce qui est aujourd'hui l'Eurasie. Les premiers fossiles ont été découverts dans ce qui est actuellement le Pakistan, avant que des restes supplémentaires ne soient signalés dans une zone géographique allant de la Chine aux Balkans. Le genre appartient à la famille également éteinte des paracérathériidés. Le nom générique du taxon signifie littéralement « près de la bête sans cornes », en référence à Aceratherium, le genre dans lequel l'espèce type, P. bugtiense, est placée à l'origine.
Cet article concerne l'animal. Pour l'article sur l'album portant l'un des noms synonymes du taxon, voir Baluchitherium (album).

| Règne | Animalia |
|---|---|
| Embranchement | Chordata |
| Sous-embr. | Vertebrata |
| Classe | Mammalia |
| Ordre | Perissodactyla |
| Super-famille | Rhinocerotoidea |
| Famille | † Paraceratheriidae |
Espèces de rang inférieur
- † P. bugtiense (espèce type) Pilgrim,
- † P. transouralicum Pavlova,
- † P. huangheense Li et al.,
- † P. linxiaense Deng et al.,
- † Baluchitherium Forster-Cooper,
- † Indricotherium Borissiak,
- † Pristinotherium Birkjukov,
- † Benaratherium ? Gabunia,
- P. bugtiense :
- Aceratherium bugtiense Pilgrim,
- Thaumastotherium osborni Forster-Cooper, (préoccupé)
- Baluchitherium osborni Forster-Cooper,
- Metamynodon bugtiensis Forster-Cooper,
- Paraceratherium zhajremensis Bayshashov,
- P. transouralicum :
- Indricotherium transouralicum Pavlova,
- Baluchitherium grangeri Osborn,
- Indricotherium asiaticum Borissiak,
- Indricotherium minus Borissiak,
- Indricotherium grangeri Osborn,
- Pristinotherium brevicervicale Birjukov,
- Noms incertains :
- Benaratherium callistratum Gabunia,
La taille exacte de Paraceratherium est inconnue en raison des fossiles incomplets, les estimations maximales vont de 4,8 mètres au garrot et une longueur d'environ 7,4 mètres, le tout pour un poids généralement estimé entre 15 et 20 tonnes. L'animal possède de grandes incisives en forme de défense et une incision nasale qui suggère qu'il aurait eu une lèvre supérieure ou une trompe préhensile. Les pattes de l'animal sont longues et ressemblent à des piliers et le cou allongé supporte un crâne mesurant environ 1,3 mètres de long. Le mode de vie de Paraceratherium peut avoir été similaire à celui des grands mammifères modernes tels que les éléphants ou les rhinocéros. Il aurait eu, comme ces derniers, un rythme de reproduction plutôt lent et un nombre réduit voire absent de prédateurs. C'est un brouteur dont l'alimentation est principalement composées de feuilles, de plantes herbacées et d'arbustes, vivant dans des habitats allant des déserts arides avec quelques arbres épars aux forêts subtropicales. Les raisons de l'extinction de cet animal sont inconnues, mais divers facteurs sont proposés.
La taxonomie des espèces de Paraceratherium est une histoire longue et complexe. D'autres genres d'indricothères datant également de l'Oligocène, tels que Baluchitherium, Indricotherium et Pristinotherium, ont été nommés, mais aucun spécimen complet n'a été découvert à ce jour, ce qui rend les comparaisons et les classifications difficiles. La plupart des scientifiques considèrent ces genres proposés comme des synonymes juniors de Paraceratherium et pensent qu'ils contiennent les espèces suivantes : P. bugtiense, P. transouralicum, P. huangheense et P. linxiaense. L'espèce la plus connue est P. transouralicum, la majorité des reconstitutions du genre étant basées sur cette dernière. Les différences entre P. bugtiense et P. transouralicum pourraient être dues à un possible dimorphisme sexuel, qui en ferait une même espèce.
Taxonomie
Historique des classifications
L'histoire taxonomique de Paraceratherium est complexe en raison de la nature fragmentaire des fossiles connus et du fait que les scientifiques occidentaux, soviétiques et chinois ont travaillés indépendamment les uns des autres pendant une grande partie du XXe siècle[1], leurs tentatives de comparer leurs découvertes pour obtenir une image plus complète de cet animal étant entravées par la politique et les conflits de l'époque[2]. De plus, ils publient leurs recherches dans leurs langues respectives[1]. Des tendances taxonomiques opposées mènent à des regroupements et des divisions successifs, ce qui a également contribué au problème[3]. Des datations géologiques inexactes ont auparavant conduit les scientifiques à croire que les formations géologiques, dont on sait aujourd'hui qu'elles sont contemporaines, seraient d'âges différents. De nombreux genres sont nommés sur la base de différences subtiles dans les caractéristiques des molaires qui ne sont pas acceptées par la plupart des scientifiques pour distinguer les espèces[note 1].
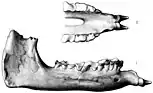
Les premières découvertes d'Indricothères sont faites par des colons occidentaux installés en Asie[2], les premiers fossiles connus étant récoltés au Baloutchistan, actuel Pakistan, en , par un soldat nommé « Vickary », mais ces fragments ne sont alors pas identifiables[5]. Les premiers fossiles maintenant reconnus comme appartenant à Paraceratherium sont découverts par le géologue et paléontologue britannique Guy Ellcock Pilgrim (en) au Baloutchistan, entre et . Son matériel consiste en une mâchoire supérieure, des dents inférieures ainsi que l'arrière d'une mandibule. Les fossiles sont récoltés dans la formation de Chitarwata du district de Dera Bugti, un lieu déjà exploré par Pilgrim auparavant. En , ce dernier les utilise comme base pour une nouvelle espèce du genre de rhinocéros éteint Aceratherium, à savoir A. bugtiense. Aceratherium est alors un taxon poubelle qui comprend plusieurs espèces non apparentées de rhinocéros sans cornes, dont beaucoup ont depuis été déplacées vers d'autres genres[1],[6]. Les incisives fossiles que Pilgrim a précédemment assignées au genre Bugtitherium s’avéreront appartenir à la même nouvelle espèce[7].
En , des fossiles plus partiels sont découverts dans le même endroit, lors d'une expédition du paléontologue britannique Clive Forster-Cooper (d). Sur la base de ces restes, Foster-Cooper déplace l'espèce A. bugtiense dans le nouveau genre Paraceratherium, signifiant littéralement « près de la bête sans cornes », en référence à Aceratherium, le nom de ce genre voulant littéralement dire « bête sans cornes »[1],[8]. Sa justification pour cette reclassification concerne les défenses inférieures de l'espèce qui sont nettement tournées vers le bas[7]. En , Forster-Cooper nomme un nouveau genre et une nouvelle espèce, Thaumastotherium osborni, sur la base de fossiles plus gros provenant des mêmes fouilles, dont il suggère plus tôt d'appartenir à un représentant mâle de l'espèce P. bugtiense. Il rebaptise par la suite le genre en Baluchitherium, l'ancien nom étant déjà utilisé, ce dernier nommant un insecte hémiptère[9],[10],[8]. Les fossiles de Baluchitherium sont si fragmentaires que Foster-Cooper l'identifie uniquement comme un périssodactyle indéterminé, tout en mentionnant la possibilité de confusion avec Paraceratherium[11]. Le paléontologue américain Henry Fairfield Osborn, d'après lequel B. osborni est nommé, suggère qu'il pourrait s'agir d'un potentiel brontothère[2].
Une expédition menée par l'Académie des sciences de l'URSS dans les années 1910 trouve par la suite des fossiles dans la formation d'Aral, près de la mer du même nom, au Kazakhstan : il s'agit d'un des squelettes d'Indricothères les plus complets connus, mais dont il lui manque le crâne. En , sur la base de ces restes, Aleksei Borissiak (ru) créé le genre Indricotherium en s'inspirant du nom d'un monstre du folklore russe, l'Indrik. Ce n'est qu'en qu'il lui attribue le nom d'espèce, I. asiaticum. Mais entre-temps, la paléontologue russe Maria Pavlova (en) l'a déjà nommé I. transouralicum en [1],[12]. Toujours en , Borissiak créé la sous-famille Indricotheriinae pour inclure les diverses formes apparentées connues à ce moment-là[13].
En , l'explorateur américain Roy Chapman Andrews mène une expédition bien documentée en Chine et en Mongolie, parrainée par le musée américain d'histoire naturelle. Divers restes d'indricothères sont trouvés dans des formations du désert de Gobi, incluant les pattes d'un spécimen debout et en position verticale, indiquant que l'animal serait mort piégé dans des sables mouvants, ainsi qu'un crâne très complet. Ces restes deviennent la base de sa description de Baluchitherium grangeri, qu'Osborn nomme officiellement en [14],[15].
En , l'espèce P. huangheense est nommée par le paléontologue chinois Yong-Xiang Li et son équipe sur la base d'une mâchoire inférieure provenant de la formation de Hanjiajing, située dans la province de Gansu, en Chine. Son épithète spécifique fait référence au nom chinois de la rivière à proximité de laquelle l'animal a été découvert : le fleuve Jaune[16]. En , le paléontologue chinois Deng Tao (en) et ses collègues décrivent la nouvelle espèce P. linxiaense, basée sur un crâne relativement complet associées d'une mandibule ainsi que des multiples vertèbres provenant de la formation Jiaozigou, située au nord-ouest de la Chine, plus précisément au sein du bassin de Linxia, d'où provient son nom[17]. Une multitude d'autres noms d'espèces et de genres, principalement basés sur des différences de taille, de forme de museau et de disposition des dents de devant, sont créés à partir de divers restes d'Indricothères. Des fossiles attribuables à Paraceratherium continuent d'être découverts dans toute l'Eurasie, notamment au Pakistan, mais en raison de la situation politique actuelle, il est difficile d'y effectuer de nouvelles fouilles[5].
Espèces et synonymes
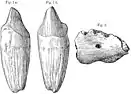
En , Forster-Cooper nomme la nouvelle espèce Metamynodon (en) bugtiensis en se basant sur un palais et d'autres fragments fossiles provenant du district de Dera Bugti, censés appartenir à un membre de grande taille de ce genre. Le chercheurs pense maintenant que ces fossiles appartenaient à un spécimen aberrant de P. bugtiense dépourvu de la troisième molaire[18],[19]. En , les paléontologues américains Walter Willis Granger et William King Gregory proposent que B. osborni de Forster-Cooper serait probablement un synonyme junior de P. bugtiense, car ces spécimens sont collectés dans la même localité et font peut-être partie d'une même espèce morphologiquement variable[20]. Le paléontologue américain William Diller Matthew et Forster-Cooper lui-même expriment des doutes similaires quelques années plus tôt. Bien qu'il soit déjà déclaré synonyme junior, le nom de genre Baluchitherium reste populaire dans divers médias en raison de la popularité entourant B. grangeri d'Osborn[3],[21].
En , les paléontologues américains Spencer G. Lucas (en) et Jay C. Sobus publient une révision des taxons d'indricothères, qui est suivie aujourd'hui par la plupart des scientifiques occidentaux. Ils concluent que Paraceratherium, incarnant nom le plus ancien proposé pour l'animal, serait le seul genre d'indricothères valide datant de l'Oligocène et contiendrait quatre espèces : P. bugtiense ; P. transouralicum, à l'origine classé dans Indricotherium ; P. prohorovi ; à l'origine classé dans Aralotherium (en) et P. orgosensis, à l'origine classé dans Dzungariotherium. Ils considèrent la plupart des autres noms comme des synonymes juniors de ces taxons, ou comme des noms douteux, basés sur des restes trop fragmentaires pour être correctement identifiés. En analysant les différences présumées entre les genres et les espèces nommés, Lucas et Sobus découvrent que celles-ci représentent très probablement des variations au sein des représentants et que la plupart des caractéristiques ne peuvent pas être distinguées entre les spécimens, comme cela avait été souligné dans les années 1930. Le fait que des crânes attribués à P. transouralicum soient bombé, tandis que d'autres sont plats au sommet est attribué à un probable dimorphisme sexuel : il est possible que les fossiles de P. bugtiense représentent des femelles, tandis que celles de P. transouralicum représentent des mâles au sein d'une même espèce[3],[22],[23].

Selon Lucas et Sobus, l'espèce type P. bugtiense, datant de la fin de l'Oligocène de l'actuel Pakistan, comprend des synonymes plus récents tels que B. osborni et P. zhajremensis. P. transouralicum, datant également de la fin de l'Oligocène des actuels Kazakhstan, Mongolie et du nord de la Chine, comprend B. grangeri et I. minus[22]. En , le paléontologue américain Donald Prothero (en) suggère que P. orgosensis peut être suffisamment distinct pour justifier son nom de genre d'origine Dzungariotherium, bien que sa position exacte nécessite une évaluation. P. prohorovi, datant de la fin de l'Oligocène du Kazakhstan est trop incomplet pour que sa position soit résolue par rapport aux autres espèces ; il en va de même pour des espèces proposées telles que I. intermedium et P. tienshanensis, ainsi que le genre Benaratherium[3],[22]. Bien que le nom de genre Indricotherium soit maintenant un synonyme junior de Paraceratherium, le nom de la sous-famille auquel il appartient dans certaines classifications, à savoir Indricotheriinae, est toujours utilisé parce que la synonymie du nom de genre n'affecte pas les noms des taxons de niveau supérieur qui en sont dérivés, ces derniers sont donc encore communément appelés indricothères[24].
Contrairement à la révision de Lucas et Sobus, un article de du paléontologue chinois Jie Ye et al. suggère qu'Indricotherium et Dzungariotherium sont des genres valides et que P. prohorovi n'appartient pas au genre Paraceratherium. Ils reconnaissent aussi la validité d'espèces telles que P. lipidus, P. tienshanensis et P. sui[25]. Un article de de Deng et de ses collègues reconnaissent également trois genres distincts[26]. Certains écrivains occidentaux utilisent de la même manière des noms autrement considérés comme invalides depuis la révision de , mais sans fournir d'analyse et de justification détaillées[3]. Deng et al. reconnait six espèces de Paraceratherium en , dont certains ont été auparavant déclarées synonymes tels que P. grangeri, P. asiaticum et P. lepidum, tout en gardant Indricotherium et Baluchitherium comme synonymes du genre[17].
Histoire évolutive
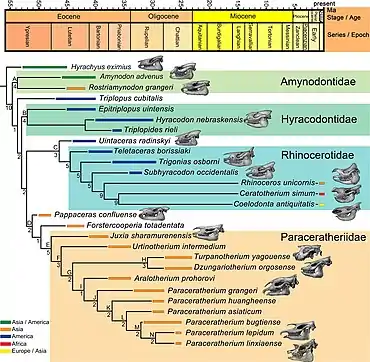
La super-famille des Rhinocerotoidea, qui comprend les rhinocéros modernes, apparaît durant le début de l'Éocène, vers environ 50 millions d'années avant notre ère, avec des précurseurs tels que Hyrachyus (en). Ce taxon contient trois familles officiellement reconnues : Amynodontidae ; Rhinocerotidae, dont font partie les rhinocéros actuels et Hyracodontidae. La diversité au sein du groupe de rhinocéros est beaucoup plus grande à l'époque préhistorique : ils passent de la taille d'un grand chien à celle d'un éléphant. Il y a des formes à longues pattes adaptées pour courir, ainsi que des formes semi-aquatiques. La plupart des espèces sont dépourvus de cornes. Les fossiles de rhinocéros sont identifiés principalement par les caractéristiques de leurs dents, qui sont la partie des animaux la plus susceptible d'être préservée au cours des fossilisations. Les molaires supérieures de la plupart des rhinocéros ont un motif en forme de pi (π) sur la couronne dentaire, et chaque molaire inférieure a des formes en « L » appariées. Diverses caractéristiques du crâne sont également utilisées pour l'identification des rhinocéros fossiles[27].
La sous-famille des Indricotheriinae, auquel appartient traditionnellement Paraceratherium, est classée pour la première fois dans la famille des Hyracodontidae par le paléontologue américain Leonard B. Radinsky (en) en . Ils sont auparavant considérés comme une sous-famille au sein des Rhinocerotoidea, voire une famille complète, les Indricotheriidae[28]. Dans une étude cladistique de sur les tapiromorphes, le paléontologue américain Luke Holbrook découvre que les indricothères sont en dehors des hyracodontidés et écrit qu'ils ne peuvent pas être un regroupement naturel[29]. Le schéma de Radinsky est l'hypothèse qui prévaut de nos jours. La famille des hyracodontidés contient des membres à longues pattes adaptés à courir, tels qu'Hyracodon, et se distinguent par des caractéristiques d'incisives. Les indricothères se distinguent des autres hyracodontes par leur plus grande taille et la structure dérivée de leurs museaux, incisives et canines. Le plus ancien indricothère connu est Forstercooperia (en), un animal de la taille d'un grand chien ayant vécu durant l'Éocène moyen et supérieur de l'ouest des actuelles Amérique du Nord et Asie. Juxia est de la taille d'une vache et est connue depuis l'Éocène moyen. Le genre Urtinotherium, connu d'Asie de la fin de l'Éocène, atteint presque la taille de Paraceratherium[22],[27]. Paraceratherium lui-même est connu d'Eurasie durant l'Oligocène, sur une période allant de 34 à 23 millions d'années[30]. Le genre se distingue des autres indricothères par sa grande taille, une incision nasale qui aurait soutenu un museau musclé et ses prémaxillaires renversés[22]. L'animal est également dépourvue des deuxièmes et troisièmes incisives, des canines et des premières prémolaires inférieures[27].
Le cladogramme ci-dessous suit l'analyse de des Indricotheriinae par Lucas et Sobus, et montre les plus proches parents de Paraceratherium[22] :
| ◄ Hyracodontidae |
| ||||||||||||||||||||||||
Lucas et ses collègues parviennent à des conclusions similaires dans une analyse précédente de Forstercooperia en , dans laquelle ils conservent toujours Paraceratherium et Indricotherium comme des genres distincts[31]. En , les chercheurs chinois Haibing Wang et ses collègues utilisent le nom Paraceratheriidae pour la famille et Paraceratheriinae pour la sous-famille, et les placent en dehors des Hyracodontidae[32]. Deng et ses collègues découvrent que leur analyse de confirme les études antérieures, suggérant que Juxia aurait évolué à partir d'un clade composé de Forstercooperia et Pappaceras (en), vers environ 40 millions d'années avant notre ère, le stock résultant évoluant en Urtinotherium à la fin de l'Éocène et Paraceratherium à l'Oligocène. Ces chercheurs ne considère pas que les hyracodontidés forment un groupe naturel et classent les paracérathériidés comme étant plus proches des rhinocéros actuels, contrairement aux études précédentes[17].
Description
Morphologie

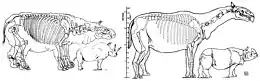
Paraceratherium est l'un des plus grands mammifères terrestres connus qui aient jamais existé, bien que sa taille précise soit incertaine en raison du manque de spécimens complets[4]. La longueur totale du corps est estimée à 8,7 mètres de la tête à la queue par Granger et Gregory en , puis rectifiée à 7,4 mètres par la paléontologue soviétique Véra Gromova en , la première estimation étant maintenant considéré comme exagérée. Le poids de Paraceratherium serait similaire à celui de certains proboscidiens aujourd'hui éteints, le plus grand squelette complet de cette ordre étant celui d'un mammouth des steppes[33],[34]. Malgré sa masse à peu près équivalente, Paraceratherium serait plus grand que n'importe quel proboscidien[4]. Sa hauteur au garrot est estimée à 5,25 mètres par Granger et Gregory, puis par 4,8 mètres par le paléontologue américain Gregory S. Paul en [35]. Le cou est estimé entre 2 et 2,5 mètres de long par les paléontologues Michael P. Taylor (en) et Mathew J. Wedel (en) en [36].
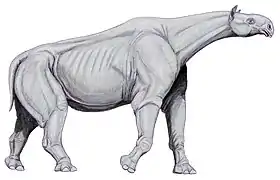
Les premières estimations de 30 tonnes sont maintenant considérées comme exagérés, le maximum admis aujourd'hui étant entre 15 et 20 tonnes pour une moyenne de 11 tonnes. Les calculs sont principalement basés sur des fossiles de P. transouralicum car cette espèce est connue par les restes les plus complets[4]. Les estimations s'appuient sur les mesures des os du crâne, des dents et des membres, mais les éléments osseux connus sont représentés par des individus de différentes tailles, de sorte que toutes les reconstructions squelettiques sont des extrapolations composites, résultant en plusieurs poids différents[33],[37].
Il n'y a aucune indication sur la couleur et la texture de peau de Paraceratherium, aucune empreinte cutanée ou momies n'étant connues. La plupart des reconstitutions proposent une peau épaisse, pliée, grise et glabre, inspirée des rhinocéros modernes. Les grands mammifères modernes tels que les éléphants et les rhinocéros sont en grande partie glabres, car la fourrure retient la chaleur corporelle. Prothero suggère aussi que, contrairement à la plupart des représentations, Paraceratherium aurait eu de grandes oreilles semblables à celle des éléphants, qu'il utiliserait pour la thermorégulation. Les oreilles des éléphants agrandissent la surface corporelle et sont remplies de vaisseaux sanguins, ce qui facilite la dissipation de l'excès de chaleur. Selon Prothero, les os robustes autour des ouvertures des oreilles sont un indice confirmant cette théorie[4]. Les paléontologues Pierre-Olivier Antoine et Darren Naish (en) expriment leur scepticisme à l'égard de cette proposition[38],[39].
En raison de la nature fragmentaire des fossiles connus de Paraceratherium, le squelette de l'animal est reconstruit de plusieurs manières différentes depuis sa découverte[40]. En , Matthew supervise un artiste pour dessiner une reconstruction du squelette sur la base des spécimens encore moins complets de P. transouralicum connus durant l'époque, en utilisant les proportions d'un rhinocéros moderne comme guide[41]. Le résultat étant trop trapu et compact, Osborn propose donc une version plus élancée plus tard la même année. Certaines reconstitutions ultérieures rendent l'animal trop mince, sans en tenir compte du squelette sous-jacent[4]. Gromova publie une reconstruction squelettique plus complète en , fondée sur le squelette d'un P. transouralicum de la formation d'Aral, auquel il manque cependant plusieurs vertèbres cervicales[42].
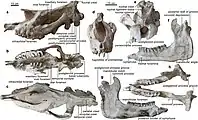
Crâne
Les plus grands crânes connus de Paraceratherium mesurent environ 1,3 mètres de long, 33 à 38 centimètres à l'arrière du crâne et 61 centimètres de large à travers les arcs zygomatiques. Paraceratherium possède un long front, lisse et dépourvu de la zone rugueuse qui sert de point d'attache pour les cornes des autres rhinocéros. Les os au-dessus de la région nasale sont longs et l'incision nasale va loin dans le crâne. Cela indique que Paraceratherium aurait eu une lèvre supérieure préhensile similaire à celle du rhinocéros noir et du rhinocéros indien, ou une courte trompe comme chez les tapirs[4]. De plus, une caractéristique distinctive de Paraceratherium est que l'incision nasale est rétractée vers les deuxième et troisième prémolaires[17] .
L'arrière du crâne est bas et étroit, sans les grandes crêtes lambdoïdes au sommet et le long de la crête sagittale, que l'on trouve par ailleurs chez les animaux à cornes et à défenses qui ont besoin de muscles puissants pour pousser et se battre. Il y a aussi une fosse profonde pour la fixation des ligaments nucaux, qui retiennent automatiquement le crâne. Le condyle occipital est très large et Paraceratherium semble avoir eu de gros et puissants muscles du cou, ce qui lui permettait de balayer fortement sa tête vers le bas tout en se nourrissant de branches d'arbres[4]. Le profil supérieur du crâne est arqué, c'est-à-dire courbé en arc, ce qui constitue une caractéristique distinctive du genre[17]. Le crâne de P. transouralicum a un front bombé, tandis que d'autres ont un front plat, probablement à cause d'un possible dimorphisme sexuel[22]. Un endocaste du cerveau de P. transouralicum montre qu'il ne représente que 8 % de la longueur du crâne, tandis que le cerveau du rhinocéros indien représente 17,7 % de la longueur de son crâne[20].
Les espèces de Paraceratherium sont principalement discernables à travers les caractéristiques du crâne. P. bugtiense se distingue par des maxillaires et des prémaxillaires relativement minces, des toits crâniens peu profonds, des processus mastoïdo-paroccipitaux relativement minces et placés en arrière sur le crâne, une crête lambdoïde qui s'étend moins en arrière et un condyle occipital avec une orientation horizontale, qu'il partage avec le genre apparenté Dzungariotherium. P. transouralicum a des maxillaires et des prémaxillaires robustes, des os zygomatiques renversés, des os frontaux en forme de dôme, des processus mastoïdo-paroccipitaux épais, une crête lambdoïde qui s'étend vers l'arrière et des condyles occipitaux avec une orientation verticale[3]. P. huangheense ne diffère de P. bugtiense que par l'anatomie de la partie arrière de la mâchoire, ainsi que par sa plus grande taille[16]. P. linxiaense diffère des autres espèces en ce que l'encoche nasale est plus profonde, avec le fond placé au-dessus du milieu de la deuxième molaire, un condyle occipital proportionnellement plus élevé par rapport à la hauteur de la surface occipitale, des os du museau courts, un diastème devant les dents jugales, un arc zygomatique haut avec une extrémité postérieure proéminente ainsi qu'une première incisive supérieure plus petite[17].
Contrairement à celles de la plupart des rhinocéros primitifs, les dents de devant de Paraceratherium sont réduites à une seule paire d'incisives, grandes et coniques, dans chaque mâchoire. Celles-ci sont décrites comme des défenses. Les incisives supérieures pointent vers le bas et les inférieures sont plus courtes et pointent vers l'avant. Parmi les rhinocéros connus, cet arrangement est unique à Paraceratherium ainsi qu'au genre apparenté Urtinotherium et les incisives peuvent avoir été plus grandes chez les mâles. Par ailleurs, ces canines trouvées derrière les incisives ont été perdues. Les incisives sont séparées de la rangée de dents jugales par un grand diastème[4]. Cette caractéristique se trouve chez les mammifères où les incisives et les dents jugales ont des spécialisations différentes[27]. Les molaires supérieures, à l'exception de la troisième molaire supérieure qui est en forme de « V », ont un motif en forme de pi (π) et un métastyle réduit[note 2]. Les prémolaires ne forment que partiellement le motif pi et chaque molaire a la taille d'un poing humain[note 3]. Les dents de la joue inférieure sont en forme de « L », ce qui est typique des rhinocéros[4].
Squelette postcrânien
Aucun ensemble complet de vertèbres et de côtes de Paraceratherium n'a encore été trouvé et la queue est complètement inconnue. L'atlas et les vertèbres de l'axe du cou sont plus larges que chez la plupart des rhinocéros modernes, avec un espace pour les ligaments et les forts muscles qui seraient nécessaires pour soutenir la tête. Le reste des vertèbres sont également très large et ont de grandes zygapophyses avec beaucoup de place pour les muscles, les tendons, les ligaments, les nerfs, le cou ainsi que la colonne vertébrale. Les épines neurales sont longues et forment une longue « bosse » le long du dos, où sont attachés les muscles du cou et les ligaments nucaux pour soutenir le crâne. Les côtes sont similaires à celles des rhinocéros modernes, mais la cage thoracique semble être plus petite par rapport aux longues pattes et aux grands corps, car les rhinocéros modernes ont des membres relativement courts. La dernière vertèbre du bas du dos est fusionnée au sacrum, une caractéristique trouvable chez les rhinocéros avancés[4]. Tout comme les dinosaures sauropodes, Paraceratherium possède des parties creuses des os dans les vertèbres pré-sacrées, qui aurait probablement aidé à alléger le squelette[43].
Les membres sont grands et robustes pour supporter le poids important de l'animal, ces derniers étant à certains égards similaires et convergents avec ceux des éléphants et des dinosaures sauropodes avec leurs morphologies également lourdes. Contrairement à ces animaux, qui ont tendance à allonger les os des membres supérieurs tout en raccourcissant, fusionnant et comprimant les os des membres inférieurs, des mains et des pieds, Paraceratherium a des courts os des membres supérieurs et des longs os des mains et des pieds, à l'exception des phalanges en forme de disque, semblable aux rhinocéros coureurs antérieurs dont il en descend. Certains os du pied mesurent près de 50 centimètres de long. Les os de la cuisse mesurent généralement 1,5 mètres, une taille dépassée seulement par ceux de certains éléphants et dinosaures. Les os des cuisses sont en forme de piliers et beaucoup plus épais et plus robustes que ceux des autres rhinocéros, et les trois trochanters sur les côtés sont très réduits, car cette robustesse diminue leurs importances. Les membres sont maintenus dans une posture en forme de colonne au lieu d'être pliés, comme chez les petits animaux, ce qui réduit le besoin de gros muscles des membres[4]. Comme la plupart des périssodactyles, les membres antérieurs ont trois orteils[44].
Paléobiologie
Mode de vie
Le zoologiste britannique Robert M. Alexander (en) suggère en que la surchauffe pouvait avoir été un sérieux problème chez Paraceratherium en raison de sa taille[45]. Selon Prothero, les meilleurs analogues vivants de Paraceratherium pourraient être de grands mammifères tels que les éléphants, les rhinocéros et les hippopotames. Pour aider à la thermorégulation, ces animaux se rafraîchissent pendant la journée en se reposant à l'ombre ou en se trempant dans l'eau ou la boue. Ils se nourrissent et se déplacent principalement la nuit. En raison de la grande taille de Paraceratherium, l'animal n'aurait pas pu courir et se déplacer rapidement, mais il aurait pu parcourir de grandes distances, ce qui serait nécessaire dans un environnement où la nourriture est rare. Il peut donc avoir eu de grands domaines vitaux et serait migrateur[4]. Prothero suggère que des animaux aussi gros que les indricothères auraient besoin de très grands domaines vitaux ou de territoires d'au moins 1 000 kilomètres carrés et qu'en raison de la rareté des ressources, il y aurait eu peu de place en Asie pour de nombreuses populations ou une multitude d'espèces et de genres presque identiques. Ce principe est appelé principe de Gause : il est utilisé pour expliquer comment le rhinocéros noir et le rhinocéros blanc exploitent des niches différentes dans les mêmes régions d'Afrique[3].
La plupart des prédateurs de leurs habitats ne sont pas plus gros qu'un loup et ne constitueraient pas une menace pour Paraceratherium, les individus adultes seraient trop gros pour être attaqués par ces derniers, mais les jeunes auraient été probablement vulnérables[30]. Des marques de morsures sur les os trouvés dans les lits de Dera Bugti, au Pakistan, indiquent que même les adultes peuvent avoir été la proie d'un crocodilien long de 10 à 11 mètres de long, Astorgosuchus (en). Comme chez les éléphants, la période de gestation de Paraceratherium peut avoir été longue et les individus peuvent avoir eu une longue durée de vie[4]. L'animal pourrait avoir vécu en petits troupeaux, peut-être constitués de femelles et de leurs juvéniles qu'elles protégeraient des potentiels prédateurs[30]. Il est proposé que 20 tonnes pourraient être le poids maximum possible pour les mammifères terrestres, et que Paraceratherium est proche de cette limite[46]. Les raisons pour lesquelles les mammifères ne peuvent pas atteindre une taille similaire à celles des grands dinosaures sauropodes sont inconnues, les facteurs étant peut-être écologiques plutôt que biomécanique, et peut-être liée aux stratégies de reproduction[33]. Le mouvement, le son et d'autres comportements vus dans les documentaires en images de synthèse tels que Sur la terre des monstres disparus sont entièrement hypothétiques[4].
Régime et alimentation

Les dents simples et à couronne basse indiquent que Paraceratherium est un brouteur avec un régime composé de feuilles et d'arbustes relativement mous. Les rhinocéros ultérieurs sont des brouteurs, avec des dents hautes car leur régime alimentaire contient du gravier qui use rapidement leurs dents. Des études basées sur les dents (en) de Paraceratherium confirment que ces créatures ont un régime à base de feuilles molles[note 4]. L'analyse isotopique montre que Paraceratherium se nourrissait principalement de plantes au carbone C3, qui sont principalement des feuilles d'arbres[47],[48]. Comme ses parents périssodactyles (équidés, tapirs et autres rhinocéros), Paraceratherium aurait été un fermenteur d'intestin postérieur : il extrairait relativement peu de nutriments de sa nourriture et devrait en manger de grandes quantités pour survivre. Comme les autres herbivores de grande taille, l'animal aurait eu un gros tube digestif[4].
Granger et Gregory soutiennent que les grandes incisives seraient utilisées comme des défenses ou pour desserrer les arbustes en déplaçant le cou vers le bas, agissant ainsi comme des pioches et des leviers[20]. Les tapirs utilisent leur trompe pour envelopper les branches tout en enlevant l'écorce avec les incisives, capacité qui aurait été utile à Paraceratherium. Certains auteurs russes suggèrent que les défenses seraient probablement utilisées pour casser les brindilles, arracher l'écorce et plier les hautes branches et qu'en raison du fait que les espèces du début de l'Oligocène auraient des défenses plus grosses que les dernières, elles avaient probablement un régime plus basé sur l'écorce que sur les feuilles. Étant donné que les espèces impliquées sont maintenant connues pour avoir été contemporaines et que les différences de défenses sont maintenant considérées comme sexuellement dimorphes, cette dernière idée n'est désormais plus acceptée[4]. Des troupeaux de Paraceratherium peuvent avoir migré tout en se nourrissant continuellement des feuilles provenant de grands arbres que les mammifères plus petits ne peuvent pas atteindre[30]. Osborn suggère que son mode de recherche de nourriture aurait été similaire à celui des giraffidés plutôt qu'aux rhinocéros modernes, dont les têtes sont portées près du sol[41].
Répartition géographique
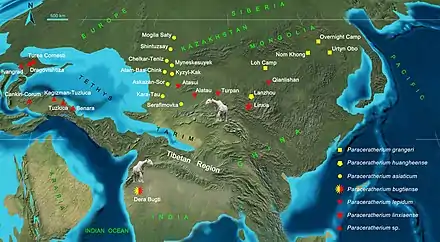
Des restes attribuables à Paraceratherium sont découverts dans des formations datant du début à la fin de l'Oligocène à travers l'Eurasie dans les actuels Chine, Mongolie, Inde, Pakistan, Kazakhstan, Géorgie, Turquie, Roumanie, Bulgarie et Balkans[5]. Leur distribution peut être corrélée avec le développement paléogéographique de l'Himalaya. L'aire de répartition des découvertes de Paraceratherium implique qu'ils habitaient une masse continentale continue avec un environnement similaire à travers elle, mais cela est contredit par les cartes paléogéographiques qui montrent que cette zone avait diverses barrières marines, de sorte que le genre a réussi à être largement distribué malgré cela[49]. La faune qui coexiste avec Paraceratherium comprend d'autres rhinocéros, des artiodactyles, des rongeurs, des chiens-ours, des belettes, des hyaenodontes, des nimravidés ainsi que des félins[30].
L'habitat de Paraceratherium semble varier dans son aire de répartition, en fonction des types de formations géologiques dans lesquelles il est découvert[30]. La formation mongolienne de Hsanda Gol (en) représente un bassin désertique aride et l'environnement aurait eu peu de grands arbres et une couverture de broussailles limitée, car la faune se compose principalement d'animaux qui se nourrissent de la cime des arbres ou près du sol[50]. Une étude basée sur du pollen fossile montre qu'une grande partie de la Chine était constituée de broussailles ligneuses, avec des plantes telles que Atriplex, Ephedra et Nitraria (en), tous adaptés aux milieux arides. Les arbres étaient rares et concentrés près des eaux souterraines[51]. Les régions de la Chine où vivait Paraceratherium avaient des lacs asséchés et des dunes de sables abondants, les fossiles de végétaux les plus courants sont les feuilles adaptées à ce type de paysage appartenant au genre Palibinia. Les arbres de Mongolie et de Chine comprennent des bouleaux, des ormes, des chênes et d'autres arbres à feuilles caduques, tandis que celles la Sibérie et du Kazakhstan ont également des noyers[30]. Le district de Dera Bugti de l'actuel Pakistan avait une forêt sèche, tempérée à subtropicale[47].
Deng et ses collègues spéculent sur la paléobiogéographie de Paraceratherium sur la base de leur analyse phylogénétique en : ils découvre que P. bugtiense est la seule espèce du genre représentée dans l'Oligocène de l'ouest du Pakistan, tandis que le genre est très diversifié sur le plateau mongol, au nord-ouest de la Chine, le nord du Kazakhstan jusqu'au plateau tibétain. Ils émettent l'hypothèse que P. asiaticum se serait dispersé vers l'ouest jusqu'au Kazakhstan au début de l'Oligocène à partir de la région ancestrale de la Mongolie, où vivait le membre le plus primitif du genre, P. grangeri, et que ses descendants pourraient avoir continué jusqu'en Asie du Sud sous le nom de P. bugtiense, se dispersant dans la région tibétaine. P. lepidum existait au Xinjiang et au Kazakhstan et P. linxiaense à Linxia, en Chine, à la fin de l'Oligocène, et il est possible que ces espèces sœurs de P. bugtiense aient pu migrer vers le nord en Asie centrale pendant la période où cette zone était devenue tropicale, étant aride au début de l'Oligocène. Cela implique que la région tibétaine n'était pas encore un plateau de haute altitude qui pourrait servir de barrière, et les grands animaux peuvent donc avoir pu se déplacer librement le long de la côte orientale de la mer de Téthys, et à travers les basses terres de la région, dont certains seraient peut-être en dessous de 2 000 mètres d'altitude lors de cette époque[17].
Extinction
Les raisons pour lesquelles Paraceratherium s'est éteint après avoir survécu pendant environ 11 millions d'années sont inconnues, mais il est peu probable qu'il y ait eu une cause unique[30]. Les théories incluent un changement climatique, un faible taux de reproduction ainsi que l'arrivée des gomphothères et d'autres herbivores depuis l'Afrique à la fin de l'Oligocène, entre 28 et 23 millions d'années avant notre ère[30]. Les nouveaux herbivores ont peut-être pu considérablement modifier les habitats dans lesquels ils sont entrés, de la même manière que les éléphants d'Afrique le font aujourd'hui, c'est-à-dire en détruisant les arbres et en transformant les forêts en prairies. Une fois que leurs sources de nourriture se sont raréfiés et que leur nombre à diminué, les populations de Paraceratherium seraient devenues plus vulnérables à d'autres menaces[52]. Des grands prédateurs comme Hyainailurus ou Amphicyon sont également entrés en Asie depuis l'Afrique au début du Miocène, ceux-ci peuvent s'être attaqués aux juvéniles de Paraceratherium ou à d'autres indricothères[30].
Culture populaire
.png.webp)
Depuis sa découverte, Paraceratherium est l'objet de plusieurs représentations au sein des médias en raison de sa grande taille, notamment dans des documentaires et même dans plusieurs œuvres de fiction, dont quelques exemples ci-dessous :
- des Paraceratherium apparaissent dans le jeu vidéo Ark: Survival Evolved où ils peuvent être domestiqués par les joueurs[53] ;
- un groupe de Paraceratherium, identifié sous le nom d'Indricotherium, apparaît dans le troisième épisode de la série-documentaire Sur la terre des monstres disparus[54] ;
- Paraceratherium figure parmi les principaux animaux du documentaire produit par France 5, Le mystère des géants disparus[55] ;
- le documentaire produit par la National Geographic intitulé de manière inapproprié Les monstres du Jurassique fait figurer un Paraceratherium parmi les dix principaux animaux de grande taille du long-métrage[56] ;
- le directeur des effets spéciaux Phil Tippett affirme s'être servi de Paraceratherium comme source d'inspiration pour la création des véhicules TB-TT qui apparaissent dans la franchise Star Wars depuis le film L'Empire contre-attaque, sorti en [57]. A noter que certaines créatures apparaissant dans le jeu vidéo Star Wars Episode I : Le Nouveau Monde Gungan, s'inspirent également de Paraceratherium pour leur morphologie[58].
Notes et références
Notes
- Ces refus proviennent du fait que ces caractéristiques peuvent varier au sein des populations d'une même espèce de rhinocéros[4].
- Un métastyle est une crête recouverte d'émail sur la face externe d'une molaire chez certains périssodactyles.
- Chez les autres mammifères, ces molaires ne sont dépassées en taille que chez les proboscidiens.
- Des études sur la micro-usure des dents de Paraceratherium n'ont pas encore été menées[4].
Références
- Prothero 2013, p. 17-34.
- (en) C. Manias, « Building Baluchitherium and Indricotherium: Imperial and International Networks in Early-Twentieth Century Paleontology », Journal of the History of Biology, vol. 48, no 2, , p. 237–78 (PMID 25537636, DOI 10.1007/s10739-014-9395-y, S2CID 207150574, lire en ligne)
- Prothero 2013, p. 67-86.
- Prothero 2013, p. 87-106.
- Prothero 2013, p. 35-52.
- (en) G. E. Pilgrim, « Notices of new mammalian genera and species from the Tertiaries of India », Records of the Geological Survey of India, vol. 40, no 1, , p. 63–71
- (en) C. F. Cooper, « On the skull and dentition of Paraceratherium bugtiense: a genus of aberrant rhinoceroses from the Lower Miocene deposits of Dera Bugti », Philosophical Transactions of the Royal Society B: Biological Sciences, vol. 212, nos 391–401, , p. 369–394 (DOI 10.1098/rstb.1924.0009
 , Bibcode 1924RSPTB.212..369F)
, Bibcode 1924RSPTB.212..369F) - (en) C. Forster-Cooper, « LXXVIII.—Paraceratherium bugtiense, a new genus of Rhinocerotidae from the Bugti Hills of Baluchistan.—Preliminary notice », Annals and Magazine of Natural History, vol. 8, no 48, , p. 711–716 (DOI 10.1080/00222931108693085, lire en ligne [archive du ], consulté le )
- (en) C. Forster-Cooper, « XLIV.— Thaumastotherium osborni, a new genus of perissodactyles from the Upper Oligocene deposits of the Bugti hills of Baluchistan. —Preliminary notice », Annals and Magazine of Natural History, vol. 12, no 70, , p. 376–381 (DOI 10.1080/00222931308693412, lire en ligne [archive du ], consulté le )
- (en) C. Forster-Cooper, « Correction of generic name », Annals and Magazine of Natural History, vol. 12, no 71, , p. 504 (DOI 10.1080/00222931308693431, lire en ligne [archive du ], consulté le )
- (en) C. Forster-Cooper, « Baluchitherium osborni (? syn. Indricotherium turgaicum, Borrissyak) », Philosophical Transactions of the Royal Society of London, vol. 212, nos 391–401, , p. 35–66 (DOI 10.1098/rstb.1924.0002 , JSTOR 92060)
- M. Pavlova, « Indricotherium transouralicum n. sp. provenant du district de Tourgay », Bulletin de la Société des Naturalistes de Moscou, Section Géologique, vol. 31, , p. 95–116
- (de) A. A. Borissiak, « Über die Unterfamilie Indricotheriinae Boriss. = Baluchitheriinae Osb », Zentralblatt für Mineralogie, Geologie und Paläontologie, vol. 18, , p. 571–575
- Prothero 2013, p. 1-16.
- (en) H. F. Osborn, « The extinct giant rhinoceros Baluchitherium of Western and Central Asia », Natural History, vol. 23, , p. 208–228 (lire en ligne [archive du ], consulté le )
- (zh + en) Yong-Xiang, L., Yun-Xiang, Z., Ji, L., Zhi-Chao, L. et Kun, X., « New fossils of paraceratheres (Perissodactyla, Mammalia) from the Early Oligocene of the Lanzhou Basin, Gansu Province, China », Vertebrata PalAsiatica, vol. 55, no 4, , p. 367–381 (DOI 10.19615/j.cnki.1000-3118.170922, lire en ligne [archive du ], consulté le )
- (en) T. Deng, X. Lu, S. Wang, L. J. Flynn, D. Sun, W. He et S. Chen, « An Oligocene giant rhino provides insights into Paraceratherium evolution », Communications Biology, vol. 4, no 1, , p. 639 (PMID 34140631, PMCID 8211792, DOI 10.1038/s42003-021-02170-6 )
- (en) C. Forster-Cooper, « LXXIV.—Metamynodon bugtiensis, sp. n., from the Dera Bugti deposits of Baluchistan.—preliminary notice », Annals and Magazine of Natural History, vol. 9, no 53, , p. 617–620 (DOI 10.1080/00222932208632717, lire en ligne)
- (en) J. Tissier, D. Becker, V. Codrea, L. Costeur, C. Fărcaș, A. Solomon, M. Venczel, O. Maridet et T. Smith, « New data on Amynodontidae (Mammalia, Perissodactyla) from Eastern Europe: Phylogenetic and palaeobiogeographic implications around the Eocene-Oligocene transition », PLOS ONE, vol. 13, no 4, , e0193774 (PMID 29668673, PMCID 5905962, DOI 10.1371/journal.pone.0193774 , Bibcode 2018PLoSO..1393774T)
- (en) W. Granger et W. K. Gregory, « Further notes on the gigantic extinct rhinoceros, Baluchitherium, from the Oligocene of Mongolia », Bulletin of the American Museum of Natural History, vol. 72, , p. 1–73 (hdl 2246/363)
- (en) C. Forster-Cooper, « The extinct rhinoceroses of Baluchistan », Philosophical Transactions of the Royal Society B: Biological Sciences, vol. 223, nos 494–508, , p. 569–616 (DOI 10.1098/rstb.1934.0013 , Bibcode 1934RSPTB.223..569F)
- (en) S. G. Lucas et J. C. Sobus, The Evolution of Perissodactyls, New York, New York & Oxford, England, Oxford University Press, , 358–378 p. (ISBN 978-0-19-506039-3, OCLC 19268080), « The systematics of indricotheres »
- (zh + en) Q. Zhan-Xiang, « A new genus of giant rhinoceros from oligocene of Dzungaria, Sinkang », Vertebrata PalAsiatica, vol. 2, , p. 182–191 (lire en ligne [archive du ], consulté le )
- (en) P. O. Antoine, L. Karadenizli, G. E. Saraç et S. Sen, « A giant rhinocerotoid (Mammalia, Perissodactyla) from the Late Oligocene of north-central Anatolia (Turkey) », Zoological Journal of the Linnean Society, vol. 152, no 3, , p. 581–592 (DOI 10.1111/j.1096-3642.2007.00366.x )
- (zh + en) Y. Ye, Meng J. et W. W. Yu, « Discovery of Paraceratherium in the northern Junggar Basin of Xinjiang », Vertebrata PalAsiatica, vol. 41, , p. 220–229 (lire en ligne [archive du ], consulté le )
- (zh + en) Z.-X. Qui, B.-Y. Wang et T. Deng, « Indricotheres (Perissodactyla, Mammalia) from Oligocene in Linxia Basin, Gansu, China », Vertebrata PalAsiatica, vol. 42, no 3, , p. 177–192 (lire en ligne [archive du ], consulté le )
- Prothero 2013, p. 53-66.
- (en) L. B. Radinsky, « The families of the Rhinocerotoidea (Mammalia, Perissodactyla) », Journal of Mammalogy, vol. 47, no 4, , p. 631–639 (DOI 10.2307/1377893, JSTOR 1377893)
- (en) L. Holbrook, « The phylogeny and classification of tapiromorph perissodactyls (Mammalia) », Cladistics, vol. 15, no 3, , p. 331–350 (DOI 10.1006/clad.1999.0107)
- Prothero 2013, p. 107-121.
- (en) S. G. Lucas, R. M. Schoch et E. Manning, « The systematics of Forstercooperia, a Middle to Late Eocene hyracodontid (Perissodactyla: Rhinocerotoidea) from Asia and Western North America », Journal of Paleontology, vol. 55, no 4, , p. 826–841 (JSTOR 1304430)
- (en) H. Wang, B. Bai, J. Meng et Y. Wang, « Earliest known unequivocal rhinocerotoid sheds new light on the origin of Giant Rhinos and phylogeny of early rhinocerotoids », Scientific Reports, vol. 6, no 1, , p. 39607 (PMID 28000789, PMCID 5175171, DOI 10.1038/srep39607 , Bibcode 2016NatSR...639607W)
- (en) M. Fortelius et J. Kappelman, « The largest land mammal ever imagined », Zoological Journal of the Linnean Society, vol. 108, , p. 85–101 (DOI 10.1111/j.1096-3642.1993.tb02560.x)
- (en) A. Larramendi, « Shoulder height, body mass and shape of proboscideans », Acta Palaeontologica Polonica, vol. 61, (DOI 10.4202/app.00136.2014, S2CID 2092950, lire en ligne [archive du ])
- (en) G. S. Paul, « Dinosaur models: The good, the bad, and using them to estimate the mass of dinosaurs », Dinofest International Proceedings, , p. 129–142 (lire en ligne [archive du ], consulté le )
- (en) M. P. Taylor et M. J. Wedel, « Why sauropods had long necks; and why giraffes have short necks », PeerJ, vol. 1, , e36 (PMID 23638372, PMCID 3628838, DOI 10.7717/peerj.36 )
- (en) T. Tsubamoto, « Estimating body mass from the astragalus in mammals », Acta Palaeontologica Polonica, , p. 259–265 (DOI 10.4202/app.2011.0067 , S2CID 54686160)
- (en) P. O. Antoine, « There were giants upon the earth in those days (Book Review) », Palaeovertebrata, vol. 38, , p. 1–3 (DOI 10.18563/pv.38.1.e4, lire en ligne [archive du ], consulté le )
- (en) Naish, D., « Tet Zoo Bookshelf: van Grouw's Unfeathered Bird, Bodio's Eternity of Eagles, Witton's Pterosaurs, Van Duzer's Sea Monsters on Medieval and Renaissance Maps! » [archive du ], blogs.scientificamerican.com/tetrapod-zoology, (consulté le )
- (en) W. Granger et W. K. Gregory, « A revised restoration of the skeleton of Baluchitherium, gigantic fossil rhinoceros of Central Asia », American Museum Novitates, no 787, , p. 1–3 (hdl 2246/2123)
- (en) H. F. Osborn, « Baluchitherium grangeri, a giant hornless rhinoceros from Mongolia », American Museum Novitates, no 78, , p. 1–15 (hdl 2246/3262)
- (ru) V. L. Gromova, « Gigantskie nosorogi », Trudy Paleontology Institut Akademii Nauk SSSR, vol. 71, , p. 154–156
- (en) P. M. Sander, A. Christian, M. Clauss, R. Fechner, C. T. Gee, E. M. Griebeler, H. C. Gunga, J. R. Hummel, H. Mallison, S. F. Perry, H. Preuschoft, O. W. M. Rauhut, K. Remes, T. Tütken, O. Wings et U. Witzel, « Biology of the sauropod dinosaurs: The evolution of gigantism », Biological Reviews, vol. 86, no 1, , p. 117–155 (PMID 21251189, PMCID 3045712, DOI 10.1111/j.1469-185X.2010.00137.x )
- (en) P. O. Antoine, S. M. Ibrahim Shah, I. U. Cheema, J. Y. Crochet, D. D. Franceschi, L. Marivaux, G. G. Métais et J. L. Welcomme, « New remains of the baluchithere Paraceratherium bugtiense from the Late/latest Oligocene of the Bugti hills, Balochistan, Pakistan », Journal of Asian Earth Sciences, vol. 24, no 1, , p. 71–77 (DOI 10.1016/j.jaes.2003.09.005, Bibcode 2004JAESc..24...71A)
- (en) R. M. Alexander, « All-time giants: the largest animals and their problems », Palaeontology, vol. 41, no 6, , p. 1231–1245 (lire en ligne [archive du ], consulté le )
- (en) M. Clauss, R. Frey, B. Kiefer, M. Lechner-Doll, W. Loehlein, C. Polster, G. E. Rössner et W. J. Streich, « The maximum attainable body size of herbivorous mammals: Morphophysiological constraints on foregut, and adaptations of hindgut fermenters » (Submitted manuscript), Oecologia, vol. 136, no 1, , p. 14–27 (PMID 12712314, DOI 10.1007/s00442-003-1254-z, Bibcode 2003Oecol.136...14C, S2CID 206989975, lire en ligne [archive du ], consulté le )
- (en) C. Martin, I. Bentaleb et P. -O. Antoine, « Pakistan mammal tooth stable isotopes show paleoclimatic and paleoenvironmental changes since the early Oligocene », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 311, nos 1–2, , p. 19–29 (DOI 10.1016/j.palaeo.2011.07.010, Bibcode 2011PPP...311...19M)
- (en) Y. Wang et T. Deng, « A 25 m.y. Isotopic record of paleodiet and environmental change from fossil mammals and paleosols from the NE margin of the Tibetan Plateau », Earth and Planetary Science Letters, vol. 236, nos 1–2, , p. 322–338 (DOI 10.1016/j.epsl.2005.05.006, Bibcode 2005E&PSL.236..322W)
- (en) S. Sen, P. O. Antoine, B. Varol, T. Ayyildiz et K. Sözeri, « Giant rhinoceros Paraceratherium and other vertebrates from Oligocene and middle Miocene deposits of the Kağızman-Tuzluca Basin, Eastern Turkey », Naturwissenschaften, vol. 98, no 5, , p. 407–423 (PMID 21465174, DOI 10.1007/s00114-011-0786-z, Bibcode 2011NW.....98..407S, S2CID 19968371)
- (en) J. Meng et M. C. McKenna, « Faunal turnovers of Palaeogene mammals from the Mongolian Plateau », Nature, vol. 394, no 6691, , p. 364–367 (DOI 10.1038/28603, Bibcode 1998Natur.394..364M, S2CID 204998953)
- (en) E. B. Leopold, G. Liu et S. Clay-Poole, « Eocene-Oligocene Climatic and Biotic Evolution », Princeton University Press, Princeton, , p. 399–420 (ISBN 978-0-691-02542-1)
- (en) P. V. Putshkov, « "Proboscidean agent" of some Tertiary megafaunal extinctions », Terra Degli Elefanti Congresso Internazionale: The World of Elephants, , p. 133–136
- (en) « Paraceratherium », sur ARK: Survival Evolved Wiki
- (en) Tim Haines, The Complete Guide to Prehistoric Life, Canada, Firefly Books, (ISBN 1-55407-125-9, lire en ligne
 ), 176
), 176 - « Le mystère des géants disparus », sur France.tv
- « Le mamifère le plus grand du monde », sur Youtube
- (en) Angela Watercutter, « 35 Years After Star Wars, Effects Whiz Phil Tippett Is Slowly Crafting a Mad God », Wired, (lire en ligne, consulté le )
- (en) Terryl Whitlatch et Bob Carrau (ill. Terryl Whitlatch), The Wildlife of Star Wars : A Field Guide, Chronicle Books, (1re éd. novembre 2001), 176 p.
Voir aussi
Bibliographie
![]() : document utilisé comme source pour la rédaction de cet article.
: document utilisé comme source pour la rédaction de cet article.
- (en) Donald Prothero, Rhinoceros Giants: The Palaeobiology of Indricotheres, Indiana, Indiana University Press, (ISBN 978-0-253-00819-0).

- Jean-Loup Welcomme et Éric Poindron, Sur les traces du géant, Flammarion, (ISBN 2-08-068355-1)
Vidéos
- (en) [vidéo] The Rise and Fall of the Tallest Mammal to Walk the Earth sur YouTube.
- (en) [vidéo] Paraceratherium - Ancient Animal sur YouTube.
Articles connexes
Liens externes
- Ressources relatives au vivant :
 Portail de la paléontologie
Portail de la paléontologie  Portail des mammifères
Portail des mammifères  Portail du Cénozoïque
Portail du Cénozoïque  Portail des records
Portail des records