Syndrome d'effondrement des colonies d'abeilles
Le syndrome d'effondrement des colonies d'abeilles (en anglais, « Colony Collapse Disorder » : CCD) est un phénomène de mortalité anormale et récurrente des colonies d'abeilles domestiques notamment en France et dans le reste de l'Europe, depuis 1998[1],[2], aux États-Unis, à partir de l'hiver 2006-2007[3]. D'autres épisodes de mortalité ont été signalés en Asie et en Égypte[4],[5] sans être pour le moment formellement associés au CCD.
Pour les articles homonymes, voir CCD.
Ce phénomène affecte par contrecoup la production apicole dans une grande partie du monde où cette espèce a été introduite. Aux États-Unis, il fut d'abord appelé « syndrome de disparition des abeilles » ou bien « Fall-Dwindle Disease » (maladie du déclin automnal des abeilles)[6], avant d'être renommé CCD.
Le phénomène prend la forme de ruches subitement vidées de presque toutes leurs abeilles, généralement à la sortie de l'hiver, plus rarement en pleine saison de butinage (en). Aux États-Unis, près de 25 % du cheptel a disparu au cours de l'hiver 2006-2007. De nombreux pays européens sont touchés depuis le début des années 2000. Les pertes peuvent atteindre, localement, jusqu'à 90 % des colonies[7]. Les taux de mortalité hivernale des ruches d'abeilles domestiques, mesurés depuis l'apparition du phénomène sont quasi systématiquement supérieurs au taux d'environ 10 % observé auparavant[réf. nécessaire].
Ce syndrome est jugé très préoccupant par les apiculteurs, mais aussi par de nombreux écologues, économistes et experts en raison de l'importance économique et écologique de l'abeille en tant que pollinisatrice : les pommiers, mais aussi les amandiers, les avocatiers, les cerisiers, les oignons, les concombres, le coton, l'arachide, le melon, etc. dépendent à 90 %, voire à 100 % des abeilles pour leur pollinisation. Selon l'INRA, la survie de 80 % des plantes à fleurs et la production de 35 % de la nourriture des hommes dépendent directement des pollinisateurs[8]. Les services rendus à la pollinisation par les abeilles sont estimés à environ quinze milliards de dollars par an aux États-Unis[9]. Leur importance doit cependant être relativisée, compte tenu du rôle non négligeable tenu par les pollinisateurs sauvages[10]. L’abeille domestique reste toutefois indispensable pour des besoins massifs et ponctuels de pollinisateurs dans des zones pauvres en milieu sauvage. Le cas le plus emblématique est la production d'amandes en Californie : les 3/4 du cheptel d'abeilles américaines y sont rassemblés par une transhumance renouvelée chaque année[11], entraînant une augmentation des risques sanitaires due à la promiscuité.
Ce phénomène épidémique fait l'objet de polémiques scientifiques et médiatiques. Les chercheurs étudiant ce phénomène ne pouvaient s'appuyer que sur un corpus très pauvre d'études et de données, notamment écoépidémiologiques et génétiques. Depuis le milieu des années 2000 de nombreux travaux ont été publiés, ce qui a permis la constitution d'une bibliographie de plus en plus solide. Des avancées considérables ont été réalisées, notamment l'identification correcte des bio-agresseurs impliqués dans le phénomène et la mise en évidence d'effets synergiques divers. La thèse d'un phénomène multifactoriel[12] est maintenant l'objet d'un consensus. Le rôle central de l'acarien Varroa destructor est cité dans un rapport du Département de l'Agriculture des États-Unis en 2012 et dans la première partie de l'étude européenne « Prevention of honey bee COlony LOSSesa » en 2018.
En Europe, des apiculteurs ont désigné les pesticides systémiques comme les principaux responsables du phénomène dès 1995[13]. À partir de 1999, l'interdiction en France et en Europe de certains pesticides incriminés n'a pas eu d'effet mesurable sur le terrain, les mortalités demeurant inchangées. De nombreuses ONG écologistes et des journalistes soutiennent la thèse selon laquelle les pesticides sont la cause primaire du phénomène[14],[15]. L'Union nationale de l'apiculture française (UNAF) reconnaît le rôle de pathologies[16], mais en considérant toujours les pesticides comme la cause primaire. Des travaux en conditions artificielles ou simulées accusent les insecticides, sans forcément en faire une cause unique du phénomène. Ces travaux sont remis en cause par d'autres études qui considèrent que le manque de connaissances sur les besoins physiologiques des abeilles rend délicat la conception des protocoles (notamment en milieu fermé et semi-ouvert) et l'interprétation des résultats : les doses et les situations testées sont considérées comme extrêmes voire totalement irréalistes. Les études scientifiques ont cependant conduit l'autorité européenne de sécurité des aliments à constater que les tests réglementaires homologuant les molécules mises en cause ne permettaient pas d'en évaluer les risques et que certains produits phytosanitaires encore utilisés « présentaient un risque pour les abeilles »[17].
Définition
Le CCD désigne un syndrome caractérisé par la présence de trois symptômes affectant une colonie d'abeilles domestiques[18],[19] :
- l'absence d'abeilles mortes dans la ruche ou à proximité ;
- un déclin subit de la population d'abeilles adultes, laissant le couvain à l'abandon ;
- Une absence de pillage ou de cleptoparasitisme de la colonie dans l'immédiat.
D'autres formes de diminution de la population sont quelquefois confondues avec le CCD :
- Le dépeuplement rapide de ruche en pleine saison de production, à l'origine du début de la médiatisation de l'affaire en 1998[20],[21].[citation nécessaire]
- Les surmortalités hivernales observées surtout en Amérique du Nord depuis le milieu des années 1990 : les ruches sont mortes à la sortie de l'hiver. Des mortalités avec le même symptôme ont déjà été observées par le passé au Colorado en 1891 et 1896 mais le syndrome possède des caractéristiques nouvelles jugées alarmantes[9] :
- les pertes sont brutales et sans signe avant-coureur apparent ;
- l'effondrement peut avoir lieu toute l'année ;
Le phénomène se répand rapidement et touche aujourd'hui la quasi-totalité des zones d'élevage d'abeilles européennes. La reine abandonnée semble en bonne santé et souvent continue à pondre, alors qu'il n'y a plus assez d'ouvrières pour s'occuper du couvain. Les ouvrières restantes ne se nourrissent plus et le butinage s'arrête[9].
Le symptôme de la ruche vide en pleine saison de butinage reste rare et l’essentiel du phénomène prend la forme de mortalités hivernales massives et récurrentes.
Historique
En France, des pertes d'abeilles ou de colonies ainsi que des récoltes de miel anormalement basses sont signalées ponctuellement depuis la fin du XIXe siècle, généralement expliquées par des conditions météorologiques défavorables ou des maladies. Les premiers signalements d'une nouvelle vague de mortalité inhabituellement élevée ont lieu en 1998. Le phénomène prend à contre-courant le développement de l'élevage d'abeilles domestiques en Europe qui démarre dans les années 1960. La production ayant peu évolué depuis l'invention de la ruche moderne à cadre amovible en 1849, elle est peu à peu révolutionnée par l'arrivée d'apiculteurs professionnels ainsi que le développement des cultures industrielles mellifères comme le colza et surtout le tournesol. La mise au point de la production de reines contrôlée à l'aide de gelée royale permet de multiplier rapidement le nombre de colonies et ouvre la voie à la sélection génétique grâce à l'insémination artificielle des reines. La productivité explose : les ruches bien menées passent de 5 à 10 kg puis à 50 kg de miel par an[22], la production française de 8 000 t en 1961 progresse à près de 20 000 tonnes dans les années 1990 d'après la FAO[23]. De nombreux apiculteurs se professionnalisent dans les années 1980.
La crise vient briser un secteur en pleine croissance en Europe : alors que le nombre de colonies domestiques a progressé de 16,2 % (± 9,2) entre 1965 et 1985 il diminue ensuite de 16,1 % (± 5) entre 1985 et 2005, s'accompagnant d'une réduction de la population d'apiculteurs de 31,4 % (± 4,5), la production de miel français retombe sous les 15 000 t en 2010[24]. La plupart des pays d'Europe sont également affectés à la même période, avec des pertes allant pour certains apiculteurs jusqu'à 90 % à 100 % du cheptel au printemps. L’interdiction d’insecticides mis en cause comme le Gaucho ou le Régent TS n’a pas eu d’impact positif sur la mortalité des colonies et les pouvoirs publics ont commencé à mettre en place des programmes de recherche sur les troubles de l’abeille[25]. En Alsace des chiffres fiables montrent une nouvelle augmentation de la mortalité en 2013 qui atteint 24 %[26].
Aux États-Unis, des disparitions importantes d'abeilles ont été localement décrites dès 1896 et ont reçu plusieurs appellations : autumn collapse, May disease, spring dwindle, disappearing disease, fall dwindle disease[27]. Plus récemment, des disparitions ont été imputées à la loque américaine dans les années 1940 puis aux acariens parasites Varroa destructor et Acarapis woodi dans les années 1980[9]. L'arrivée de ces acariens aux États-Unis a fait passer la proportion de colonies ne survivant pas à l'hivernage de 5-10 % à 15-25 %[28].
À l'inverse de l'Europe la crise apicole américaine s'inscrit dans un contexte de recul de l'apiculture et de mutation : la production de miel a commencé à décroître dès l'après-guerre, passant de 120 000 t dans les années 1960 à 67 000 t en 2011[29]. En revanche la location de ruche pour la pollinisation[30] est devenu l'activité dominante des apiculteurs, entrainant une intensification de la production d’essaim, des déplacements longues distances et une remise en cause des pratiques traditionnelles comme la reconstitution des réserves des ruches dans les pâtures du Montana (elles-mêmes en régression au profit des cultures comme le maïs) en fin de saison[31].
Durant l'hiver 2006-2007 la crise éclate aux États-Unis, les pertes sont jugées catastrophiques par les spécialistes, certains apiculteurs ont perdu 70 % de leurs abeilles[32]. Plusieurs scientifiques ont considéré cette situation comme menaçant la pollinisation de plusieurs cultures maraîchères et fruitières aux États-Unis notamment[33],[34], cependant les effets sur le terrain ne semblent pas significatifs, les cultures américaines ne souffrent pas d'un quelconque manque de pollinisateur[35].
L'Organisation des Nations unies pour l'alimentation et l'agriculture estime que le nombre de ruches a globalement progressé d'environ 45 % sur l'ensemble du globe pendant la seconde moitié du XXe siècle[36] : la production de miel a fortement augmenté au Brésil (7 000 t à 41 000 t), en Argentine (20 000 t à 59 000 t), au Mexique (24 000 t à 57 000 t) et en Chine (53 000 t à 450 000 t) entre 1961 et 2011.
Chiffres
En Europe, les données disponibles avant 2008 sont peu comparables, en l’absence d’une méthode de mesure commune et d’un réseau de surveillance global. À partir de la création du réseau COLOSS en 2008, une coordination internationale permet d’affiner les mesures et d’harmoniser les protocoles[37].
En 2006 et 2007, neuf états européens sur treize ont déclaré un taux de mortalité supérieur au seuil normal de 10 %[25]. Entre 2009 et 2010, ce taux de disparition a augmenté mais il varie grandement d'un pays à l’autre, allant de 7 % à 30 %. Pour six des treize provinces et territoires du Canada, ce taux allait de 16 % à 25 % pour l’hiver 2009–2010 alors que pour cinq des vingt-deux provinces de Chine, il n’était que de 4 %[38].
En France, le taux de perte dues au syndrome d'effondrement pour l’hiver 2010–2011 chez les apiculteurs professionnels est estimé entre 0,75 % et 1 % (entre 17 % et 22 % de pertes annuelles, mais seulement 4,5 % des colonies perdues présentant les symptômes du syndrome d'effondrement[39]).
Aux États-Unis, une étude annuelle sur les pertes hivernales de colonies est conduite par le Service de recherche agricole et l’association Apiary Inspectors of America depuis 2007.
Pour l’hiver 2006–2007, pour les 13 États ayant répondu à l'enquête, 23,8 % des apiculteurs ayant rapporté des pertes de colonies avec absence d'abeilles mortes présentaient les caractéristiques du syndrome d'effondrement déterminés par les enquêteurs (50 % ou plus des colonies détruites désertées par les abeilles et absence de cadavres d'abeilles dans la ruche ou à proximité), avec un taux de pertes de 45 % contre 25,4 % pour les apiculteurs considérés comme ne relevant pas du syndrome ; au total, 31,8 % des colonies ont été perdues. 51,9 % des apiculteurs ont rapporté des pertes anormalement élevées, correspondant à 55,4 % de leur cheptel, alors que les apiculteurs ayant rapporté des pertes habituelles n’ont perdu que 15,9 % de leur cheptel[40].
Pour l'hiver 2007–2008, l'enquête a porté sur 19,4 % de l'ensemble des colonies présentes aux États-Unis. 35,8 % des colonies ont été perdues. 37,9 % des apiculteurs ont perdu au moins certaines de leurs colonies avec des symptômes du syndrome. Leur taux de perte hivernale était également plus élevé (40,8 % en moyenne) que ceux n'ayant pas rencontré ces symptômes (17,1 %). Les apiculteurs ayant estimé que leurs pertes étaient normales ont rencontré un taux de perte moyen de 21,7 %[28].
Pour l'hiver 2008-2009, l'enquête a porté sur 20,1 % des colonies. Le taux de perte est descendu à 28,6 %, avec un taux considéré comme normal à 17,6 %. Seulement 26,2 % des apiculteurs ont rencontré les symptômes du syndrome d'effondrement, qui concernaient 60,3 % des cas de leurs colonies perdues. Ces apiculteurs n'ont toutefois pas eu un taux de perte significativement plus élevé que ceux ne rencontrant pas le syndrome[41].
Pour l'hiver 2009-2010, sur la base de 17,7 % des colonies déclarées pendant l'été 2009, le taux de perte est remonté à 34,4 %, pour un taux acceptable à 14,5 %. 28,9 % des apiculteurs ont rencontré les symptômes du syndrome (sur la base de 65 % des répondants seulement), avec un taux de perte moyen de 62,2 %. Pour ces apiculteurs, 44,1 % des colonies ont été perdues, contre 26,7 % pour ceux n'ayant pas été confrontés au syndrome[42].
Pour l'hiver 2010-2011, sur la base de 11,5 % des colonies déclarées pendant l'été 2010, le taux de perte a été de 29,9 %, pour un taux acceptable évalué à 13,2 %. La part de colonies présentant les symptômes du syndrome d'effondrement est estimée à 26,3 %. Ces apiculteurs ont perdu en moyenne 62,3 % de leurs colonies, contre 56,5 % pour ceux n'étant pas affectés par le syndrome[43].
Les causes de mortalité supposées par les apiculteurs répondant à l'enquête sont une défaillance de la reine, la famine et la présence du varroa. Le nombre total de colonies exploitées aux États-Unis est en augmentation, de 2,34 millions en 2008 à 2,68 millions en 2010. L'explication avancée est que les apiculteurs multiplient leurs colonies pour compenser les taux élevés de pertes hivernales[43].
Une étude de scientifiques de l'Agence nationale de sécurité sanitaire de l'alimentation, de l'environnement et du travail fait le point sur la démographie des abeilles en Europe[44] :
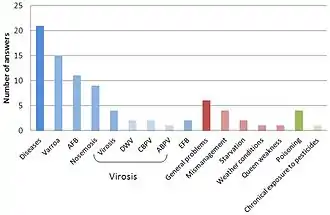
D'après ces chercheurs, la plupart des laboratoires évoquent dans l'ordre :
- les maladies parasitaires ou virales (69 % des cas),
- des problèmes de management d'élevage (14 % de cas)
- Les pesticides (5 %)
pour expliquer les mortalités. Les causes évoqués par les apiculteurs sont similaires.
Hypothèses explicatives



L’année 2007 voit la publication de nombreux travaux de recherche visant à identifier les causes de la disparition des colonies. Mi-2008, l’Union européenne, via le système COST, organise un réseau de recherche international pour la prévention de la perte des colonies : COLOSS[37].
En 2009, 25 % des apiculteurs japonais déclarent être confrontés au phénomène[4].
Les causes suggérées comprennent les pesticides[45], les infections par divers agents pathogènes, en particulier ceux transmis par les acariens Varroa et Acarapis, la malnutrition, les facteurs génétiques, les immunodéficiences, la perte d'habitat, la modification des pratiques apicoles ou une combinaison de facteurs[46],[47]. De nombreuses spéculations ont entouré les contributions des pesticides de la famille des néonicotinoïdes à la disparition des colonies d'abeilles, mais de nombreux ruchers en effondrement ne montrent aucune trace de néonicotinoïdes[47].
Des explications différentes selon les pays
Les causes de la mortalité des abeilles semblent différentes et sont d'ampleur différente selon les pays étudiés[48] :
- en Belgique, on estime que le varroa (acarien parasite) affaiblit les abeilles, les rendant vulnérables aux virus et bactéries car le varroa est lui-même vecteur de virus ;
- en Espagne le nosema ceranae (champignon microscopique) est considéré comme la principale cause de la mortalité des abeilles ;
- aux États-Unis, on a trouvé une corrélation entre l'effondrement des abeilles, apparemment désorientées et ne retournant pas à leur ruche, avec une souche de virus IAPV (Dicistroviridae) ;
- en France, on suspecte aussi des pesticides ou des synergies entre pesticides, dont le Gaucho, qui par précaution a été interdit sur le tournesol depuis 1999 ;
- à Hawaï le rôle de Varroa destructor et du virus des ailes déformées (DWV) est confirmé par l'effondrement des colonies à la suite de l'arrivée du parasite sur l'archipel ;
- en Suisse, selon le site internet Swissinfo, scientifiques et apiculteurs s'accordent pour désigner le Varroa comme la cause principale de la crise apicole[49]. La Tribune de Genève souligne que si certains produits phytosanitaires sont aussi incriminés, soupçonnés qu'ils sont « d'affaiblir, même à des doses infimes, les capacités de résistance et d'orientation des abeilles[50] », les interdire ne résoudrait pas la crise si cela revenait à augmenter l'usage de pratiques ou de produits encore plus dangereux.
« L’influence d’insecticides sur la mort des abeilles doit être relativisée, aucun lien direct entre l’utilisation de pesticides et la mort des abeilles n’a pu être prouvé »
— Nadja Pieren (UDC/BE)
« Le lobby pharmaceutique est très fort [...] le rôle des insecticides ne fait aucun doute »
— Thomas Minder (politicien suisse sans étiquette mais proche des écologistes[51], entrepreneur dans le dentifrice et les produits d'hygiène buccale)
.
- Le Conseil fédéral a suspendu les néonicotinoïdes clothianidine, imidaclopride et thiaméthoxame présents dans des pesticides commercialisés par le groupe suisse Syngenta et l'Allemand Bayer pour deux ans en attendant des études complémentaires. Deux motions ont été adoptées, visant à mettre en place un plan d'action national pour étudier scientifiquement les causes de la mortalité des abeilles, et une seconde visant à réduire les risques et l'usage des produits phytosanitaires d'ici 2023. L’Union suisse des paysans, Apisuisse et le Centre de recherches apicoles d’Agroscope souhaitent un plan national de mesures afin de mieux cerner les causes de la mortalité des abeilles. Ils prônent une approche globale plutôt que des interdictions « précipitées » de produits phytosanitaires[52] ;
- en Égypte, une étude a pu mettre en évidence que les ruches bénéficiant d'une flore variée et d'un environnement sans pesticides n'était pas touchées par le syndrome contrairement aux autres[4].
Rapport des autorités américaines (2012)
À l'automne 2012, le Département de l'Agriculture des États-Unis et l’Environmental Protection Agency ont publié un rapport[53] permettant d'établir des conclusions solides :
- un consensus émerge sur le fait qu'un ensemble complexe de stress et de pathogènes est associé au CCD ;
- le parasite acarien Varroa destructor reste le parasite le plus préjudiciable et le plus étroitement lié aux déclins des colonies ;
- de multiples virus sont associés au CCD ;
- le Varroa est connu pour amplifier la charge virale dans les abeilles ;
- la loque européenne est plus souvent détectée aux États-Unis et pourrait être liée à des pertes de colonies ;
- la nutrition a un impact majeur dans la survie individuelle des abeilles et dans la longévité des colonies ;
- la flore intestinale joue un rôle clé dans la nutrition, la détoxification des produits chimiques et la protection contre les maladies ;
- l'effet immédiat et sublétal de pesticides sur les abeilles est de plus en plus documenté et est une préoccupation importante.
Néanmoins des travaux en condition de champ sont nécessaires pour établir le niveau réel d'exposition aux pesticides et les risques associés en termes de déclin général des populations d'abeilles aux États-Unis ; - la cryoconservation de la semence des abeilles permet déjà de préserver le germoplasme des meilleures abeilles domestiques pour des croisements. L'amélioration de la variabilité génétique doit aboutir à l'amélioration de la résistance et de la productivité des nouvelles colonies ;
- les retombées du séquençage du génome de l'abeille sont maintenant utilisées pour comprendre et aborder la question de la reproduction, de l’interaction avec les parasites et les méthodes d'élevage permettant de réduire le stress des abeilles et les rendre plus productives.
L'étude européenne Prevention of honey bee COlony LOSSes (2013)
Des d'études de quatre ans sur la santé des cheptels apicoles européens ont conclu :
- le Varroa destructor en combinaison avec des virus est la principale menace pour la survie des colonies,
- des interactions entre parasites, pathogènes et pesticides peuvent affecter la santé individuelle des abeilles,
- Nosema ceranae peut affecter les colonies mais n'est pas un facteur majeur[54].
ANSES 2015
L’Anses s’est autosaisie en 2012[55] sur la question des co-expositions des abeilles à différents facteurs de stress et leur rôle dans les phénomènes d’affaiblissement, d’effondrement ou de mortalité des colonies d’abeilles, avec une attention toute particulière sur les interactions entre ces facteurs.
« Les résultats de l’expertise mettent en évidence un nombre important d’agents infectieux et parasitaires affectant les colonies d’abeilles, de nombreux résidus de xénobiotiques (insecticides, fongicides et acaricides) présents dans les matrices apicoles, ainsi qu’une grande diversité de facteurs de stress auxquels les abeilles peuvent être exposées, de manière concomitante ou successive. L’expertise souligne le caractère multifactoriel des causes de mortalité des colonies d’abeilles et met en évidence le rôle des co-expositions aux pesticides et aux agents infectieux dans le déterminisme de leur effondrement.
Dans ce contexte, l’Agence recommande d’intervenir sur l’ensemble de ces facteurs, notamment au travers de l’appropriation et du respect des bonnes pratiques apicoles, mais aussi de la diminution globale de l’exposition des abeilles aux pesticides. L’Agence souligne également l’importance de disposer à terme d’un réseau d’observation harmonisé et structuré au niveau national, permettant de produire des bilans de l’état de santé des colonies, notamment par la création de ruchers de référence. Enfin, l’Agence recommande- dans le cadre de discussions à engager au niveau européen – l’élaboration puis l’intégration de tests supplémentaires pour mesurer l’effet de co-expositions à des fongicides et des acaricides dans la procédure d’évaluation de la toxicité des insecticides. »
— Anses, Santé des abeilles : impact de la co-exposition des colonies aux pesticides et aux agents infectieux
Recensement et hiérarchisation des causes possibles
Le rapport de l'AFSSA intitulé Mortalités, effondrements et affaiblissements des colonies d'abeilles et présenté en colloque le 17 juin 2009[25] énumère quarante facteurs contribuant potentiellement à la surmortalité des abeilles.
Causes primaires
De nombreux travaux scientifiques permettent d'identifier des facteurs qui ont un effet certain sur les abeilles et qui pourraient expliquer seuls ou en association la crise apicole.
Les pathologies anciennes et émergentes
Les ruches victimes du syndrome semblent mieux se rétablir après une désinfection de la ruche par irradiation[56],[57] ce qui laisse à penser que l'origine de la crise apicole est un agent biologique et qu'il se transmet de façon verticale et horizontale. Les abeilles possèdent peu de gènes de résistance aux pathogènes, seulement 1/3 des familles de gènes immunitaires qui équipent des insectes communs comme la mouche drosophile et l'anophèle sont présents chez l'abeille[58]. Cette faible robustesse génétique rend les abeilles plus sensibles à l'avènement de nouveaux pathogènes que la plupart des insectes. 29 agents pathogènes de l'abeille potentiellement cause de mortalité ont été recensés par l'ANSES : 4 prédateurs (dont le frelon asiatique), 5 parasites (comme le Varroa destructor), 3 champignons (dont celui responsable de la nosémose), 5 bactéries (dont celle responsable de la loque américaine) et 12 virus (comme celui responsable de la maladie de la paralysie chronique)[59].
Maladies parasitaires
Les varroas, et plus particulièrement l’apparition et la diffusion d'une nouvelle espèce, le Varroa destructor est le principal facteur mis en cause dans de nombreux rapports et travaux scientifiques. Selon le centre de recherche sur l'abeille suisse de l'agroscope Liebefeld-Posieux, le Varroa destructor peut être désigné comme un marqueur prédictif du CCD[60]. Selon un rapport de stage de l'université de Montpellier, à la Réunion, des mesures doivent être mise en place pour éviter l'introduction du varroa sur l'île car « le risque acceptable est quasiment nul car les conséquences de l'arrivée du parasite sur l'île seraient catastrophiques »[61]. De même sur l'île d'Ouessant, selon le Conservatoire de l'abeille noire bretonne, l'absence de varroa, d'agriculture intensive et de pesticides explique des taux de mortalité exceptionnellement bas (moins de 3 %)[62],[63]. En novembre 2014, le Conservatoire d'Ouessant a fait valider l'absence de varroa par le laboratoire Labéo dans l'Orne (61), agréé au niveau européen, sur un protocole défini par l'ANSES et appliqué par des Agents Sanitaires du Finistère. De nombreux travaux scientifiques le désignent comme la clé de cette crise sanitaire[64],[65],[66],[67],[68] et reconnaissent son rôle central dans les mortalités[69]. En Ontario il est responsable de 85 % de la mortalité hivernale des colonies[70], les auteurs de l'étude le considèrent même comme la cause principale de l'effondrement des cheptels en climat froid.
Aux États-Unis, 39 causes ont été analysées lors d'un atelier financé par Bayer un des plus importants producteurs de pesticides au monde en septembre 2012, les résultats de cette rencontre ont été publiés en février 2014[71] : si les participants n'ont pu analyser en profondeur que 3 facteurs, et ont restreint leurs interventions à la seule surmortalité hivernale des abeilles, en excluant de traiter le cas du syndrome d’effondrement des colonies ; ils concluent que Varroa destructor, notamment en combinaison avec différent virus, est une cause « probable » du déclin des populations d'abeilles, les problèmes d'alimentations une cause « possible » et les insecticides néonicotinoïdes une cause « improbable » mais un contributeur possible.
L'identification de ce ravageur fut longue : en 1904 l'espèce Varroa jacobsoni est identifiée chez Apis ceranae[72]. Ce dernier fut ponctuellement mis en contact avec l'abeille européenne en 1950 (Corée), 1958 (Japon) puis de façon plus significative dans les années 1960[73] quand des reines d'origine italienne sélectionnées pour leur production massive de gelée royale furent envoyées en Asie. Ce croisement des populations entraina la création d'une nouvelle espèce de Varroa, le Varroa destructor, issu du Varroa jacobosoni avec lequel il fut confondu, jusqu'en 2000 où il fut enfin décrit par les chercheurs australiens Anderson et Trueman[74], la génétique confirmant ensuite la séparation des deux espèces[75]. Ce problème d'identification a ralenti la prise de conscience du problème : l'infection étant bénigne chez Apis ceranae, le rôle de ce nouveau ravageur ne fut pris au sérieux que dans les années 2000. D'autres génotypes pourraient émerger en cas de nouveau contact entre les populations européennes et asiatiques[76] et font peser une menace supplémentaire sur le cheptel d'abeilles domestiques.
Varroa destructor est capable de modifier la composition chimique de sa cuticule pour passer d'une espèce à l'autre[77]. La présence de marqueurs chimiques adaptés et adaptables expliquent "furtivité" des parasites, les abeilles ne peuvent éliminer les larves contaminés sans détecter l'odeur du Varroa[77]. D'après les auteurs de l'étude la coévolution entre Apis Ceranae et Varroa a donné au parasite une excellente capacité d'adaptation de ces marqueurs chimiques. L'abeille européenne n'a pas coévolué et n'a pas une aussi bonne capacité de détection et d'action que l'abeille asiatique, d'où sa grande sensibilité face à ce parasite très bien adapté.
Profitant d'échange de reines et de colonies entre l'Europe et l'Asie Varroa destructor atteint l'Europe dans les années 1970 et la France en 1982 via l'Italie. Il est signalé aux États-Unis à la fin des années 1980[78], où il précède de peu une chute spectaculaire du rendement des ruches[2]. Ce décalage est conforme au délai observé entre le début de la crise apicole aux États-Unis et en Europe. Sa présence dans les ruches domestiques est devenue quasi systématique[79]. Seules quelques îles isolées échappent ainsi au CCD : l'Australie[80], la Réunion, Ouessant[81]. À la Réunion les scientifiques du CIRAD considèrent que son introduction provoquerait la perte de la quasi-totalité de la production de miel[82]. L'introduction de Varroa en Nouvelle-Zélande[83] au début des années 2000 s'est traduite par des mortalités anormales néanmoins limitées par l'usage de produits acaricides, mais cette solution reste provisoire[84]. En 2010 c'est au tour de Madagascar d'être contaminée, des mortalités massives sont rapidement observées[85] : 60 % des colonies infectées ont disparu. Cet effondrement généralisé et rapide de cheptels apicoles est identique à ce que l'Europe et l'Amérique du Nord ont subi quelques années plus tôt.
Le Varroa destructor a de multiples effets sur les colonies infectées : il les affaiblit[86], perturbe la capacité des abeilles à maintenir leur hydratation[87], altère leur système immunitaire[88],[89],[90], et véhicule des virus. Les abeilles qui naissent dans des colonies fortement parasitées sont faibles, parfois mal formées et meurent rapidement[73]. Le parasite se multiplie relativement lentement, d'un facteur 7 à 10 par an[91]. Si la colonie est contaminée par une fondatrice, il faudra plusieurs années avant que la population de Varroa ne mette en danger la colonie. Néanmoins la population peut exploser d'un facteur 1000 en cas de conditions météorologiques très favorables au parasite : le seuil de danger (30 % des adultes atteints) peut alors être dépassé très rapidement.
Traité dès le début des années 1980 avec succès[92] il a peu à peu acquis des résistances aux traitements dès le milieu des années 1990 en France[93] et en Italie[94], ce qui est contemporain du début de la crise apicole. La situation se dégrada rapidement avec la diffusion des résistances au Royaume-Uni à partir de 2000[95] et aux États-Unis à partir de 2001[96] : à la suite d'un signalement d'un apiculteur, des scientifiques du USDA-ARS Bee Research Laboratory ont réalisé des tests avec ces colonies, mettant en évidence la présence de Varroa résistants au coumaphos.

L'opération ne prend que 2 à 4 secondes.
D’autres parasites tels qu’Acarapis woodi et Paenibacillus larvae ont déjà causé des mortalités documentées par le passé[9]. La loque européenne est un autre parasite du couvain de l'abeille qui semble en extension, parallèlement au syndrome d'effondrement, mais sa faible virulence et la grande spécificité des symptômes ne permettent pas d'en faire le facteur unique déclencheur des mortalités. Il en va de même d’Apocephalus borealis, une petite mouche endoparasitoïde également mise en cause[97],[98].
Maladies virales
Le virus IAPV : une étude parue dans la revue Science en 2007 fait état de l'analyse des organismes commensaux des abeilles s'étalant sur une période de trois ans[18]. Ce rapport a déterminé que le virus israélien de la paralysie aiguë (Israeli acute paralysis virus of bees, ou IAPV), initialement décrit par un chercheur israélien, est fortement corrélé avec le syndrome d'effondrement des colonies. Selon l'un des coauteurs de l'étude, Ian Lipkin : « nos résultats indiquent que l'IAPV est un marqueur significatif du CCD. L'étape suivante est de déterminer si l'IAPV, seul ou de concert avec d'autres facteurs peut induire le syndrome chez des abeilles saines. » Pour le moment on ne sait pas si ce virus est un symptôme ou une cause de l'effondrement. Les effets de l'IAPV sont multiples :
- en 2013 une équipe de chercheurs chinois décrit des modifications de comportement des abeilles à la suite de l'infection par le virus IAPV : les abeilles contaminées réagissent à des concentrations de sucre plus faibles et voient leur taux de retour à la ruche (homing) fortement réduit. Les chercheurs concluent que le virus, très présent dans la tête de l'abeille, perturbe le fonctionnement cérébral permettant l'apprentissage, la navigation et l'orientation des abeilles[99] ;
- les pupes artificiellement contaminées ont leur métamorphose stoppée et subissent une mortalité importante dépendant de leur patrimoine génétique[100].
En 2009 une équipe israélienne a mis au point une méthode de lutte contre le virus en utilisant le RNA silencing (en)[101]. Ce traitement est développé par l'entreprise Beelogic, rachetée par Monsanto en 2011, et vise à le mettre sur le marché sous la dénomination "Remembee". Le Varroa destructor est un des vecteurs reconnus de l'IAPV[102]. D'autres virus (ex. : virus de la maladie noire, virus des ailes déformées) et bactéries sont clairement mis en cause, ce qui était suggéré par l'aspect épidémique et brutal des foyers de syndrome d’effondrement des colonies d'abeilles.
D'autres virus sont montrés du doigt[103],[104] : le virus des ailes déformées (DWV) est notamment un marqueur reconnu du CCD[105], peut provoquer une baisse drastique de la fécondité des reines[106]. Il peut lui aussi être traité à l'aide d'ARN silencing[107]. La progression mondiale du Varroa a sélectionné des variantes du DWV, ce qui a permis à ce virus de devenir un des plus largement distribués et des plus contagieux des virus des insectes sur la planète[108].
Nosémose
L'infection fongique par le Nosema ceranae, dont la présence en Europe est contemporaine du début de la crise apicole[109] fait l'objet d'une attention de plus en plus soutenue de la part de la communauté scientifique, certains lui attribuent même un rôle clé dans la crise apicole[110]. Son arrivée en Europe et aux États-Unis coïncide avec le début de la crise : 1998 en Europe[109] et 2006 aux États-Unis[111], mais les travaux scientifiques correspondant ne seront publiés qu'à partir de 2006-2007.
Ce champignon microscopique unicellulaire commensal de l'abeille asiatique Apis cerana est morphologiquement identique au Nosema apis autochtone ce qui a ralenti l'identification de cette maladie émergente : les scientifiques et vétérinaires[112] ont longtemps confondu ces deux pathogènes car les spores isolées pour diagnostiquer la maladie sont identiques. Il ne peut être identifié qu'en faisant appel à une analyse ADN. L'effet de ces deux espèces est très différent : Nosema apis craint la chaleur et provoque des diarrhées durant l'hiver que les apiculteurs reconnaissent facilement et savent traiter, Nosema ceranae colonise l'intestin plus en amont, affectionne les températures estivales et ne provoque pas de symptôme permettant un diagnostic différentiel[113]. Décrit pour la première fois en 1996, il est détecté dans l'abeille européenne en Europe en 2006[114] et ne sera reconnu comme pathogène à part entière de l'abeille européenne qu'en 2007 avec la publication de travaux de Wei-Fone Huang du Département d'entomologie de Taïwan[115]. D'autres travaux vont découvrir par la suite que ce pathogène est présent en Europe depuis au moins 1998 et aux États-Unis depuis 2006 : cela coïncide avec le début de la crise apicole, notamment des phénomènes de mortalité estivale inexplicables.
Depuis les données le mettant en cause s'accumulent, il est souvent retrouvé dans le corps des abeilles mortes et joue certainement un rôle important dans le phénomène de mortalité mondiale[116],[117]. Il est capable de provoquer une mortalité massive[118], sans abeille morte visible dans la ruche[3], une réduction de la production de miel et des autres produits de la ruche, ainsi que les fameux troubles du comportement des abeilles et des bourdons qui caractérisent le CCD[119] :
- Perturbation du butinage ;
- Réduction du soin aux larves : les jeunes ouvrières partent butiner plus jeunes ;
- Réduction du taux de retour à la ruche (homing) ;
- Problèmes d'orientation[120],[121].
- Réduction de l'espérance de vie[122]
Ces changements de comportement s'expliqueraient par une perturbation hormonale des jeunes ouvrières : une fois contaminées, leur taux d'hormone juvénile augmente provoquant l'acquisition précoce du comportement de butinage et une mortalité plus précoce (9 jours, ce qui est considérable pour un insecte qui vit 40 à 60 jours). Les jeunes ouvrières quittent alors la ruche, délaissant le soin du couvain et perturbant la capacité de la ruche à réguler ses activités[123]. Les scientifiques de l'INRA d'Avignon suggèrent qu'il s'agit de mécanisme de défense de la colonie : les abeilles infectées choisiraient volontairement d'accomplir des tâches plus risquées, les abeilles saines resteraient s'occuper des larves[6].
Une équipe de chercheurs espagnols sous la direction du docteur Higes a découvert plusieurs colonies d'abeilles atteintes du syndrome et qui ne présentaient comme unique anomalie que la présence de ce seul champignon microscopique. Après traitement des abeilles survivantes avec l'antifongique flumagillin, ils ont observé une guérison totale des symptômes[124]. Il a aussi noté que les ruches atteintes étaient rapidement décimées, en 9 jours seulement. Cette avancée permet de proposer aux apiculteurs un suivi des abeilles pour la présence de ce parasite ainsi qu'un traitement. Pour autant, des résistances sont à prévoir et pourraient remettre en cause cette stratégie, d'autant plus que l'antibiotique affecte la physiologie de l'abeille[125]. Dans une nouvelle review publiée en 2013 Higes et son équipe concluent que cette infection fongique joue un rôle global dans les mortalités des ruches et dans les pertes de productivité[117]. Parmi les traitements testés contre Nosema ceranae, peu d'alternatives aux antibiotiques sont efficaces[126], pour le moment seul des apports d'acides organiques produits par Lactobacillus johnsonii CRL1647 ont permis une amélioration de la vigueur de la ruche et des réserves de graisse des abeilles, seuls ou en association avec le flumagillin[127]. Les auteurs concluent que leurs études ouvrent la voie à de nouvelles substances de contrôle contre les nosémoses.
Pour autant Nosema apis ne peut être totalement innocenté, une étude américaine de 2012 montrant que si sa présence n'est pas un marqueur du CCD, les abeilles des ruches malades ont une charge en ADN de Nosema apis 20 fois supérieure à celles provenant de ruches saines[128]. De plus, les néonicotinoïdes et autres insecticides affaiblissent les abeilles, diminuent l'hygiène des ruches, permettent un développement anormal du Nosema et ainsi provoquer un effondrement des colonies[129]. En 2015 une nouvelle étude[130] apporte de nouveaux éléments décrivant les effets de l'infection (à faible dose selon les auteurs) par Nosema apis: en utilisant des puces RFID une équipe de chercheurs universitaires australiens a mis en évidence une réduction drastique de l'efficacité de la récolte de pollen (les abeilles malades privilégient les plantes riches en nectar au détriment de celle offrant du pollen) et de l'espérance de vie des abeilles contaminés, elles deviennent butineuse plus tard et meurent plus tôt. D'après les auteurs cette maladie fréquente pourrait affecte les capacités de pollinisation des abeilles.
Effets synergiques
Ces pathologies anciennes et nouvelles provoquent des effets de synergies mis en évidence par des travaux scientifiques.
Le Varroa destructor est à l'origine d'un grand nombre de synergies. Il est un important vecteur de virus[67]. Plus le parasite reste longtemps sur l'abeille, plus la charge virale de l'abeille parasité sera importante[131]. Plus une femelle varroa reste longtemps sur son hôte, plus ses filles (pondu sur le couvain) risque de provoquer des déformations très handicapantes sur la génération suivante d'ouvrières[132]. Une combinaison entre Varroa et une variante particulière du virus des ailes déformées (DWV) semble être la cause des CCD dans les îles Hawaï[133] : l'irruption de Varroa modifie l'équilibre entre les virus déjà présents chez l'abeille, une variante particulière du DWV devient dominante, la charge virale explose d'un facteur un million et provoque la mort de la colonie plusieurs années après la contamination. Plus de 65 % des colonies de l'île d'Oahu ont été détruites (en 2012) depuis l'introduction de Varroa destructor en 2007[134]. C'est la seule étude qui propose un véritable scénario pour les CCD car elle repose sur des observations de terrain, autant sur des colonies sauvages que d'élevage, avec des dimensions spatiales et temporelles qui font concorder la cause présumée et l'observation de mortalité. D'autres travaux confirment cette synergie négative entre Varroa et le virus DWV[135],[136] mais aussi avec le virus AKI (Acute-Kashmir-Israeli complex)[137]. Un effet synergique entre le Varroa et l'acarien de la trachée Acarapis woodi a aussi été observé[138]. En 2015, une étude publiée dans la revue Science met en évidence la responsabilité des pratiques apicoles: les souches de DWV qui déciment les colonies d'abeilles domestiques viennent toutes d'Europe et que leur diffusion a suivi celle de Varroa[139].
Nosema ceranae est aussi impliqué dans des phénomènes de mortalité synergique. Une équipe américaine a mis en évidence la présence combinée de Nosema ceranae et de virus IIV (invertebrate iridescent virus) chez des ruches touchés par le CCD durant le pic de 2006-2007, 2008 et 2009 en Floride[110]. Cette même étude montre que des ruches non atteintes par le CCD venant d'Australie et de ruchers sédentaires du Montana ne sont pas contaminées par cette paire de pathogènes. En 2010 des mortalités massives des colonies d'abeilles du Péloponnèse ont été attribuées à une probable synergie entre la mycose Nosema ceranae et des virus[140]. Par contre Nosema ceranae serait plutôt antagoniste du virus DWV[141]. L'analyse des corrélations entre la charge de différentes espèces de pathogènes dans une étude américaine[128] démontrerait que chez les ruches malades la charge de certains pathogènes serait covariante, ce qui expliquerait la rapidité du dépeuplement des colonies atteintes par le CCD[142].
En 2013, des chercheurs brésiliens ont mis en évidence une possible synergie entre l'infection par Nosema ceranae et l'utilisation de fongicides[143]. En septembre 2013 des scientifiques slovènes concluent à l'existence d'un effet synergique entre Nosema ceranae et le CBPV (virus de la paralysie chronique de l'abeille)[144].
En 2015 une étude de l'Université de Manitoba avance que la contamination par Nosema réduirait les capacités de l'abeille à se débarrasser des Varroa[145].
Pathogène ou combinaison encore inconnue
En 2013 des chercheurs belges[66] ont mis en évidence des liens synergiques entre Varroa destructor, Nosema ceranae et le trypanosome de l'abeille Crithidia mellificae qui peut désormais être considéré comme un des facteurs du CCD. Cette étude montre aussi que les abeilles belges sont contaminées par six pathogènes jusque-là non signalés en Europe : ALPV (Aphid Lethal Paralysis Virus) lignée Brookings, VdMLV (Varroa destructor Macula-like Virus), viruses of the LSV complex (Lake Sinai Virus), Spiroplasma melliferum (bactérie), Apicystis bombi (protozoaire) et Apocephalus borealis. Ces résultats montrent que la santé des abeilles est un sujet de recherche loin d'être épuisé, il est possible que tous les agents impliqués ne soient pas encore identifiés.
Les pratiques apicoles et agricoles intensives
Les pratiques intensives font l'objet de critiques : un article du East Bay Express[146] du 1er août 2007 s'interroge au sujet de l'intensification de l'apiculture industrielle pratiquée aux États-Unis. « Les abeilles sont plus libres de leurs mouvements que n'importe quel autre animal d'élevage », mais « une exploitation apicole commerciale ressemble plus à une cité HLM qu'à un pré campagnard » et pourraient expliquer les mortalités.
Transhumance et concentration des ruchers
La transhumance, pratique très ancienne[147] dans l'histoire de l'apiculture, s'est intensifiée avec la professionnalisation et la production de miel spécifique. Source de stress pour les abeilles, elle les rend plus vulnérables aux parasites et aux maladies, et pourrait diminuer leur capacité à fonctionner naturellement[146]. Ces échanges facilitent aussi la propagation des maladies entre les ruchers. Dans la nature, on trouve au maximum trois à quatre ruches sauvages par kilomètre carré, qui "connaissent" leur environnement et bénéficient d'une alimentation très variée (pollen, nectar et miellats de milliers de plantes différentes), pour une production intensive de miel ou la pollinisation sous contrat c'est une à huit ruches par hectare qui sont déposées[148] et ne peuvent se nourrir que d'une source alimentaire unique.
La dégradation de l'alimentation naturelle
L'alimentation est un facteur très important de la crise apicole, la malnutrition des abeilles est très courante. Elle accroît le stress généré par la mobilité qui leur est imposée par les apiculteurs les louant pour la pollinisation des cultures. La réduction de la biodiversité florale du fait de la réduction des zones de friches, prairies, jachères, l'appauvrissement des rotations culturales notamment en légumineuses, l'artificialisation des terres, et le fauchage systématique de la végétation herbacée ont fortement réduit les ressources alimentaires quantitativement mais aussi qualitativement. Une alimentation pauvre en pollen des larves affectera les capacités de butinage et de communication (danse) des abeilles adultes[149]. Toutefois l'infestation par varroa peut annuler les effets bénéfiques d'un environnement riche en ressources alimentaires[150].
La diversité qualitative de l'alimentation et une alimentation suffisante sont deux facteurs importants pour le système immunitaire et la santé. En Europe les changements dans la PAC (réduction des surfaces de luzerne, tournesol, fauche des jachères plus intensives) ont eu un impact fort sur les ressources alimentaires des abeilles à partir du début des années 1990[151]. Aux États-Unis c'est la mise en culture de vastes surfaces de prairies qui servaient autrefois à reposer les ruches après plusieurs utilisations comme pollinisateurs de culture qui est pointée du doigt[31]. Un cercle vicieux se forme : la diminution de la diversité et du nombre des plantes à fleurs affaiblit les pollinisateurs et diminue leur nombre, mal nourris les butineurs sont plus sensibles aux agressions biologiques, climatiques et anthropiques. Ce déclin parallèle à celui de la biodiversité florale est également mis en évidence pour les papillons[152] et plus généralement les pollinisateurs sauvages qui sont également en déclin.
Cette monotonie des sources alimentaires affaiblit le système immunitaire des abeilles : les mélanges de différents pollens sont plus nourrissants qu'une seule espèce[153]. Une étude de l'INRA-Avignon a confirmé ce phénomène : une moindre diversité en pollen de son alimentation affecte le système immunitaire des hyménoptères ; lors de nombreux tests (avec six préparations de pollen), les pollens polyfloraux (issus de différentes espèces végétales) ont toujours renforcé la santé des abeilles, « même quand le pollen monofloral était plus riche en protéines[154]. » Si les protéines sont une condition nécessaire à une bonne immunité chez les insectes[155], chez l'adulte comme pour la larve[156], d'autres composés pourraient intervenir. Les monocultures florales ont réduit la qualité du bol alimentaire des butineurs : en consommant du pollen d'un grand nombre d'espèces, les insectes ont le plus de chances de trouver au bon moment les oligoéléments (minéraux, acides aminés[157] acide folique, biotine et vitamines A, D, E et K) et les aliments (protides, glucides et lipides dont stérols) dont ils ont vitalement besoin[31].
Cette relation entre la santé, la productivité et la qualité de l'alimentation a notamment été observée en France par des expérimentations menées entre 2005 et 2006 dans le Loiret basées sur la comparaison de ruchers et montrant une augmentation de 7 % de la production de miel dans les zones où des jachères apicoles riches en pollens variés étaient implantées[158]. Des travaux scientifiques confirment l'effet positif des jachères fleuries sur les bourdons[159],[160],[161].Une expérimentation menée par les groupes Syngenta et Sainsbury’s au Royaume-Uni aurait montré que les populations d'insectes butineurs, notamment les bourdons et les papillons, pouvaient augmenter très rapidement (+600 % en 3 ans pour les bourdons, facteur 12 pour les papillons)[162] à l'aide d'aménagement de l'espace agricole et rural. Néanmoins ces travaux n'ont pas été publiés dans un journal scientifique à comité de lecture et doivent être interprétés avec une grande prudence. Plusieurs études universitaires semblent confirmer le lien fort entre gestion du paysage agricole et les populations de pollinisateurs : en testant des fauches plus tardives des prairies et le maintien de zones refuges non fauchées, une augmentation des populations d'insectes pollinisateurs est observée, progrès qui s'accumulent d'année en année confirmant a priori une vraie dynamique de population positive et pas seulement une concentration des pollinisateurs[163],[164].
L'alimentation artificielle : le « nourrissement »
Depuis les années 1970 l'utilisation de sirop de maïs à haute teneur en fructose est devenue très fréquente pour améliorer le rendement des ruches, la plupart des agriculteurs commerciaux distribuent ce substitut, seul ou additivé, à l'alimentation naturelle des abeilles. Le nourissement permet d'extraire plus de miel et de laisser plus de place pour la production de couvain dans la partie basse de la ruche. Couplé avec une sélection génétique qui favorise la prolificité des reines, la perte de diversité florale de l'espace agricole, le nourissement est au centre d'un cercle vicieux: pour profiter des cultures industrielles à fort rendement en nectar mais à floraison courte ou pour assurer des contrats de pollinisation, les apiculteurs pratiquent le nourissement spéculatif[165] à la sortie de l'hiver pour avoir des ruches pleines de butineuses. Lourdement chargées en abeilles mais pauvres en réserves puisque l'apiculteur prélève la hausse et que la partie basse est remplie de couvain, les ruches doivent ensuite recevoir des apports supplémentaires dès la fin de l'été pour passer l'hiver[166]. Les abeilles domestiques sont devenues très dépendantes de cette alimentation exogène. Cette alimentation synthétique est pauvre, elle affaiblit le système immunitaire des abeilles, notamment leur capacité à éliminer les toxines naturelles ou synthétiques (pesticides[167] et polluants)[168].
Un autre risque est la formation d'hydroxyméthylfurfural (HMF), une molécule toxique qui se forme dans le sirop de glucose-fructose en cas d'exposition à des températures entre 45 et 50 °C[169], ce risque est connu des apiculteurs et une norme existe pour le miel[170], les données permettant d'évaluer l'impact de cette molécule sur la santé des abeilles sont encore rare[171],[172], même si des travaux belges récents décrivent un cas de mortalité anormale pouvant être lié à une distribution de sirop de sucre de betterave très riche en HMF[173]. Le nourrissement est une fois de plus mis en cause par une étude du Département d'entomologie et de développement cellulaire de l'Université de l'Illinois[174]. Les chercheurs ont en effet mis en évidence le rôle très important de certains constituants du miel, notamment l'acide paracoumarique dans l'activation des gènes de détoxification. L'ajout de ce composé dans un sirop classique de nourrissement augmente de 60 % la vitesse d'élimination métabolique du coumaphos, un insecticide utilisé dans la lutte contre Varroa. Les auteurs concluent que l'usage de sirop de nourrissement affaiblit les défenses des abeilles et pourrait contribuer au phénomène d'effondrement des colonies exposées à des pathogènes et des pesticides. Une autre étude de l'Université de Graz exclu que le HFM puisse provoquer des mortalités massives sans exclure que ses effets sublétaux ne puissent contribuer à l'affaiblissement des colonies exposées[175].
Sélection génétique à sens unique
Autre pratique montrée du doigt la sélection intensive d'abeilles uniquement sur des critères de productivité et de faible agressivité à l'aide d'insémination artificielle[176], la quasi-disparition des souches sauvages, l'utilisation de reines produites à la chaîne par un nombre très limité de fournisseurs, l'utilisation systématique de traitements chimiques contre les maladies et les parasites a stoppé l'évolution biologique normale de l'abeille européenne[177]. Cette sélection sauvage s'est faite sans contrôle, avec notamment des importations de reines des pays de l'Est et d'Asie, ce qui explique l'arrivée du Varroa puis de Nosema ceranae, qui proviennent d'abeilles européennes chinoises en contact avec l'abeille asiatique d’Apis cerana. La fixation des caractères par consanguinité est aussi une pratique courante et très risquée :
« Comme on le constate toujours généralement, une consanguinité inconsidérée cause, dans tous les cas, une grave perte de vitalité de l’abeille. Une telle perte s’étend à toutes les capacités et expressions de la vie. Trop souvent, elle met réellement en péril l’existence de la colonie qui en est affectée. Les pertes catastrophiques de colonies, que l’on continue à signaler, sont, le plus souvent, si ce n’est toujours, la conséquence d’une altération de l’élan vital, causée par la consanguinité. Il s’agit, dans tous les cas, d’un défaut insidieux et sournois, qui se révèle lors de conditions climatiques défavorables, auxquelles une constitution affaiblie ne peut plus résister. C’est en de tels moments que la Nature reprend à nouveau le contrôle en éliminant les individus inaptes. La diminution de l’élan vital se manifeste également par une attention amoindrie aux soins du couvain et par une sensibilité accrue à la maladie, aussi bien dans le couvain que chez l’abeille adulte. Comme l’expérience pratique l’a démontré, les espèces d’abeilles les plus productives peuvent être réduites à néant en peu de générations, par la faute d’une consanguinité inconsidérée »
— Frère Adam Karl Kehrle, sélectionneur de la souche Buckfast, Conférence présentée à Paris le 31 janvier 1981[22]
La production dite « sur île », consiste à isoler l'abeille au maximum des éléments pathogènes, comme pour l'élevage de la Buckfast au Danemark. Celle-ci en plus d'être hybride, a des caractéristiques génétiques basées sur un faible nombre d'individus. Bien que productive, cette abeille est très sensible à tout élément extérieur non habituel. De plus, elle a une séquence ADN réduite et instable dans le temps, au contraire d'abeilles plus rustiques.
Cette recherche du rendement a permis de décupler la production par ruche (de 5 à 50 kg) mais s'est traduite par une forte perte de rusticité des colonies. Moins adaptées à l'environnement, les abeilles domestiques sont plus fragiles que celles qui bénéficient d'une génétique plus solide[178]. Dans la nature les reines s'accouplent avec 12 à 26 mâles issus de diverses colonies environnantes ce qui permet un bon brassage génétique[179], sans pour autant impliquer des échanges avec des populations vivant dans des conditions trop différentes. En élevage la sélection intensive vise au contraire à fixer les caractères par la consanguinité. Certains apiculteurs mettent en cause ces abeilles issues de schémas de sélection intensive et de fécondation contrôlée[180],[181]. De nombreux apiculteurs pratiquent le remplacement des reines quand elles sont peu productives[182],[183], or les reines transmettent une partie de leurs immunités à leurs descendances[184], les substitutions à l'aide de reines importées pourraient entraver ce phénomène d'adaptation naturelle aux conditions locales. D'après les scientifiques du Département de Zoologie de l'Université de Graz cette caractéristique pourrait néanmoins ouvrir la voix à des programmes d'amélioration de l'immunité des abeilles via les reines.
Des travaux sont en cours pour améliorer la résistance génétique aux maladies et parasites. En Suède les colonies de l’île de Gotland n'ont subi aucun traitement contre le varroa pendant plus d'une décennie, certaines colonies sont ainsi devenues résistantes. D'après des scientifiques de l'Université Suédoise des Sciences Agricoles[185] ces abeilles ont acquis une résistance multiple au varroa et aux virus associés. Ils ont introduit des ruches non résistantes sur l'île et on comparé les mortalités, les résultats sont sans appel : toutes les ruches non résistante sont mortes pendant l'hiver alors que les colonies résistantes ont survécu malgré une forte charge en varroa et en virus associé.
Qualité du sperme stocké par les reines
Les reines sont fécondées une fois par un nombre important de mâles (>10). En élevage les reines sont généralement inséminées artificiellement avec un nombre plus restreint de mâles sélectionnés par l'éleveur. Une étude récente a montré que les colonies effondrées et celles dont les reines ont été volontairement remplacées par les apiculteurs avaient des reines fécondées avec du sperme peu viable (50-55 % contre 92 %)[186]. La viabilité individuel du sperme des mâles et les conditions thermiques pendant le transport des reines fécondés sont à ce jour les deux seuls facteurs explicatifs connus.
Les pesticides agricoles
Les pesticides sont potentiellement mis en cause, notamment à cause des divers effets sublétaux qu'ils peuvent induire chez les arthropodes non-cibles (comme les abeilles)[187] et leur possible toxicité lors d'exposition chronique[188]. Parmi ces pesticides, certains insecticides (l'imidaclopride (Gaucho)) et d'autres néonicotinoïdes (comme la clothianidine (Poncho Pro)), le fipronil (Régent TS), le thiaméthoxame (Cruiser OSR), les produits de démoustication, etc.) voire des désherbants ou des fongicides, ou des sous-produits de dégradation ou des métabolites de ces produits ayant un effet imprévu sur la capacité de l'abeille adulte à s'orienter (à partir d'un seuil lié à des effets cumulatifs ou synergiques, qui pourraient par exemple concerner toute une classe d'âge de larves, ce qui expliquerait la brutalité du syndrome). Les études standard exigées pour la mise sur le marché de ces produits n'ont jamais montré d'effet, mais leur financement par les industriels de la chimie les rend peu convaincantes aux yeux des environnementalistes. D'autre part, alors que les populations d'abeilles s'effondrent de façon dramatique dans la plupart des pays utilisateurs de pesticides, les populations d'abeilles de l'île de Cuba, où les apiculteurs sont prévenus et anticipent l'épandage de pesticides, se portent très bien et sont en bonne santé.
En 2006, une étude de terrain de l'université de Liège tend à relativiser la corrélation entre les traitements des cultures et la mortalité des ruchers en pointant comme cause première l'acarien Varroa[189].
En 2007, une étude canadienne[190] n'a pas observé d'effet « à long terme » (130 jours) sur la survie, le développement du couvain et la productivité après avoir posé des ruches dans des parcelles de canola (colza de printemps) traité à la clothianidine. Les résidus mesurés étaient 8 à 22 fois inférieurs à la dose sans effet notable observable.
En 2008, l'AFSSA estime le ne pas pouvoir établir la responsabilité des insecticides utilisés en traitement de semence sans pour autant l'écarter[25].
En 2009, une étude belge de terrain confirme l'innocuité de l'imidaclopride en traitement de semence du maïs : elle n'a pas observé de corrélation entre le taux de mortalité et la présence de maïs traité à portée de butinage (3 000 m). Les auteurs signalent cependant une corrélation négative entre le nombre de ruches dans le rucher et la mortalité, ainsi que des usages de produits interdits et/ou inefficaces contre le Varroa[191].
En 2010, des travaux ont montré un effet synergique entre Nosema ceranae et l'imidaclopride[192], mais une contre-analyse des résultats par les auteurs conclut à l'absence de synergie. Nosema ceranae — avec ou sans pesticides — altère gravement l'immunité intestinale de l'abeille, ce qui l'empêche de réagir face à de nouvelle agression[193]. Les abeilles infectées par Nosema et nourries avec un sirop contenant de l'imidaclopride présentent une mortalité supérieure au cumul de mortalité des témoins ne recevant que du sirop contaminé ou l'infection par Nosema ceranae : pour une dose de 7 µg/kg la mortalité du témoin est de 5 %, par l'imidaclopride est d'environ 15 % en 10 jours, 27 % avec Nosema ceranae seul et elle atteint 47 % en combinant les deux effets, ce qui est un peu supérieur à un simple effet additif. L'expérience montre que les abeilles atteintes par Nosema ceranae consomment plus de sirop, et que cet effet semble amplifié par l'insecticide, ce qui augmente d'autant plus leur contamination par l'insecticide mais aussi les risques que les abeilles encourent en partant butiner.
En 2012, deux études séparées publiées simultanément dans Science concluent que les pesticides (respectivement le thiaméthoxame et l'imidaclopride) représenteraient bien un danger pour les abeilles et les bourdons[194],[195],[196]. L’étude est critiquée car utilisant des doses de pesticides trop élevées en comparaison de l’exposition naturelle[197] et pour des problèmes statistiques et une mauvaise prise en compte de la mortalité naturelle lors du butinage et lié au stress de l’expérience (abeilles relâchées dans un environnement inconnu)[198],[199]. L'ANSES et l’EFSA concluent que les doses étaient effectivement non représentatives, sans remettre en cause les conclusions des deux études et en recommandant des expérimentations complémentaires avant d’envisager une remise en cause des autorisations de mise sur le marché des préparations incriminées[200],[201]. Une étude allemande dirigée par Johannes Fischer reprenant un protocole très proche mais avec des doses plus réalistes ne parviendra pas à reproduire les effets observés par l'équipe de l'INRA d'Avignon, confirmant en revanche la pertinence du protocole utilisant des puces RFID pour observer les effets sublétaux des doses plus élevées[202]. L'un des auteurs, Mickaël Henry, de l'INRA d'Avignon, répondra à ces critiques en décembre 2013[203] sur l'interprétation statistique des résultats, tout en admettant les faiblesses du protocole au niveau de la dose (la dose journalière maximum fut donnée en une prise, donc subie sur un seul vol, les auteurs n'ont pas eu d'autres possibilités techniques pour simuler l'exposition récurrente), la prise en compte de la mortalité naturelle (le butinage est une activité risquée que les abeilles effectuent en fin de vie) et du stress du à l'environnement inconnu imposé par l'expérience (qui suffit pour provoquer un taux de non-retour anormal chez les témoins : 16,9 % contre 1,5 % seulement pour le butinage dans un environnement familier pour l'abeille). En avril 2012, l’INRA prépare une expérimentation en conditions réelles[204]. Des chercheurs de l’Université d'Exeter, financés par l'ONG Society of Chemical Industry, concluent dans la revue Pest Management Science que les néonicotinoïdes ne peuvent être mis en accusation[205] Une synthèse faite par l'université de Wageningue[206] met en relief les limites des connaissances actuelles sur les néonicotinoïdes et estime que les taux mesurés dans l'environnement sont toujours plus faibles que ceux induisant une toxicité aiguë ou chronique. Pour cette synthèse, les études qui montrent des effets comportementaux ne sont pas confirmées par les essais sur le terrain avec des doses correspondant à ce qui est mesuré dans les plantes traitées.
En 2013, le Département de l'Environnement, de l'Alimentation et des Affaires rurales du Royaume-Uni conclut (en mars), sur la base d'une étude de terrain limitée et menée sur des bourdons que ces insecticides n'ont pas eu d'effet négatif démontrable in situ sur la croissance des colonies. Il remet également en cause les résultats expérimentaux de laboratoire des études précédentes en raison de biais de protocoles, mais sans nier la dangerosité des insecticides pour les colonies[207]. En octobre, des chercheurs de Syngenta publient sur PLOS ONE une « Étude de terrain sur 4 ans des effets à long terme d'une exposition répétée de colonies d'abeilles à des cultures à fleurs traitées au thiamethoxam ». Deux protocoles sont utilisés : 1) une exposition forcée en cage, et 2) la pose des ruches très près d'un champ traité (maïs et colza) dans plusieurs sites français. Les auteurs ont conclu que les dangers du thiamethoxam sur les abeilles sont faibles[208]. Une synthèse menée par Jeroen van der Sluijs de l'université d'Utrecht (avec des chercheurs de l'université du Sussex, du CNRS et d'INRA) et publiée dans la revue Current Opinion in Environmental Sustainability rappelle en 2013 que les néonicotinoïdes sont de plus en plus utilisés (plus de 25 % de parts de marché vers 2012) et que leur large diffusion dans l'environnement et leur rémanence dans le sol et l'eau les rend bio-disponibles à dose sublétale toute l'année pour les pollinisateurs. Ces néonicotinoïdes sont fréquemment trouvés dans les essaims d'abeilles. Les auteurs concluent que « les données limitées disponibles suggèrent qu'ils sont susceptibles de présenter une toxicité similaire pour pratiquement tous les autres insectes pollinisateurs sauvages » et « soumis à des doses réalistes de terrain, ils provoquent une large gamme d'effets néfastes pour l'abeille et le bourdon, en affectant la performance des colonies, affaiblissant le butinage et le couvain, augmentant la fragilité face aux maladies et aux parasites ». Les auteurs plaident pour une transition vers des alternatives aux néonicotinoïdes sans effets sur les pollinisateurs[129]. Fin 2013 et début 2014 deux études[209],[210] sur l'effet de l'imidaclopride et du thiaméthoxame chez le bourdon terrestre sont publiées par l'Université d’Exter : des bourdons ont été exposés 14 jours (durée comparable à une floraison) à une alimentation contaminée puis nourris 14 jours par des aliments "propres" :
- les effets du thiaméthoxame ne sont visibles qu'à forte dose (39 et 98 μg·kg-1), aux doses mesurées dans des conditions réalistes de culture les colonies ne sont pas affectées (entre 1 et 11 μg·kg-1) ;
- l'imidaclopride a un effet répressif sur l'alimentation plus puissant que le thiaméthoxame, notamment sur la consommation de pollen, ce qui expliquerait son effet plus fort sur la dynamique de population de la colonie : dès qu'1 μg·kg-1 est dépassé la production de couvain chute ;
- durant la période d'« alimentation propre », les colonies retrouvent rapidement une dynamique de croissance, après 28 jours il n'est plus possible de différencier les colonies.
Ceci semble disculper le thiaméthoxame (Cruiser) et démontrer un effet réel de l'imidaclopride (Gaucho) (mais non persistant chez le bourdon). L'extrapolation de ces résultats aux abeilles domestiques ne peut être réalisée sans travaux équivalents. Pour l'agronome et ancien PDG d'Aventis CropScience Alain Godard, un autre élément remet en cause la thèse de l'empoisonnement généralisé par les pesticides systémiques : dans les années 1970 des doses massives d'insecticides très toxiques (notamment plus toxiques pour les pollinisateurs[211]) et très rémanents comme le lindane furent utilisées sur les cultures, sans aucune précaution, sans que des mortalités massives et systématiques soient constatées. Pour Alain Godard, « si les abeilles avaient dû disparaitre de par les effets des pesticides, c’est bien à cette époque que cela aurait dû avoir lieu »[212]. À cette époque les accidents n'étaient pas rares, dans une synthèse de l'INRA publiée en 1984, les auteurs décrivaient déjà les dégâts subis par l'apiculture dans les années 1950, des milliers de ruches avaient été détruites par les insecticides organochlorés et des statistiques américaines chiffrent à 70 000 la destruction de colonies d'abeilles en Californie, pour la seule année 1967, en raison de l'usage agricole de pesticides[213]. Mais ces chiffres certes impressionnants ne sont pas du tout à la même échelle que le CCD, car ils s'expliquaient alors par des contaminations très localisées d'une ruche ou d'un rucher par un traitement mal positionné ou trop dosé, et les apiculteurs identifiaient aisément l'origine du problème. Le syndrome d'effondrement des colonies d'abeilles concerne un nombre de ruches considérablement plus élevé : le déclenchement de la crise en 2006 a provoqué la perte de 1,5 million de colonies aux États-Unis en quelques mois, soit plus de 60 % du cheptel[214], les mortalités récurrentes du CCD éliminent chaque année des millions de ruches en Europe.
En 2014, alors que le phénomène d'effondrement se poursuit aux États-Unis[215], le débat sur les doses et les effets cumulés sur le long terme continue, car la barrière entre la dose sans effet et la dose perturbant les abeilles est mince, et pourrait être dépassée dans les plus mauvais scénarios d'exposition[216] d'après Johannes Fischer (mais la significativité des résultats de l'étude de 2013 est faible, les barres d'erreurs se recoupant sur la quasi-totalité des mesures). Une étude de Chensheng Lu de l'Université de Harvard semble montrer qu'une exposition sublétale quoique très largement supérieure aux doses homologuées (1,9 litre de sirop de sucre contenant 135 µg/l d'insecticide par colonie et par semaine, valeurs 3 à 100 fois plus élevées que celles mesurées dans le nectar[206]) et sur une période d'exposition anormalement longue (13 semaines, soit 4 fois plus longtemps qu'une floraison classique) de néonicotinoïdes (imidaclopride ou clothianidine) affectent bien l'hivernage des colonies saines qui y ont été exposées. Elle conduit à une mortalité de 50 % avec des symptômes évoquant la CCD chez les colonies exposées en fin d'hiver, bien qu'aucun symptôme particulier ne soit observé en été ou en automne. Dans les colonies non exposées, les abeilles mortes ont été rapidement remplacées par de nouvelles abeilles émergentes. Une seule des six colonies de contrôle a été perdue en raison d'une infection à Nosema. Ces résultats seront très critiqués en raison du protocole utilisant des doses très élevées[217] et l'auteur accusé de desservir la cause en alimentant le débat avec de la mauvaise science[218], renforçant la confusion.
En 2015, une étude[219] sur trois ans de l'Université du Maryland rapporte l'absence de mortalité anormale après l'exposition à des doses normales d'imidaclopride. Les premiers signes apparaissent avec des doses 4 fois supérieures à la réglementation et les mortalités deviennent massives à 20 fois la dose. D'après les auteurs ces travaux n'absolvent pas totalement les pesticides mais excluent qu'ils puissent à eux seuls expliquer la crise. Il est à noter que les ruches bénéficiaient d'une alimentation optimale et n'étaient pas exposées à d'autres pesticides. Une autre étude semble confirmer l'absence d'effet synergique entre les pesticides agricoles et les maladies parasitaires de l'abeille[220].
Influence des néonicotinoïdes sur la dynamique de butinage
Certains nectars floraux contiennent naturellement de petites quantités d'alcaloïdes (substances défensives pour la plante), notamment de la nicotine, molécule dont sont dérivés les insecticides de la famille des néonicotinoïdes. Ces molécules naturelles sont toxiques pour les abeilles en cas d'exposition répétée, mais tolérées cependant par les abeilles domestiques aux doses où elles sont présentes dans la nature[221]. La qualité du nectar floral, elle-même influencée par le contexte agro-écologique, influence les capacités des abeilles à les détecter[221]. Fin 2005, une étude a montré que les abeilles préféraient un sucre légèrement enrichi en nicotine qu'un sucre normal[222]. Cette attirance s'expliquerait par l'effet antiparasitaire des alcaloïdes[223].
Les nectars des plantes traitées par des pesticides systémiques peuvent contenir des doses détectables et mesurables de ces pesticides de synthèse si ces plantes proviennent de semences traitées ou qu'elles ont été directement traitées, il n'y a pas de trace détectable dans les cultures suivantes[224].
En janvier 2015, une étude[225] publiée par la revue Nature montre qu'en laboratoire, les abeilles comme les bourdons ont une préférence pour les mélanges de sucre et de pesticide néonicotinoïde par rapport à du sucre brut. Les auteurs en concluent que les pollinisateurs pourraient être plus significativement exposés aux insecticides dans la nature qu'on ne le pensait jusqu'alors, les abeilles pouvant littéralement activement « rechercher les insecticides néonicotinoïdes nocifs dans leur environnement »[226]. En octobre 2015, une nouvelle étude, dont deux des auteurs avaient participé à l'étude de janvier, remet en cause cette thèse. D'après ces nouvelles données, il apparaitrait que la mémoire à court et long terme (uniquement dans le cas de l'imidaclopride) des abeilles est perturbée par le nectar contaminé. Les scientifiques concluent que les néonics pourraient affecter l'efficacité du butinage des abeilles au lieu de les inciter à récolter préférentiellement du nectar contaminé.
Les pesticides à usage vétérinaire
Pour lutter contre Varroa et d'autres pathogènes, des pesticides et des antibiotiques sont utilisés. Les produits les plus courants sont :
- le coumaphos, un organophosphoré interdit en Union Européenne, utilisé contre Varroa ;
- Le tau-fluvalinate, un pyrèthre de synthèse (lointain) dérivé d'une protéine du lait[227], la valine, contre Varroa ;
- L'amitraze utilisé contre Varroa, autorisé en France[228] (Apivar) ;
- L'oxytétracycline contre la loque américaine ;
- L'acide formique, autorisé en apiculture biologique contre Varroa ;
- Le thymol, de l'huile essentielle de thym, autorisé en apiculture biologique contre Varroa.
D'autres voies sont explorées :
- Une équipe égyptienne a testé avec succès deux champignons, le Metarhizium anisopliae le Beauveria bassiana[229]. Les auteurs concluent que leur efficacité et l'absence d'effet négatif sur les ruches en font des biopesticides potentiels efficaces contre le Varroa.
- Des travaux bulgare[230] et brésilien[231] montrent que la composition de la propolis des abeilles résistantes au Varroa est différente et que leur comportement est différent : meilleur comportement hygiénique, stockage de pollen et de miel supérieur que les lignées sensibles.
Ces pesticides, même d'origine naturelle[232], s'ils ne semblent pas dépeupler ou affaiblir les ruches aux doses recommandées, ne sont pas pour autant sans effets sur les abeilles[233], provoquant notamment des mortalités directes en cas de surdosage[234]. Ces produits peuvent avoir des effets sublétaux : en s'accumulant dans la cire, se mélangeant aux produits phytosanitaires agricoles (fongicides et insecticides) ils provoqueraient d'une part un ralentissement de la croissance du couvain, ce qui pourrait rendre ce dernier plus sensible à l'infection par le Varroa, et d'autre part une réduction de la survie des adultes[235].
Ils agissent aussi en synergie ou addition avec les produits de traitement utilisés contre Varroa : des abeilles expérimentalement exposées à l'Apistan (Fluvalinate) meurent 1,9 fois plus quand elles sont exposées ensuite à de la bifenthrine (insecticide de la même famille) alors qu'il n'y a pas de différence significative de mortalité pour celles qu'on expose ensuite au carbaryl et au parathion méthyl[236]. Le fluvalinate seul provoque une réduction de la capacité d'apprentissage, de mémoire, de réaction au saccharrose et une réduction du taux de survie[237]. L'amitraze modifie durablement le rythme cardiaque des abeilles exposés[238]. Certains produits de traitement comme la tétracycline et l'insecticide à base de methoxyfenozide (pourtant présenté comme sans effet sur les pollinisateur) et une combinaison de fluvalinate et de coumaphos réduisent la proliférations des cellules souches de l'intestins, ce qui pourrait affecter les capacités de défense immunitaire des abeilles[239]. L'efficacité de ces produits est faible, une étude américaine[233] a montré qu'ils réduisent la survie du couvain, affectent la production de reines, sans pour autant améliorer de façon significative la survie des abeilles. Les auteurs concluent qu'il est important de minimiser l'usage de ces produits.
Synergie entre pesticides agricoles et vétérinaires
Une étude américaine de 2010 menée par les universités du Nebraska et de Pennsylvanie affirme que si en l'état des connaissances un pesticide unique ne peut être incriminé comme cause du syndrome, le mélange entre les produits de traitement de la ruche et les traitements agricoles pourraient générer des phénomènes de toxicité synergique qui pourraient contribuer au phénomène[240]. Certains effets sublétaux attribués aux néonicotinoïdes pourraient aussi être provoqués par les produits de traitement contre le Varroa destructor : les abeilles traitées avec du tau-fluvatinate voient leur butinage perturbé, effet semblable à celui de l'imidaclopride[241]. La contamination de la cire par des produits toxiques d'origine apicole, agricole et industrielle qui s'y accumulent et restent présents pendant des décennies pourrait aussi affaiblir durablement les essaims[242] et provoquer des mortalités massives en cas d'exposition à des traitements sans danger dans des ruches « propres ». Aux États-Unis, l'effet cumulé des traitements contre Varroa destructor (Coumaphos et tau-fluvalinate), un antibiotique et d'autres pesticides ont été mis en cause en 2011[243]. Par contre la combinaison de coumaphos et d'imidaclopride ne semble pas provoquer d'effet synergique[244].
Une autre étude[245] met en évidence des effets contrastés mais en général additifs entre 4 pesticides communément présents dans les ruches : fluvalinate, coumaphos, chlorothalonil, et chloropyrifos. Les deux premiers sont des produits de traitement contre Varroa, le coumaphos est interdit en UE, le chlorothalonil un fongicide utilisé sur une large gamme de cultures (blé, orge, poids[246][réf. incomplète]), enfin le chloropyrifos, de la même famille que le coumaphos, est un insecticide organophosphoré d'ancienne génération[247]. Ces produits provoquent un doublement de la mortalité des larves, le plus dangereux semble être le chlorothalonil. L'étude des mélanges montre des effets variants de la synergie (d'une part entre le chlorothalonil et le fluvalinate pour une concentrations de 34 mg/L et 3 mg/L, et d'autre part entre le chlorothalonil à 34 mg/L et le coumaphos à 8 mg/L) à l'antagonisme selon la dose (mélange chlorothalonil et fluvalinate, mais concentration réduite d'un facteur 10). L'addition de coumaphos réduit significativement la toxicité du mélange de fluvalinate et de chlorothalonil, tous les autres effets sont additifs. L'effet d'un solvant, le N-méthyl-2-pyrrolidone, commun dans la formulation de certains pesticides[248], montrent qu'une exposition chronique au mélange de toutes ces molécules peut potentiellement affecter les populations d'abeilles et nécessite des recherches supplémentaires. Les auteurs[Qui ?] concluent que la toxicité des molécules présentent dans l'alimentation des abeilles doit être calculée par addition des toxicités de chaque molécule[249].
Les résultats de cette étude sont à considérer avec prudence car les doses testées sont très supérieures à celles mesurées couramment dans les ruches : pour le coumaphos et le fluvalinate par exemple les doses courantes sont de l'ordre de 1 à 2 mg/kg, ce qui est très inférieur aux doses considérées comme dangereuses dans l'étude[250],[251], ces deux produits sont de surcroît rarement associés en Europe, le coumaphos étant interdit[252]. Néanmoins la concentration de ces produits peut varier en fonction des modalités de traitement : des pics de concentration sont mesurés durant les phases de traitement, avec des chiffres très élevés si le traitement est maintenu dans la ruche sur de longue période[253], dans ce cas les valeurs cités dans l'étude peuvent être ponctuellement atteintes, avec un risque certain pour la colonie.
Causes environnementales possibles

- Un facteur environnemental non compris, qui pourrait par exemple impliquer le dépassement d'un seuil de bioaccumulation d'un (ou plusieurs) polluant, éventuel perturbateur phéromonal, avec pour effet inattendu de perturber la capacité à retrouver leur ruche.
- Une synergie entre plusieurs des causes évoquées ci-dessus pourrait être à l'origine d'un syndrome de type dit « maladie environnementale »[254].
- C'est l'hypothèse qui semble la plus probable, médiatisée par exemple en 2007 par le professeur Joe Cummins de l'Institute of Science in Society qui met en cause à la fois des champignons parasites utilisés en lutte intégrée agricole, des virus, bactéries et la pollution électromagnétique croissante à laquelle sont exposés les insectes (notamment depuis l’avènement du téléphone portable et du Wi-Fi). Selon lui, ces causes combinées affaibliraient le système immunitaire des abeilles, mais le rôle des pesticides reste pour partie nié par les industriels producteurs[255].
- La pollution par les moteurs diesels est évoquée par l'Université de Southampton[256] : les micro-particules pourraient affecter le cerveau des insectes. Cette hypothèse n'a pas encore fait l'objet de test expérimentaux.
- L'urbanisation et les pratiques d'élevages augmenteraient la pression parasitaire, des chercheurs de l'Université de Caroline du Nord ont remarqué que les abeilles ouvrières "sauvages" et éloignées des espaces urbanisés étaient trois fois moins susceptible de mourir prématurément[257].
Organismes génétiquement modifiés
Les OGM ont également été rapidement accusés[258], car leur culture en plein champ dans certains pays a précédé de peu ce nouveau syndrome, et parce que de nombreux OGM ont été génétiquement modifiés pour produire leur propre insecticide. Par exemple, le pollen vivant de nombreux maïs transgéniques exprime la protéine transgénique insecticide Bt. Une étude[259],[260] de l'Université d'Iéna a porté (de 2001 à 2004) sur l’effet des pollens GM exprimant la toxine Bt sur les abeilles. Comme annoncé par les fabricants, sur les individus sains, aucun effet toxique du pollen n’a été démontré, mais les abeilles affectées par un parasite s'y sont montrées beaucoup plus sensibles, la mortalité étant alors beaucoup plus élevée chez les abeilles expérimentales nourries au pollen GM (durant 6 semaines, durée très supérieure à la floraison des plantes OGM). Les chercheurs supposent que le pollen GM pourrait affecter l'immunité de l'abeille, le Pr Hans-Hinrich Kaatz (université de Halle) estime que le Bt pourrait ne pas tuer l'abeille mais agresser les cellules des parois de son intestin, facilitant l'infection par d'autres pathogènes. Dans les 6 semaines d'expérience, les abeilles nourries avec des doses importantes de Bt (10 fois la dose normale) ont été plus nombreuses à mourir. La reine y est exposée plus longtemps, et l'effet sur plusieurs générations de larves n'a pas été étudié. Une étude de mars 2013 confirme l'absence d'impact du pollen BT et montre aussi la présence de protéines Cry provenant de sources naturelles[261].
Cette hypothèse n'est généralement pas retenue comme cause de la disparition des apidés, car des abeilles disparaissent touchées dans des zones où les OGM sont très rares ou totalement absents[262]. Enfin, l'Europe où les OGM sont peu présents a été touchée avant l'Amérique du Nord où ils sont le plus cultivés. Si les OGM produisant leur propre insecticide peuvent avoir des effets néfastes sur les abeilles, les insecticides « classiques » largement utilisés par les agriculteurs en ont également, et l'usage d'OGM a permis d'en réduire l'usage. Deux études concluent ainsi que les champs d'OGM Bt (coton Bt et maïs Bt) auraient plutôt moins d'effet que ceux traités par pulvérisation d'insecticides classiques, sur les abeilles proprement dites[263],[264]. Une méta-analyse indépendante de 2007 affirme que ces derniers sont hors de cause[265].
Autres hypothèses
- Plantations massives d'arbres et de fleurs très résistants souvent d'origine exotique dont le pollen s'avère toxique pour les abeilles et bourdons locaux. C'est le cas en France pour le Tilleul à larges feuilles (Tilia platyphyllos), le Tilleul argenté (Tilia tomentosa) et le Tilleul de Crimée (T. x euchlora) souvent planté en milieu urbain pour sa résistance.[réf. nécessaire] Début juillet, en ville, on trouve beaucoup d'abeilles et de bourdons morts sur l'asphalte des rues plantées de ces essences de tilleuls.[citation nécessaire] On a parfois compté jusqu'à 200 insectes morts par jour et par arbre[266]. Si ces végétaux peuvent localement affecter des ruchers ayant un environnement peu propice, l'effondrement des colonies est un phénomène global, très variable d'une année sur l'autre, cette hypothèse ne peut être retenue.
- Les lignes hautes tensions[267], les relais téléphoniques[268], pour cause des fortes ondes électromagnétiques que ces dispositifs produisent, font également partie des hypothèses régulièrement mises en avant.
Zones préservées
Les îles
L’île d’Ouessant, à l'extrême ouest des côtes françaises. Les pesticides y sont rares et les abeilles se portent bien. En effet, les mortalités hivernales dans les ruches y sont mesurées à 3 % en moyenne contre 30 % à 40 % sur le continent, à tel point que l’abeille noire d’Ouessant est multipliée pour remplacer celles qui disparaissent ailleurs en France[269] mais il faut aussi noter l'absence du varroa destructor sur ces abeilles (voir ci-dessus). Ouessant fait partie de la Réserve de biosphère Îles de la mer d'Iroise délimitée en 1988 par UNESCO[270].
Le contexte insulaire de Cuba est favorable aux abeilles où l'on observe une population saine[271].
Pollinisateurs sauvages
Parallèlement aux déclins des abeilles domestiques européennes, les pollinisateurs sauvages voient aussi leurs populations affectées[272]. Ce phénomène peut être lié à une dégradation des habitats, au changement climatique et l'utilisation de produits phytosanitaires dans l'agriculture[273]. La dégradation des habitats comprend notamment le déclin de la richesse florale : si certaines espèces polyvalentes comme nos abeilles domestiques peuvent profiter des cultures agricoles mellifères[274], d'autres sont très spécialisées et leurs populations évoluent parallèlement avec l'abondance de leurs sources de nourriture. Enrichir le paysage en diversifiant les espèces cultivés, créant des habitats de hautes qualités permet d'améliorer considérablement l'état des populations de pollinisateurs sauvages[275].
Des scientifiques ont récemment pointé du doigt l'influence de l'état sanitaire des abeilles domestiques, notamment leur rôle dans la transmission des maladies et parasites émergentes : une étude anglaise de 2013[276] montre que Nosema ceranae est capable d'infecter le bourdon terrestre : de nombreux individus capturés dans la nature sont infectés et des expérimentations montrent que le pathogène réduit la survie de 48 % et provoque des effets sublétaux comportementaux.
Dans un article publié en février 2014 dans la revue Nature[277], une équipe européenne a montré que les bourdons sauvages en contact avec des ruches d'abeilles domestiques ou des colonies de bourdons d'élevage sont atteints par les mêmes lignées du virus des ailes déformées et avec des taux de prévalences semblables. Les auteurs préviennent que les ruches d'abeilles d'élevages pourraient être une source majeur de transmission des maladies émergentes chez les pollinisateurs sauvages et pourrait être une cause de leur déclin.
Enfin, les pesticides peuvent nuire considérablement aux bourdons dans le monde développé[194]. L'étude Chronic sublethal stress causes bee colony failure sur les bourdons publiée dans Ecology Letters le 6 octobre 2013 a permis de démontrer que l'exposition des bourdons terrestres à un insecticide à une dose comparable aux doses les plus élevées mesurées dans du nectar de plante traité peut provoquer la mort des colonies[278]. D'après le journaliste au Monde, Stéphane Foucart cette étude montrerait que« l'exposition à des doses très faibles de néonicotinoïdes peut conduire les colonies d'insectes sociaux au déclin », pour autant, note un apidologue français « ces travaux ne constituent pas une preuve biologique mais sont plutôt un outil qui permet éventuellement de comprendre comment les choses peuvent se passer. [...] Une dizaine de modèles analogues ont déjà été construits »[279].
En 2015 une étude universitaire[280] publiée dans la revue Science démontre que la réduction de l'aire de répartition (et non la densité de population) des bourdons s'expliquent par le changement climatique. De nombreux espèces ne parviennent pas à déplacer leur aire de répartition aussi vite que le déplacement de leurs plantes nourricières. En juin 2015 une étude réalisée par 58 chercheurs[281] et publiée dans Nature permet de mieux cerner la problématique. Les pollinisateurs sauvages en déclin ne sont pas les espèces, les plus communes, qui jouent un rôle significatif dans la pollinisation des cultures. Les auteurs notent que ces espèces résistent bien à l'environnement agricole et que quelques aménagements peu couteux suffiraient pour renforcer ces populations. Les auteurs suggèrent d'ajuster la communication et les méthodes de conservation pour mieux cibler les espèces sauvages réellement menacées.
Progrès de la modélisation
Pour comprendre un phénomène multifactoriel où de nombreux phénomènes additifs ou synergiques sont évoqués, la modélisation est une des voies de recherche pertinente pour synthétiser les connaissances : elle permet, en s'appuyant sur des résultats isolés, de tester rapidement de nombreux paramètres et surtout les très nombreuses combinaisons entre ces paramètres. Plusieurs modèles mathématiques ont tenté de simuler les facteurs multiples qui expliqueraient le déclin de l'abeille domestique[282],[283], mais si les Building block model (en) de ces modèles sont individuellement bien validés par l'expérimentation[284], aucun ces modèles ne prend pas en compte simultanément la dynamique interne de la colonie (génétique, qualité de la reine, population), le comportement de recherche de nourriture dans un environnement réaliste (distance et qualité des ressources) et les facteurs de stress internes et externes (climat, maladies, parasites et expositions chimiques)[285]. Au niveau européen, le programme BEEHAVE, soutenu par l'Université d'Exeter, l'université de Warwick, le Centre de recherche environnementale de Helmholtz et la firme Syngenta, vise à mettre au point un modèle unique permettant de simuler l'effet des facteurs de stress sur des colonies virtuelles dans un environnement paramétrable. Les premiers résultats du modèle, non publiés[286], confirmeraient l'effet cumulatif de plusieurs facteurs (Varroa, la distance des ressources alimentaires et le virus des ailes déformées), mais aussi un délai de plusieurs années entre l'introduction du facteur de stress et la mort effective des colonies.
Par ailleurs,on trouve des plateformes opérationnelles de recensement des mortalités massives telles que la chaîne de solidarité sur la surveillance des abeilles ApiVigi, qui permettent de collecter et trier toutes les observations à partir de signalements informatiques effectués sur le site apivigi.com, pour les mettre à disposition de tous.
Références
- [PDF] Mortalités, effondrements et affaiblissements des colonies d’abeilles, sur le site anses.fr
- [PDF] Mortalités, effondrements et affaiblissements des colonies d’abeilles, sur le site beekeeping.com
- [PDF]+(en) Colony Collapse Disorder Progress Report, sur le site usda.gov de juin 2010
- (en) Bernard Vaissière et al., « Global honey bee colony disorders and other threats to infect pollnitors », Programme des Nations unies pour l'environnement, (lire en ligne [PDF])
- (en) Michael McCarthy, « Decline of honey bees now a global phenomenon, says United Nations », The Independent, (lire en ligne)
- Servel 2002
- Dupont 2007
- Chauveau 2009
- Johnson 2007
- Les abeilles sauvages, reines de la pollinisation, sur le site larecherche.fr du 5 mars 2013
- On achève bien les abeilles, sur le site courrierinternational.com du 22 février 2007
- L'étau se resserre autour des "tueurs" de l'abeille, sur le site lemonde.fr du 18 septembre 2009
- Pesticides et surmortalité des abeilles, sur le site sciences-po.fr
- Disparition des abeilles, la fin d’un mystère de Natacha Calestrémé, Montparnasse, 7 juin 2011, 64 min (EAN 3346030023228)
- Notre poison quotidien de Marie-Monique Robin, Arte, 23 mars 2011, 112 minutes
- L'UNAF demande le retrait immédiat des produits phytosanitaires toxiques pour les abeilles, sur le site actu-environnement.com
- Éditorial, « Pesticides : pitié pour les abeilles ! », Le Monde, , p. 1 (ISSN 0395-2037)
- Cox-Foster 2007
- vanEngelsdorp et al. 2009
- Le Rucher dépose plainte, sur ladepeche.fr du 6 octobre 1998, consulté le 5 juin 2017
- Les abeilles meurent toujours, sur ladepeche.fr du 4 avril 1999, consulté le 5 juin 2017
- Une contribution à l’élevage des abeilles : l’exemple de l’abeille buckfast, sur le site pedigreeapis.org
- SDite de statistiques de la FAO
- [PDF] Une activité apicole de plus en plus professionnelle, sur le site gouv.fr
- Agence française de sécurité sanitaire des aliments 2009
- [PDF] BILAN DE L’ENQUETE REGIONALE : Pertes de colonies d’abeilles pendant l’hiver 2012-2013, sur le site alsace.chambagri.fr
- MAAREC 2007
- vanEngelsdorp et al. 2008
- FAOStat, sur fao.org
- « Bee-renting Businesses Buzzing | BC Business », sur bcbusiness.ca (consulté le )
- Tardieu 2009
- Possley 2007
- Frazier 2007
- Dumas 2007
- « Remarks as Prepared for Delivery by Secretary Vilsack at the Stakeholder Workshop on Coexistence in Raleigh, North Carolina | USDA Newsroom », sur www.usda.gov (consulté le )
- Potts et al. 2010
- COST 2008
- van der Zee 2012
- Holzmann 2011
- vanEngelsdorp et al. 2007
- vanEngelsdorp et al. 2010
- vanEngelsdorp et al. 2011
- vanEngelsdorp et al. 2012
- Chauzat MP, Cauquil L, Roy L, Franco S, Hendrikx P, Ribière-Chabert M.PLoS One. Demographics of the European apicultural industry. Nov 13;8(11):e79018. doi: 10.1371/journal.pone.0079018. eCollection 2013. lire en ligne
- CCD Steering Committee, « Colony Collapse Disorder Progress Report », United States Department of Agriculture, (consulté le )
- « Multiple causes for colony collapse – report », 3 News NZ, (lire en ligne [archive du ], consulté le )
- Almudena Cepero, Jorgen Ravoet, Tamara Gómez-Moracho, José Luis Bernal, Maria J. Del Nozal, Carolina Bartolomé, Xulio Maside, Aránzazu Meana, Amelia V. González-Porto, Dirk C. de Graaf, Raquel Martín-Hernández et Mariano Higes, « Holistic screening of collapsing honey bee colonies in Spain: a case study », BMC Research Notes, vol. 7, , p. 649 (ISSN 1756-0500, PMID 25223634, PMCID 4180541, DOI 10.1186/1756-0500-7-649)
- Mollier 2009
- Olivier Pauchard, « Pas de solution miracle contre le varroa », sur www.swissinfo.ch
- « Les paysans et les apiculteurs veulent un plan de mesures », La Tribune de Genève, (ISSN 1010-2248, lire en ligne)
- Andreas Keiser, « Un indigné qui a lutté, seul, contre tous », sur www.swissinfo.ch
- « Un plan pour protéger les abeilles des pesticides », Le Temps, (lire en ligne)
- (en)[PDF]Report on the National Stakeholders Conference on Honey Bee Health, sur usda.gov, consulté le 5 juin 2017
- (en) « Spotlight: BEE DOC », sur coloss.org, (consulté le )
- « Santé des abeilles : impact de la co-exposition des colonies aux pesticides et aux agents infectieux », sur anses.fr, (consulté le )
- (en)Bees Vanish, and Scientists Race for Reasons, sur nytimes.com du 24 avril 2007, consulté le 5 juin 2017
- (en)Bee Colony Collapse Disorder: Could It Be Parasites, Pathogens Or Pesticides?, sur sciencedaily.com du 12 mai 2007, consulté le 5 juin 2017
- (en)Immune pathways and defence mechanisms in honey bees Apis mellifera, sur wiley.com du 27 octobre 2006, consulté le 5 juin 2017
- Géraldine Boezio et Pascal Peduzzi, « Rapport du Programme des Nations Unies sur la mortalité des abeilles et sur les menaces qui pèsent sur les insectes pollinisateurs », Revue Semestrielle de Droit Animalier, no 2, , p. 172 (lire en ligne).
- (en) Dainat B, Evans JD, Chen YP, Gauthier L, Neumann P., « Predictive markers of honey bee colony collapse », PLoS One, vol. 7, no 2, , e32151. (PMID 22384162, DOI 10.1371/journal.pone.0032151, lire en ligne [html])
- A propos du risque d'introduction de Varroa sur l'Ile de la Réunion, sur apivet.eu du 28 septembre 2012, consulté le 5 juin 2017
- Oriane Raffin, « Ils tentent de sauver les abeilles= », sur arte.tv, Arte, (consulté le )
- Dominique Raizon, « L'Abeille Noire d’Ouessant est en pleine forme », RFI, (lire en ligne)
- (en) Predictive Markers of Honey Bee Colony Collapse, sur le site plosone.org, consulté le 5 juin 2017
- (en)Winter Survival of Individual Honey Bees and Honey Bee Colonies Depends on Level of Varroa destructor Infestation, sur le site plosone.org
- (en)Comprehensive Bee Pathogen Screening in Belgium Reveals Crithidia mellificae as a New Contributory Factor to Winter Mortality, sur nih.gov
- (en) Francis RM, Nielsen SL, Kryger P., « Varroa-virus interaction in collapsing honey bee colonies », PLoS One, vol. 8, no 3, , e57540 (PMID 23526946, DOI 10.1371/journal.pone.0057540, lire en ligne [html])
- (en) Genersch E, Aubert M., « Emerging and re-emerging viruses of the honey bee (Apis mellifera L.) », Vet Res, vol. 41, no 6, , p. 54 (PMID 20423694, DOI 10.1051/vetres/2010027, lire en ligne [html])
- Varroa destructor is the main culprit for the death and reduced populations of overwintered honey bee (Apis mellifera) colonies in Ontario, Canada, sur apidologie.org de aout 2010, consulté le 5 juin 2017
- Varroa destructor is the main culprit for the death and reduced populations of overwintered honey bee (Apis mellifera) colonies in Ontario, Canada, sur apidologie.org du 8 janvier 2010, consulté le 5 juin 2017
- Jane P. Staveley, Sheryl A. Law, Anne Fairbrother et Charles A. Menzie, « A Causal Analysis of Observed Declines in Managed Honey Bees (Apis mellifera) », Human and Ecological Risk Assessment, vol. 20, , p. 566-591 (ISSN 1080-7039, PMID 24363549, PMCID 3869053, DOI 10.1080/10807039.2013.831263, lire en ligne, consulté le )
- (en)Varroa destructor (Anderson and Trueman, 2000); the change in classification within the genus Varroa (Oudemans, 1904), sur nih.gov, consulté le 5 juin 2017.
- l'invasion du varroa destructor, sur apistory.fr, consulté le 5 juin 2017
- (en) Anderson DL, Trueman JW, « Varroa jacobsoni (Acari: Varroidae) is more than one species », Exp Appl Acarol, vol. 24, no 3, , p. 165-89. (PMID 11108385)
- (en)New asian type of varroa destructor, sur apidologie.org, consulté le 5 juin 2017
- (en) Y. Le Conte, Z. Y. Huang, M. Roux et Z. J. Zeng, « Varroa destructor changes its cuticular hydrocarbons to mimic new hosts », Biology Letters, vol. 11, , p. 20150233 (ISSN 1744-9561 et 1744-957X, DOI 10.1098/rsbl.2015.0233, lire en ligne, consulté le )
- Geographic variation in Varroa jacobsoni (Acari, Varroidae) : application of multivariate morphometric techniques, sur apidologie.org de novembre 1989, consulté le 5 juin 2017
- Charrière 2012
- Non trouvé le 5 juin 2017, sur daff.gov.au
- http://www.apiterra.fr/2010/07/le-conservatoire-de-labeille-noire-a-ouessant-35/
- http://nicolas.blogs.com/files/rapport_porciani_saeps_2012-analyse_de_risque.pdf
- « Varroa spread takes heavy toll », sur Stuff, (consulté le ).
- (en) « Beekeepers face new threat from varroa mite », sur The New Zealand Herald, (consulté le ).
- http://reunion-mayotte.cirad.fr/actualites/varroa_a_madagascar
- (en) Wolfgang Ritter, « Varroa mites and honey bee health: can Varroa explain part of the colony losses? / Apidologie », Apidologie, vol. 41, no 3, , p. 353–363 (ISSN 1297-9678, DOI 10.1051/apido/2010017, lire en ligne, consulté le ).
- (en) Francesco Nazzi, « How does the mite Varroa destructor kill the honeybee Apis mellifera? Alteration of cuticular hydrcarbons and water loss in infested honeybees - PubMed », Journal of insect physiology, vol. 58, no 12, , p. 1548–1555 (ISSN 1879-1611, PMID 23041382, DOI 10.1016/j.jinsphys.2012.09.008, lire en ligne, consulté le ).
- (en) D Cox-Foster, « Effects of parasitization by Varroa destructor on survivorship and physiological traits of Apis mellifera in correlation with viral incidence and microbial challenge - PubMed », Parasitology, vol. 134, no Pt 3, , p. 405–412 (ISSN 0031-1820, PMID 17078903, DOI 10.1017/S0031182006000710, lire en ligne, consulté le ).
- « Bee Mites Suppress Bee Immunity, Open Door For Viruses And Bacteria », sur ScienceDaily (consulté le ).
- (en) Diana L. Cox-Foster, « Impact of an ectoparasite on the immunity and pathology of an invertebrate: Evidence for host immunosuppression and viral amplification », Proceedings of the National Academy of Sciences, vol. 102, no 21, , p. 7470–7475 (ISSN 1091-6490, PMID 15897457, DOI 10.1073/pnas.0501860102, lire en ligne, consulté le ).
- « La Varroase des Abeilles », sur eap.mcgill.ca (consulté le ).
- A. SAKLI, « ESSAI PRÉLIMINAIRE DE TRAITEMENT DE LA VARROASE (à VARROA JACOBSONI) DE L'ABEILLE DOMESTIQUE PAR L'AMITRAZ / Apidologie », Apidologie, vol. 12, no 1, , p. 31–36 (ISSN 0044-8435, DOI doi:10.1051/apido:19810102, lire en ligne, consulté le ).
- (en) S. Di Pasquale, « Fluvalinate resistance of Varroa jacobsoni Oudemans (Acari: Varroidae) in Mediterranean apiaries of France / Apidologie », Apidologie, vol. 28, no 6, , p. 375–384 (ISSN 0044-8435, DOI doi:10.1051/apido:19970605, lire en ligne, consulté le ).
- (en) N. Milani, « The resistance of Varroa jacobsoni Oud to pyrethroids: a laboratory assay / Apidologie », Apidologie, vol. 26, no 5, , p. 415–429 (ISSN 0044-8435, DOI doi:10.1051/apido:19950507, lire en ligne, consulté le ).
- (en) Medwin H. Bew, « First report of Varroa destructor resistance to pyrethroids in the UK / Apidologie », Apidologie, vol. 33, no 4, , p. 357–366 (ISSN 1297-9678, DOI doi:10.1051/apido:2002027, lire en ligne, consulté le ).
- A scientific note on Varroa destructor resistance to coumaphos in the United States Jeff S. PETTIS USDA-ARS Bee Research Laboratory, Bldg 476 BARC-E, Beltsville, MD 20705, USA June 2003 http://hal.archives-ouvertes.fr/docs/00/89/18/66/PDF/hal-00891866.pdf
- Scala 2012
- Core 2012
- (en) Li Z, Chen Y, Zhang S, Chen S, Li W, Yan L, Shi L, Wu L, Sohr A, Su S, « Viral infection affects sucrose responsiveness and homing ability of forager honey bees, Apis mellifera L », PLoS One, vol. 8, no 10, , e77354. (PMID 24130876, PMCID PMC3795060, DOI 10.1371/journal.pone.0077354, lire en ligne [html])
- (en) Boncristiani HF, Evans JD, Chen Y, Pettis J, Murphy C, Lopez DL, Simone-Finstrom M, Strand M, Tarpy DR, Rueppell O, « In vitro infection of pupae with Israeli acute paralysis virus suggests disturbance of transcriptional homeostasis in honey bees (Apis mellifera) », PLoS One, vol. 8, no 9, , e73429. (PMID 24039938, PMCID PMC3764161, DOI 10.1371/journal.pone.0073429, lire en ligne [html])
- http://www.beeologics.com/wp-content/uploads/Reprint_from_Journal-Insect_Molecular_Biology.pdf
- (en) Di Prisco G, Pennacchio F, Caprio E, Boncristiani HF Jr, Evans JD, Chen Y, « Varroa destructor is an effective vector of Israeli acute paralysis virus in the honeybee, Apis mellifera », J Gen Virol, vol. 92, no Pt 1, , p. 151-5. (PMID 20926637, DOI 10.1099/vir.0.023853-0, lire en ligne [html])
- (en) Ai H1, Yan X, Han R, « Occurrence and prevalence of seven bee viruses in Apis mellifera and Apis cerana apiaries in China », J Invertebr Pathol, vol. 109, no 1, , p. 160-4. (PMID 22062807, DOI 10.1016/j.jip.2011.10.006)
- (en) Ribière M, Olivier V, Blanchard P, « Chronic bee paralysis: a disease and a virus like no other? », J Invertebr Pathol, vol. 103, no Suppl 1, , S120-31. (PMID 19909978, DOI 10.1016/j.jip.2009.06.013)
- (en) Dainat B, Neumann P, « Clinical signs of deformed wing virus infection are predictive markers for honey bee colony losses », J Invertebr Pathol, vol. 112, no 3, , p. 278-80. (PMID 23270875, DOI 10.1016/j.jip.2012.12.009)
- (en) Gauthier L, Ravallec M, Tournaire M, Cousserans F, Bergoin M, Dainat B, de Miranda JR, « Viruses associated with ovarian degeneration in Apis mellifera L. queens », PLoS One, vol. 6, no 1, , e16217. (PMID 21283547, DOI 10.1371/journal.pone.0016217, lire en ligne [html])
- (en) Desai SD, Eu YJ, Whyard S, Currie RW, « Reduction in deformed wing virus infection in larval and adult honey bees (Apis mellifera L.) by double-stranded RNA ingestion », Insect Mol Biol, vol. 21, no 4, , p. 446-55. (PMID 22690671, DOI 10.1111/j.1365-2583.2012.01150.x)
- (en) Martin SJ, Highfield AC, Brettell L, Villalobos EM, Budge GE, Powell M, Nikaido S, Schroeder DC, « Global honey bee viral landscape altered by a parasitic mite », Science, vol. 336, no 6086, , p. 1304-6. (PMID 22679096, DOI 10.1126/science.1220941)
- (en) Ingemar Fries, « Nosema ceranae has infected Apis mellifera in Europe since at least 1998 and may be more virulent than Nosema apis / Apidologie », Apidologie, vol. 38, no 6, , p. 558–565 (ISSN 1297-9678, DOI doi:10.1051/apido:2007037, lire en ligne, consulté le ).
- Robert A. Cramer Jr, « Iridovirus and Microsporidian Linked to Honey Bee Colony Decline », PLOS ONE, vol. 5, no 10, , e13181 (ISSN 1932-6203, DOI 10.1371/journal.pone.0013181, lire en ligne, consulté le ).
- http://myweb.dal.ca/gf331484/Williams_First%20detect.pdf
- « Quand un contrôleur sanitaire de la DDSV confond Nosema apis et Nosema ceranae - », sur Environnement.fr, (consulté le ).
- (en) « Nosema Ceranae : Kiss of Death or Much Ado about Nothing? - Scientific Beekeeping », sur Scientific Beekeeping, (consulté le ).
- (en) Aránzazu Meana, « Nosema ceranae, a new microsporidian parasite in honeybees in Europe - PubMed », Journal of invertebrate pathology, vol. 92, no 2, , p. 93–95 (ISSN 0022-2011, PMID 16574143, DOI 10.1016/j.jip.2006.02.005, lire en ligne, consulté le ).
- http://www.apidologie.org/articles/apido/pdf/2007/01/m6062.pdf
- Chong et Maugh 2007
- (en) Raquel Martín-Hernández, « Nosema ceranae (Microsporidia), a controversial 21st century honey bee pathogen - PubMed », Environmental microbiology reports, vol. 5, no 1, , p. 17–29 (ISSN 1758-2229, PMID 23757127, DOI 10.1111/1758-2229.12024, lire en ligne, consulté le ).
- (en) Marla Spivak, « Physiological and behavioral changes in honey bees (Apis mellifera) induced by Nosema ceranae infection - PubMed », PloS one, vol. 8, no 3, , e58165 (ISSN 1932-6203, PMID 23483987, DOI 10.1371/journal.pone.0058165, lire en ligne, consulté le ).
- (en) William O H Hughes, « Emerging dangers: deadly effects of an emergent parasite in a new pollinator host - PubMed », Journal of invertebrate pathology, vol. 114, no 2, , p. 114–119 (ISSN 1096-0805, PMID 23816821, DOI 10.1016/j.jip.2013.06.005, lire en ligne, consulté le ).
- (en) Yves Le Conte, « Flight behavior and pheromone changes associated to Nosema ceranae infection of honey bee workers (Apis mellifera) in field conditions - PubMed », Journal of invertebrate pathology, vol. 113, no 1, , p. 42–51 (ISSN 1096-0805, PMID 23352958, DOI 10.1016/j.jip.2013.01.002, lire en ligne, consulté le ).
- (en) Mariano Higes, « Nosema spp. infection and its negative effects on honey bees (Apis mellifera iberiensis) at the colony level - PubMed », Veterinary research, vol. 44, no 1, , p. 25 (ISSN 1297-9716, PMID 23574888, DOI 10.1186/1297-9716-44-25, lire en ligne, consulté le ).
- Daren M. Eiri, Guntima Suwannapong, Matthew Endler et James C. Nieh, « Nosema ceranae Can Infect Honey Bee Larvae and Reduces Subsequent Adult Longevity », PLoS ONE, vol. 10, , e0126330 (PMID 26018139, PMCID 4446295, DOI 10.1371/journal.pone.0126330, lire en ligne, consulté le )
- Mike Goblirsch, Zachary Y. Huang, Marla Spivak, « Physiological and Behavioral Changes in Honey Bees (Apis mellifera) Induced by Nosema ceranae Infection », PLoS ONE, vol. 8, no 3, (PMID 23483987, DOI 10.1371/journal.pone.0058165, lire en ligne, consulté le ).
- Higes 2009
- Brian S. Imai, « Nosema ceranae Escapes Fumagillin Control in Honey Bees », PLOS Pathogens, vol. 9, no 3, , e1003185 (ISSN 1553-7374, DOI 10.1371/journal.ppat.1003185, lire en ligne, consulté le ).
- Botías C, Martín-Hernández R, Meana A, Higes M. Screening alternative therapies to control Nosemosis type C in honey bee (Apis mellifera iberiensis) colonies. Res Vet Sci. 2013 Dec;95(3):1041-5. doi: 10.1016/j.rvsc.2013.09.012. Epub 2013 Oct 3. lien PubMed
- Vet Microbiol. 2013 Dec 27;167(3-4):474-83. doi: 10.1016/j.vetmic.2013.07.030. Epub 2013 Aug 9. Effects of the organic acids produced by a lactic acid bacterium in Apis mellifera colony development, Nosema ceranae control and fumagillin efficiency. Maggi M, Negri P, Plischuk S, Szawarski N, De Piano F, De Feudis L, Eguaras M, Audisio C.
- Holt HL, Aronstein KA, Grozinger CM. Chronic parasitization by Nosema microsporidia causes global expression changes in core nutritional, metabolic and behavioral pathways in honey bee workers (Apis mellifera). BMC Genomics, 2013 Nov 18;14:799. doi: 10.1186/1471-2164-14-799. article complet
- Jeroen van der Sluij (dir.), Noa Simon-Delso, Laura Maxim, Jean-Marc Bonmatin et Luc P Belzunces, « Neonicotinoids, bee disorders and the sustainability of pollinator service », Current Opinion in Environmental Sustainability, vol. 5, , p. 293–305
- Lori Lach, Madlen Kratz et Boris Baer, « Parasitized honey bees are less likely to forage and carry less pollen », Journal of Invertebrate Pathology, vol. 130, , p. 64-71 (DOI 10.1016/j.jip.2015.06.003, lire en ligne, consulté le )
- Gennaro Di Prisco, Xuan Zhang, Francesco Pennacchio et Emilio Caprio, « Dynamics of persistent and acute deformed wing virus infections in honey bees, Apis mellifera », Viruses, vol. 3, , p. 2425–2441 (ISSN 1999-4915, PMID 22355447, PMCID 3280512, DOI 10.3390/v3122425, lire en ligne, consulté le )
- Vincent Piou, Jérémy Tabart, Virginie Urrutia et Jean-Louis Hemptinne, « Impact of the Phoretic Phase on Reproduction and Damage Caused by Varroa destructor (Anderson and Trueman) to Its Host, the European Honey Bee (Apis mellifera L.) », PLOS ONE, vol. 11, , e0153482 (ISSN 1932-6203, PMID 27096154, PMCID 4838260, DOI 10.1371/journal.pone.0153482, lire en ligne, consulté le )
- Martin 2012
- Jennifer Welsh, « Mites and Virus Team Up to Wipe Out Beehives », sur Livescience.com, (consulté le ).
- (en) Yan Ping Chen, « Dynamics of persistent and acute deformed wing virus infections in honey bees, Apis mellifera - PubMed », Viruses, vol. 3, no 12, , p. 2425–2441 (ISSN 1999-4915, PMID 22355447, DOI 10.3390/v3122425, lire en ligne, consulté le ).
- (en) Peter Neumann, « Dead or alive: deformed wing virus and Varroa destructor reduce the life span of winter honeybees - PubMed », Applied and environmental microbiology, vol. 78, no 4, , p. 981–987 (ISSN 1098-5336, PMID 22179240, DOI 10.1128/AEM.06537-11, lire en ligne, consulté le ).
- Per Kryger, « Varroa-Virus Interaction in Collapsing Honey Bee Colonies », PLOS ONE, vol. 8, no 3, , e57540 (ISSN 1932-6203, DOI 10.1371/journal.pone.0057540, lire en ligne, consulté le ).
- (en) Mark L. Winston, « Honey bee colony mortality and productivity with single and dual infestations of parasitic mite species / Apidologie », Apidologie, vol. 32, no 6, , p. 567–575 (ISSN 1297-9678, DOI doi:10.1051/apido:2001144, lire en ligne, consulté le ).
- (en) L. Wilfert, G. Long, H. C. Leggett et P. Schmid-Hempel, « Deformed wing virus is a recent global epidemic in honeybees driven by Varroa mites », Science, vol. 351, , p. 594–597 (ISSN 0036-8075 et 1095-9203, DOI 10.1126/science.aac9976, lire en ligne, consulté le )
- (en) F Mutinelli, « Sudden deaths and colony population decline in Greek honey bee colonies - PubMed », Journal of invertebrate pathology, vol. 105, no 3, , p. 335–340 (ISSN 1096-0805, PMID 20804765, DOI 10.1016/j.jip.2010.08.004, lire en ligne, consulté le ).
- (en) Peter Neumann, « Negative correlation between Nosema ceranae spore loads and deformed wing virus infection levels in adult honey bee workers - PubMed », Journal of invertebrate pathology, vol. 108, no 3, , p. 224–225 (ISSN 1096-0805, PMID 21939664, DOI 10.1016/j.jip.2011.08.012, lire en ligne, consulté le ).
- R. Scott Cornman, David R. Tarpy et al., « Pathogen Webs in Collapsing Honey Bee Colonies », PLoS ONE, vol. 7, no 8, (PMID 22927991, DOI 10.1371/journal.pone.0043562, lire en ligne, consulté le ).
- (en) Crop Pollination Exposes Honey Bees to Pesticides Which Alters Their Susceptibility to the Gut Pathogen Nosema ceranae, Pettis JS, Lichtenberg EM, Andree M, Stitzinger J, Rose R, et al., 2013
- (en) Aleš Gregorc, « Chronic bee paralysis virus and Nosema ceranae experimental co-infection of winter honey bee workers (Apis mellifera L.) - PubMed », Viruses, vol. 5, no 9, , p. 2282–2297 (ISSN 1999-4915, PMID 24056674, DOI 10.3390/v5092282, lire en ligne, consulté le ).
- Rassol Bahreini et Robert W. Currie, « The influence of Nosema (Microspora: Nosematidae) infection on honey bee (Hymenoptera: Apidae) defense against Varroa destructor (Mesostigmata: Varroidae) », Journal of Invertebrate Pathology, vol. 132, , p. 57-65 (ISSN 1096-0805, PMID 26283465, DOI 10.1016/j.jip.2015.07.019, lire en ligne, consulté le )
- Kuchinskas 2007
- Pierre-Yves Laffont, Transhumance et estivage en occident des origines aux enjeux actuels, , 415 p. (ISBN 978-2-85816-843-9, lire en ligne), p. 263.
- « UNE AUTRE RESSOURCE POUR L'APICULTEUR : L'AIDE A LA POLLINISATION », sur apicolturaonline.it (consulté le ).
- Hailey N. Scofield et Heather R. Mattila, « Honey Bee Workers That Are Pollen Stressed as Larvae Become Poor Foragers and Waggle Dancers as Adults », PLoS ONE, vol. 10, , e0121731 (PMID 25853902, PMCID 4390236, DOI 10.1371/journal.pone.0121731, lire en ligne, consulté le )
- Adam G. Dolezal, Jimena Carrillo-Tripp, W. Allen Miller et Bryony C. Bonning, « Intensively Cultivated Landscape and Varroa Mite Infestation Are Associated with Reduced Honey Bee Nutritional State », PLOS ONE, vol. 11, , e0153531 (ISSN 1932-6203, PMID 27070422, PMCID 4829173, DOI 10.1371/journal.pone.0153531, lire en ligne, consulté le )
- Réseau Biodiversité pour les Abeilles
- « Le déclin des papillons démontre celui de la biodiversité », sur Futura (consulté le ).
- Tasei et Aupinel 2008
- Alaux et al. 2010
- Lee, Simpson et Wilson 2008
- Rinderer et Rothenbuhler 1974
- Li 2007
- « Les jachères au secours des abeilles » http://www.actu-environnement.com/ae/news/2042.php4
- (en) « Sowing field margins with a pollen and nectar seed mix enhances the density and diversity of bumblebees Bombus spp., on arable farms in England » : « citant (en) Pywell R.F., Warman E.A., Hulmes L., Hulmes S., Nuttall P., Sparks T.H., Critchley C.N.R. & Sherwood A. (2006) « Effectiveness of new agri-environment schemes in providing foraging resources for bumblebees in intensively farmed landscapes » Biological Conservation, 129, 192-206 »
- (en) « Bumblebees Bombus spp. use forage plants sown on field margins as a pollen source, at Manor Farm, Eddletorpe, North Yorkshire, England » : « citant (en) Carvell C., Westrich P., Meek W.R., Pywell R.F. & Nowakowski M. (2006) « Assessing the value of annual and perennial forage mixtures for bumblebees by direct observation and pollen analysis » Apidologie, 37, 326-340. »
- (en) « Establishment of clover-rich field margins as a forage resource for bumblebees Bombus spp. on Romney Marsh, Kent, England » : « citant (en) Gardiner T., Edwards M. & Hill J. (2008) « Establishment of clover-rich field margins as a forage resource for bumblebees Bombus spp. on Romney Marsh, Kent, England » Conservation Evidence, 5, 51-57. »
- (en) « Improving the fortunes of pollinators, whilst retaining farm profitability »
- (en) Buri P, Humbert JY, Arlettaz R, « Promoting pollinating insects in intensive agricultural matrices: field-scale experimental manipulation of hay-meadow mowing regimes and its effects on bees », PLoS One, vol. 9, no 1, , e85635. (PMID 24416434, PMCID PMC3887108, DOI 10.1371/journal.pone.0085635, lire en ligne [html])
- (en) Thomas J. Wood, John M. Holland, William O. H. Hughes et Dave Goulson, « Targeted agri-environment schemes significantly improve the population size of common farmland bumblebee species », Molecular Ecology, vol. 24, , p. 1668-1680 (ISSN 1365-294X, DOI 10.1111/mec.13144, lire en ligne, consulté le )
- « Apiservices », sur apiculture.com (consulté le ).
- « Le nourrissement / L'Abeille du Forez », sur zoo-logique.org (consulté le ).
- Daniel R. Schmehl, Peter E. A. Teal, James L. Frazier et Christina M. Grozinger, « Genomic analysis of the interaction between pesticide exposure and nutrition in honey bees (Apis mellifera) », Journal of Insect Physiology, vol. 71, , p. 177-190 (DOI 10.1016/j.jinsphys.2014.10.002, lire en ligne, consulté le )
- Bob Yirka, « Researchers find high-fructose corn syrup may be tied to worldwide collapse of bee colonies », sur Phys.org, (consulté le ).
- Science X staff, « Heat forms potentially harmful substance in high-fructose corn syrup », sur Phys.org, (consulté le ).
- « HostMetro.com », sur apiculture-populaire.com (consulté le ).
- http://www.gembloux.ulg.ac.be/entomologie-fonctionnelle-et-evolutive/enseignement/tfe/hmf/
- http://www.uwplatt.edu/~cornettc/Ag%20and%20Food%20Publish.pdf
- (en) Claude Saegerman, « Hydroxymethylfurfural: a possible emergent cause of honey bee mortality? - PubMed », Journal of agricultural and food chemistry, vol. 61, no 49, , p. 11865–11870 (ISSN 1520-5118, PMID 24127696, DOI 10.1021/jf403280n, lire en ligne, consulté le ).
- (en) May R. Berenbaum, « Honey constituents up-regulate detoxification and immunity genes in the western honey bee Apis mellifera », Proceedings of the National Academy of Sciences, vol. 110, no 22, , p. 8842–8846 (ISSN 1091-6490, PMID 23630255, DOI 10.1073/pnas.1303884110, lire en ligne, consulté le ).
- Sophie Krainer, Robert Brodschneider, Jutta Vollmann et Karl Crailsheim, « Effect of hydroxymethylfurfural (HMF) on mortality of artificially reared honey bee larvae (Apis mellifera carnica) », Ecotoxicology (London, England), (ISSN 1573-3017, PMID 26590927, DOI 10.1007/s10646-015-1590-x, lire en ligne, consulté le )
- http://www.paysan-breton.fr/article/12122/l%92insemination-artificielle,-pour-les-abeilles-aussi.html
- http://www.agefi.com/une/detail/archive/2012/october/artikel/les-vrais-effets-de-la-mortalite-des-abeilles.html
- Maxime Lambert, « Abeilles : plus la reine s'accouple, plus la colonie a des chances de survivre », sur Maxisciences.com, (consulté le ).
- « Le CIRAD se penche sur la carte génétique des abeilles de la Réunion », sur Zinfos 974, l'actualité de l'île de La Réunion (consulté le ).
- « Connexion », sur apiculture-france.com (consulté le ).
- http://www.pollinis.org/spip.php?page=article&id_article=82
- « Apiservices », sur beekeeping.com (consulté le ).
- « Le renouvellement des reines / L'Abeille du Forez », sur zoo-logique.org (consulté le ).
- Hernández López J1, Schuehly W, Crailsheim K, Riessberger-Gallé U. Trans-generational immune priming in honeybees. Proc Biol Sci. 2014 Apr 30;281(1785):20140454. doi: 10.1098/rspb.2014.0454. Print 2014. http://rspb.royalsocietypublishing.org/content/281/1785/20140454.long
- Locke B, Forsgren E, de Miranda JR (2014) Increased Tolerance and Resistance to Virus Infections: A Possible Factor in the Survival of Varroa destructor-Resistant Honey Bees (Apis mellifera). PLoS ONE 9(6): e99998. doi:10.1371/journal.pone.0099998 http://www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0099998
- Jeffery S. Pettis, Nathan Rice, Katie Joselow et Dennis vanEngelsdorp, « Colony Failure Linked to Low Sperm Viability in Honey Bee (Apis mellifera) Queens and an Exploration of Potential Causative Factors », PloS One, vol. 11, , e0147220 (ISSN 1932-6203, PMID 26863438, PMCID 4749221, DOI 10.1371/journal.pone.0147220, lire en ligne, consulté le )
- (en) Desneux et al., « The sublethal effects of pesticides on beneficial arthropods », Annual Review of Entomology,
- (en) Rondeau G et al., « Delayed and time-cumulative toxicity of imidacloprid in bees, ants and termites », Scientific Reports,
- Haubruge 2006
- (en) G. Christopher Cutler and Cynthia D. Scott-Dupree « Exposure to Clothianidin Seed-Treated Canola Has No Long-Term Impact on Honey Bees » Journal of Economic Entomology, 100(3):765-772. 2007. Published By: Entomological Society of America DOI: 10.1603/0022-0493(2007)100[765:ETCSCH]2.0.CO;2 URL:http://www.bioone.org/doi/full/10.1603/0022- 0493%282007%29100%5B765%3AETCSCH%5D2.0.CO%3B2
- E. Haubruge, « Does Imidacloprid Seed-Treated Maize Have an Impact on Honey Bee Mortality? », Journal of Economic Entomology, vol. 102, no 2, , p. 616–623 (ISSN 1938-291X, DOI 10.1603/029.102.0220, lire en ligne, consulté le ).
- (en) Alaux C, Brunet JL, Dussaubat C, Mondet F, Tchamitchan S, Cousin M, Brillard J, Baldy A, Belzunces LP, Le Conte Y, « Interactions between Nosema microspores and a neonicotinoid weaken honeybees (Apis mellifera) », Environ Microbiol, vol. 12, no 3, , p. 774-82. (PMID 20050872, PMCID PMC2847190, DOI 10.1111/j.1462-2920.2009.02123.x, lire en ligne [html])
- (en) Aufauvre J, Misme-Aucouturier B, Viguès B, Texier C, Delbac F. & Blot N. « Transcriptome Analyses of the Honeybee Response to Nosema ceranae and Insecticides » PloS one 2014;9(3):e91686. DOI:10.1371/journal.pone.0091686
- Whitehorn 2012
- Henry 2012
- Stokstad 2012
- Cresswell 2012
- (en) Guez D, « A common pesticide decreases foraging success and survival in honey bees: questioning the ecological relevance », Front Physiol, no 4, , p. 37. (PMID 23472059, PMCID PMC3590638, DOI 10.3389/fphys.2013.00037, lire en ligne [html])
- (en) Guez D, « Henry et al. (2012) homing failure formula, assumptions, and basic mathematics: a comment », Front Physiol, no 4, , p. 142. (PMID 23801964, PMCID PMC3687715, DOI 10.3389/fphys.2013.00142, lire en ligne [html])
- Autorité européenne de sécurité des aliments 2012
- Agence nationale de sécurité sanitaire de l'alimentation, de l'environnement et du travail 2012
- (en) Schneider CW, Tautz J, Grünewald B, Fuchs S., « RFID tracking of sublethal effects of two neonicotinoid insecticides on the foraging behavior of Apis mellifera », PLoS One, vol. 7, no 1, , e30023 (PMID 22253863, DOI 10.1371/journal.pone.0030023, lire en ligne [html])
- (en) Henry M, « Assessing homing failure in honeybees exposed to pesticides: Guez's (2013) criticism illustrates pitfalls and challenges », Front Physiol, no 4, , p. 352. (PMID 24348427, PMCID PMC3845207, DOI 10.3389/fphys.2013.00352, lire en ligne [html])
- INRA 2012
- (en) Cresswell JE, Desneux N, vanEngelsdorp D. (2012) « Dietary traces of neonicotinoid pesticides as a cause of population declines in honey bees: an evaluation by Hill's epidemiological criteria » Pest Manag Sci. ;68(6):819-27. DOI:10.1002/ps.3290 PMID
- (en) Blacquière T, Smagghe G, van Gestel CA, Mommaerts V., « Neonicotinoids in bees: a review on concentrations, side-effects and risk assessment », Ecotoxicology, vol. 21, no 4, , p. 973-92. (PMID 22350105, PMCID PMC3338325, DOI 10.1007/s10646-012-0863-x, lire en ligne [html])
- Département de l'Environnement, de l'Alimentation et des Affaires rurales 2013
- (en) Edward Pilling, Peter Campbell, A Four-Year Field Program Investigating Long-Term Effects of Repeated Exposure of Honey Bee Colonies to Flowering Crops Treated with Thiamethoxam, Syngenta, 23 octobre 2013
- DeepDyve, Inc., « DeepDyve - Instant access to the journals you need. », sur DeepDyve (consulté le ).
- (en) Laycock I, Cresswell JE, « Repression and recuperation of brood production in Bombus terrestris bumble bees exposed to a pulse of the neonicotinoid pesticide imidacloprid », PLoS One, vol. 8, no 11, , e79872. (PMID 24224015, PMCID PMC3817108, DOI 10.1371/journal.pone.0079872, lire en ligne [html])
- https://biblio.ugent.be/input/download?func=downloadFile&recordOId=2027235&fileOId=2027245
- « Abeilles et pesticides : quelques rappels à l’histoire », sur Blog d'Alain Godard, (consulté le )
- Jean Louveaux, Pollinisation et productions végétales, Editions Quae, , p. 565-575
- https://www.lesechos.fr/20/08/2007/LesEchos/19985-63-ECH_la-mort-des-abeilles-met-la-planete-en-danger.htm
- Dennis vanEngelsdorp & al. (2014) Preliminary Results: Honey Bee Colony Losses in the United States, 2013-2014 ; 6 mai 2014 (rapport préliminaire)
- (en) Johannes Fischer, Teresa Müller, Anne-Kathrin Spatz, Uwe Greggers, Bernd Grünewald, Randolf Menzel « Neonicotinoids Interfere with Specific Components of Navigation in Honeybees » PloS one 2014;9(3):e91364. DOI:10.1371/journal.pone.0091364
- « Bee Experts Dismantle Touted 'Harvard' Neonics-Colony Collapse Disorder Study As 'Activist Science' », sur The Huffington Post (consulté le )
- « Bad Science Doesn't Help Bees », sur BloombergView, (consulté le )
- Galen P. Dively, Michael S. Embrey, Alaa Kamel et David J. Hawthorne, « Assessment of Chronic Sublethal Effects of Imidacloprid on Honey Bee Colony Health », PLoS ONE, vol. 10, , e0118748 (PMID 25786127, PMCID 4364903, DOI 10.1371/journal.pone.0118748, lire en ligne, consulté le )
- Gina Retschnig, Geoffrey R. Williams, Richard Odemer et Janina Boltin, « Effects, but no interactions, of ubiquitous pesticide and parasite stressors on honey bee (Apis mellifera) lifespan and behaviour in a colony environment », Environmental Microbiology, (ISSN 1462-2920, PMID 25728008, DOI 10.1111/1462-2920.12825, lire en ligne, consulté le )
- Gegear, R. J., Manson, J. S., & Thomson, J. D. (2007). Ecological context influences pollinator deterrence by alkaloids in floral nectar. Ecology Letters, 10(5), 375-382 ([résumé]).
- Natarajan Singaravelan, Gidi Nee'man, Moshe Inbar, Ido Izhaki (2005), Feeding Responses of Free-flying Honeybees to Secondary Compounds Mimicking Floral Nectars Journal of Chemical Ecology, December 2005, Volume 31, Issue 12, p. 2791-2804, 18 décembre 2005 (résumé)
- (en) Leif L. Richardson, Lynn S. Adler, Anne S. Leonard et Jonathan Andicoechea, « Secondary metabolites in floral nectar reduce parasite infections in bumblebees », Proceedings of the Royal Society of London B: Biological Sciences, vol. 282, , p. 20142471 (ISSN 0962-8452 et 1471-2954, PMID 25694627, DOI 10.1098/rspb.2014.2471, lire en ligne, consulté le )
- R. Charvet, M. Katouzian-Safadi, M.-E. Colin et P.-A. Marchand, « [Systemic insecticides: new risk for pollinator insects] », Annales Pharmaceutiques Françaises, vol. 62, , p. 29-35 (ISSN 0003-4509, PMID 14747770, lire en ligne, consulté le )
- (en) Sébastien C. Kessler, Erin Jo Tiedeken, Kerry L. Simcock et Sophie Derveau, « Bees prefer foods containing neonicotinoid pesticides », Nature, vol. 521, , p. 74-76 (ISSN 0028-0836, DOI 10.1038/nature14414, lire en ligne, consulté le )
- Rachel Feltman (2015), article intitulé Speaking of Science New studies find that bees actually want to eat the pesticides that hurt them publié par le Washington Post le 22 avril 2015
- L'Arboriculture fruitière, no 578 à 584, Société pomologique de France, 2004, p. 14
- http://www.adapi.itsap.asso.fr/infos/22222222222222222.php
- Ahmed AA1, Abd-Elhady2: Efficacy of two fungus-based biopesticide against the honeybee ectoparasitic mite, Varroa destructor. 1Department of Economic Entomology and Agricultural Zoology, Faculty of Agriculture, Menoufiya University, Egypt. 2Departmnent of Pesticides, Faculty of Agriculture, Menoufiya University, Shebin El-Kom 32511 Egypt. HK2.Pak J Biol Sci. 2013 Aug 15;16(16):819-25. http://scialert.net/fulltext/?doi=pjbs.2013.819.825
- Popova M, Reyes M, Le Conte Y, Bankova V: Propolis chemical composition and honeybee resistance against Varroa destructor. Nat Prod Res. 2014 Jan 31. Institute of Organic Chemistry with Centre of Phytochemistry, Bulgarian Academy of Sciences, Sofia, Bulgaria. https://www.ncbi.nlm.nih.gov/pubmed/24483289
- Nicodemo D, De Jong D, Couto RH, Malheiros EB: Honey bee lines selected for high propolis production also have superior hygienic behavior and increased honey and pollen stores. Genet Mol Res. 2013 Dec 19;12(4):6931-8. doi: 10.4238/2013.December.19.12. Curso de Zootecnia, Universidade Estadual Paulista, Dracena, SP, Brasil nicodemo@dracena.unesp.br. https://www.ncbi.nlm.nih.gov/pubmed/24391041
- (en) Michel Treilhou, « Thymol as an alternative to pesticides: persistence and effects of Apilife Var on the phototactic behavior of the honeybee Apis mellifera - PubMed », Environmental science and pollution research international, vol. 21, no 7, , p. 4934–4939 (ISSN 1614-7499, PMID 24043507, DOI 10.1007/s11356-013-2143-6, lire en ligne, consulté le ).
- Jennifer A. Berry, W. Michael Hood et al., « Field-Level Sublethal Effects of Approved Bee Hive Chemicals on Honey Bees (Apis mellifera L) », PLoS ONE, vol. 8, no 10, (PMID 24204638, DOI 10.1371/journal.pone.0076536, lire en ligne, consulté le ).
- Bernard NICOLLET - Tous droits réservés - téléchargement non autorisé, « Traitement Acide Formique contre Varroa des abeilles », sur abeille-et-nature.com (consulté le ).
- Walter S. Sheppard, « Sub-Lethal Effects of Pesticide Residues in Brood Comb on Worker Honey Bee (Apis mellifera) Development and Longevity », PLOS ONE, vol. 6, no 2, , e14720 (ISSN 1932-6203, DOI 10.1371/journal.pone.0014720, lire en ligne, consulté le ).
- Ellis 1997
- (en) Neil Kirk Hillier, « Effects of fluvalinate on honey bee learning, memory, responsiveness to sucrose, and survival - PubMed », The Journal of experimental biology, vol. 216, no Pt 15, , p. 2931–2938 (ISSN 1477-9145, PMID 23619403, DOI 10.1242/jeb.086538, lire en ligne, consulté le ).
- Chrisovalantis Papaefthimiou, Alexandros Papachristoforou, George Theophilidis, Biphasic responses of the honeybee heart to nanomolar concentrations of amitraz, Pesticide Biochemistry and Physiology, Volume 107, Issue 1, September 2013, Pages 132–137, doi: https://dx.doi.org/10.1016/j.pestbp.2013.06.005, url : http://www.sciencedirect.com/science/article/pii/S0048357513001120
- Xenobiotic Effects on Intestinal Stem Cell Proliferation in Adult Honey Bee (Apis mellifera L) Workers Cordelia Forkpah, 1 Luke R. Dixon, 1 Susan E. Fahrbach, 2 and Olav Rueppell1,* Nicolas Desneux, Editor PLoS One. 2014; 9(3): e91180. Published online Mar 7, 2014. doi: 10.1371/journal.pone.0091180
- (en) Reed M. Johnson et al., « Pesticides and honey bee toxicity – USA », Apidologie, vol. 41, , p. 312-331 (DOI https://dx.doi.org/10.1051/apido/2010018, lire en ligne)
- (en) Bethany S. Teeters et al., « Using video-tracking to assess sublethal effects of pesticides on honey bees », Environmental Toxicology and Chemistry, The Society of Environmental Toxicology and Chemistry, vol. 31, , p. 1349–1354 (DOI 10.1002/etc.1830, lire en ligne)
- Nathan Tene et al., « Recherche de résidus de pesticides dans la cire d’abeille : comparaison entre les cires de corps et d’opercules. », non publié, Université de Toulouse, Centre Universitaire Champollion, équipe Chimie et Biochimie des Interactions, (lire en ligne [PDF])
- (en) David J. Hawthorne, « Killing Them with Kindness? In-Hive Medications May Inhibit Xenobiotic Efflux Transporters and Endanger Honey Bees », PLoS ONE, Université de Gand, (DOI 10.1371/journal.pone.0026796, lire en ligne)
- (en) Sally M. Williamson et al., « Acute exposure to a sublethal dose of imidacloprid and coumaphos enhances olfactory learning and memory in the honeybee Apis mellifera », Invertebrate Neuroscience, Springer, (DOI 10.1007/s10158-012-0144-7, lire en ligne)
- Wanyi Zhu, Daniel R. Schmehl, Christopher A. Mullin et James L. Frazier, « Four common pesticides, their mixtures and a formulation solvent in the hive environment have high oral toxicity to honey bee larvae », PloS One, vol. 9, no 1, , e77547 (ISSN 1932-6203, PMID 24416121, PMCID PMC3885384, DOI 10.1371/journal.pone.0077547, lire en ligne, consulté le )
- « 220152335 / ephy », sur anses.fr (consulté le ).
- « Chloropyrifos-Éthyl », sur e-phy.agriculture.gouv.fr (consulté le )
- (en) « SAGE Journals : Your gateway to world-class research journals », sur SAGE Journals (consulté le ).
- James L. Frazier, « Four Common Pesticides, Their Mixtures and a Formulation Solvent in the Hive Environment Have High Oral Toxicity to Honey Bee Larvae », PLOS ONE, vol. 9, no 1, , e77547 (ISSN 1932-6203, DOI 10.1371/journal.pone.0077547, lire en ligne, consulté le ).
- http://onlinelibrary.wiley.com/doi/10.1002/ejlt.200500284/abstract
- (en) G C Diamantidis, « Determination of fluvalinate residues in beeswax by gas chromatography with electron-capture detection - PubMed », Journal of AOAC International, vol. 83, no 5, , p. 1225–1228 (ISSN 1060-3271, PMID 11048864, lire en ligne, consulté le ).
- (en) Antonio Herrera, « Impact of the use of fluvalinate on different types of beeswax from Spanish hives - PubMed », Archives of environmental contamination and toxicology, vol. 58, no 3, , p. 733–739 (ISSN 1432-0703, PMID 19756841, DOI 10.1007/s00244-009-9387-7, lire en ligne, consulté le ).
- http://www.californiabeecompany.com/articles/acaricides_e-1.pdf
- http://same-apiculture.colinweb.fr/Le-declin-des-abeilles
- Cummins 2007
- Are global honey bee declines caused by diesel pollution? Date: November 2, 2011 Source: University of Southampton Summary: Scientists are investigating a possible link between tiny particles of pollution found in diesel fumes and the global collapse of honey bee colonies. http://www.sciencedaily.com/releases/2011/10/111007073153.htm
- Elsa Youngsteadt, R. Holden Appler, Margarita M. López-Uribe et David R. Tarpy, « Urbanization Increases Pathogen Pressure on Feral and Managed Honey Bees », PloS One, vol. 10, , e0142031 (ISSN 1932-6203, PMID 26536606, DOI 10.1371/journal.pone.0142031, lire en ligne, consulté le )
- Mackintosh 2007
- Latsch 2007
- Kaatz 2005
- Christoph C. Tebbe, « Effect of Stacked Insecticidal Cry Proteins from Maize Pollen on Nurse Bees (Apis mellifera carnica) and Their Gut Bacteria », PLOS ONE, vol. 8, no 3, , e59589 (ISSN 1932-6203, DOI 10.1371/journal.pone.0059589, lire en ligne, consulté le ).
- http://www.annualreviews.org/doi/pdf/10.1146/annurev.arplant.043008.092013
- Rose, Dively et Pettis 2007
- Marvier et al. 2007
- (en) Jian Duan (dir.) et Joseph Huesing, « A Meta-Analysis of Effects of Bt Crops on Honey Bees (Hymenoptera: Apidae) », PLOS ONE, (DOI 10.1371/journal.pone.0001415, lire en ligne)
- Baal et al. 1991
- La Voix du Nord 2012
- www.contaminations-chimiques.info 2007
- auteur1=Dominique Raizon |titre=L'Abeille Noire d’Ouessant est en pleine forme |éditeur=rfi.fr|volume= |numéro= |jour=5 |mois=avril |année=2012 |pages= |issn= |lire en ligne=http://www.rfi.fr/science/20120404-abeille-noire-ouessant-est-pleine-forme
- de biosphère îles et de la mer d'Iroise
- Rouzic, Samuel Perichon Le; Peña et al., « L’élevage des abeilles mélipones sur l’île de Cuba : une enquête et... », Études caribéennes, Université des Antilles, nos 27-28, (ISSN 1779-0980, lire en ligne, consulté le ).
- http://www.larecherche.fr/savoirs/environnement/meme-abeilles-sauvages-declinent-01-12-2009-87693
- (en) Erik Stokstad, « Pollinator Diversity Declining in Europe », Science, no 5785, , p. 286 (DOI 10.1126/science.313.5785.286a)
- Mass-flowering crops enhance wild bee abundance. Oecologia. 2013 Jun;172(2):477-84. doi: 10.1007/s00442-012-2515-5. Epub 2012 Nov 1. Holzschuh A1, Dormann CF, Tscharntke T, Steffan-Dewenter I. 1Department of Animal Ecology and Tropical Biology, Biocenter, University of Würzburg, Am Hubland, 97074, Würzburg, Germany. andrea.holzschuh@uni-wuerzburg.de https://www.ncbi.nlm.nih.gov/pubmed/23114428
- A global quantitative synthesis of local and landscape effects on wild bee pollinators in agroecosystems. Kennedy CM1, Lonsdorf E, Neel MC, Williams NM, Ricketts TH, Winfree R, Bommarco R, Brittain C, Burley AL, Cariveau D, Carvalheiro LG, Chacoff NP, Cunningham SA, Danforth BN, Dudenhöffer JH, Elle E, Gaines HR, Garibaldi LA, Gratton C, Holzschuh A, Isaacs R, Javorek SK, Jha S, Klein AM, Krewenka K, Mandelik Y, Mayfield MM, Morandin L, Neame LA, Otieno M, Park M, Potts SG, Rundlöf M, Saez A, Steffan-Dewenter I, Taki H, Viana BF, Westphal C, Wilson JK, Greenleaf SS, Kremen C. Ecol Lett. 2013 May;16(5):584-99. doi: 10.1111/ele.12082. Epub 2013 Mar 11. 1Urban Wildlife Institute, Lincoln Park Zoo, Chicago, IL 60614, USA. ckennedy@tnc.org https://www.ncbi.nlm.nih.gov/pubmed/23489285
- Emerging dangers: deadly effects of an emergent parasite in a new pollinator host. Graystock P1, Yates K, Darvill B, Goulson D, Hughes WO. J Invertebr Pathol. 2013 Oct;114(2):114-9. doi: 10.1016/j.jip.2013.06.005. https://www.ncbi.nlm.nih.gov/pubmed/23816821
- Disease associations between honeybees and bumblebees as a threat to wild pollinators M. A. Fürst, D. P. McMahon, J. L. Osborne, R. J. Paxton & M. J. F. Brown Nature 506, 364–366 doi:10.1038/nature12977http://www.nature.com/nature/journal/v506/n7488/full/nature12977.html
- John Bryden, « Chronic sublethal stress causes bee colony failure », Ecology Letters, vol. 16, , p. 1463–1469 (DOI 10.1111/ele.12188)
- Stéphane Foucart, « Pesticides : les mathématiques au secours des abeilles », Le Monde, (lire en ligne)
- (en) Jeremy T. Kerr, Alana Pindar, Paul Galpern et Laurence Packer, « Climate change impacts on bumblebees converge across continents », Science, vol. 349, , p. 177-180 (ISSN 0036-8075 et 1095-9203, PMID 26160945, DOI 10.1126/science.aaa7031, lire en ligne, consulté le )
- (en) David Kleijn, Rachael Winfree, Ignasi Bartomeus et Luísa G. Carvalheiro, « Delivery of crop pollination services is an insufficient argument for wild pollinator conservation », Nature Communications, vol. 6, (PMID 26079893, PMCID 4490361, DOI 10.1038/ncomms8414, lire en ligne, consulté le )
- (en) BEEPOP: A honeybee population dynamics simulation model, sur le site sciencedirect.com, consulté le 17 novembre 2013
- (en) Khoury DS, Barron AB, Myerscough MR, « Modelling food and population dynamics in honey bee colonies », PLoS One, vol. 8, no 5, , e59084. (PMID 23667418, PMCID PMC3647073, DOI 10.1371/journal.pone.0059084, lire en ligne [html])
- [PDF] A review of honeybee models and a short introduction to the new integrated colony model BEEHAVE, sur le site europa.eu
- (en) Becher MA, Osborne JL, Thorbek P, Kennedy PJ, Grimm V, « Towards a systems approach for understanding honeybee decline: a stocktaking and synthesis of existing models », J Appl Ecol, vol. 50, no 4, , p. 868-880. (PMID 24223431, PMCID PMC3810709, DOI 10.1111/1365-2664.12112, lire en ligne [html])
- (en) Volker Grimm, Matthias Becher, Peter Kennedy, Juliane Horn, Pernille Thorbek, Juliet Osborne, « The new integrated model BEEHAVE: a tool for exploring multiple stressors of honeybee colonies » [PDF], sur sesss08.setac.eu
Voir aussi
Articles connexes
- Apiologie (en), branche correspondante de l'entomologie.
- Apiculture
- Abeille et Abeille à miel
- Gaucho (insecticide), Régent TS et Varroa
- Syngenta
- Extinction des espèces
- Biodiversité
- Monoculture
- Écotoxicologie
- Fragmentation (écologie)
- Maladie noire
- Pollinisation
- Pollinisateur
- Intoxication des abeilles
- Maladies de l'abeille domestique
- Vanishing of the Bees (film documentaire)
- Situation au Royaume-Uni (en)
Bibliographie
Par ordre chronologique de publication.
Publications scientifiques
- (en) Thomas E. Rinderer et Walter C. Rothenbuhler, « The influence of pollen on the susceptibility of honey-bee larvae to Bacillus larvae », Journal of Invertebrate Pathology, Elsevier, vol. 23, no 3, , p. 347-350 (DOI 10.1016/0022-2011(74)90100-1, lire en ligne, consulté le )
- T. Baal et al., « Groupe de travail des Instituts de recherche apicole d'Allemagne. Réunion de Celle du 26-28 mars 1991 : Facteurs physiologiques, éthologiques et écologiques concernant la mortalité d'insectes sous les tilleurs à floraison tardive », Apidologie, EDP Sciences, vol. 22, no 4, , p. 431-432 (ISSN 0044-8435 et 1297-9678, DOI 10.1051/apido:19910407, résumé, lire en ligne)
- (en) M. D. Ellis, « The effect of Apistan® on honey bee (Apis mellifera L). Responses to methyl parathion, carbaryl and bifenthrin exposure », Apidologie, EDP Sciences, vol. 28, , p. 123-127 (ISSN 0044-8435, DOI 10.1051/apido:19970303, résumé, lire en ligne, consulté le )
- (de) Hans-Hinrich Kaatz, Auswirkungen von BT-Maispollen auf die Honigbiene : Methodenentwicklung zu Wirkungsprüfung und Monitoring: Verbundprojekt: "Sicherheitsforschung und Monitoring-Methoden zum Anbau von Bt-Mais" ; Schlußbericht 2004 ; Berichtszeitraum vom 30.6.2001 - 31.12.2004, (OCLC 255710178)
- (en) J. D. Evans, « Immune pathways and defence mechanisms in honey bees Apis mellifera », Insect Molecular Biology, Royal Entomological Society, vol. 15, no 5, , p. 645-656 (ISSN 1365-2583, DOI 10.1111/j.1365-2583.2006.00682.x, lire en ligne)
- Eric Haubruge et al., « Le dépérissement de l'abeille domestique, Apis mellifera L., 1758 (Hymenoptera : Apidae) : faits et causes probables », Notes fauniques de Gembloux, Faculté des sciences agronomiques de Gembloux, vol. 59, no 1, , p. 3-21 (ISSN 0770-2019, lire en ligne, consulté le )
- (en) (en) M. Frazier, D. vanEngelsdor, D. Caron, « FAQ's - Colony Collapse Disorder », Mid-Atlantic Apiculture Research and Extension Consortium, (consulté le )
- (en) Joe Cummins, « Parasitic Fungi and Pesticides Act Synergistically to Kill Honeybees? », Institute of Science in Society, (consulté le )
- (en) Michelle Marvier, Chanel McCreedy, James Regetz et Peter Kareiva, « A Meta-Analysis of Effects of Bt Cotton and Maize on Nontarget Invertebrates », Science, vol. 316, no 5830, , p. 1475-1477 (ISSN 0036-8075, DOI 10.1126/science.1139208, résumé)
- (en) Renée Johnson, Recent honey bee colony declines, Service de recherche du Congrès, (lire en ligne)
- (en) Robyn Rose, Galen P. Dively et Jeff Pettis, « Effects of Bt corn pollen on honey bees: emphasis on protocol development », Apidologie, EDP Sciences, vol. 38, no 4, , p. 368-377 (ISSN 0044-8435, DOI 10.1051/apido:2007022, résumé, lire en ligne, consulté le )
- (en) Peng Li et al., « Amino acids and immune function », British Journal of Nutrition, Cambridge University Press, vol. 98, no 2, , p. 237-252 (DOI 10.1017/S000711450769936X, lire en ligne, consulté le )
- (en) Diana L. Cox-Foster et al., « A Metagenomic Survey of Microbes in Honey Bee Colony Collapse Disorder », Science, vol. 318, no 5848, , p. 283-287 (ISSN 0036-8075, DOI 10.1126/science.1146498, résumé)
- (en) Jean-Noël Tasei et Pierrick Aupinel, « Nutritive value of 15 single pollens and pollen mixes tested on larvae produced by bumblebee workers (Bombus terrestris, Hymenoptera: Apidae) », Apidologie, EDP Sciences, vol. 39, no 4, , p. 397-409 (ISSN 0044-8435 et 1297-9678, DOI 10.1051/apido:2008017, résumé, lire en ligne, consulté le )
- (en) K. P. Lee, S. J. Simpson et K. Wilson, « Dietary protein-quality influences melanization and immune function in an insect », Functional Ecology, John Wiley & Sons, Inc., vol. 22, no 6, , p. 1052-1061 (DOI 10.1111/j.1365-2435.2008.01459.x, lire en ligne, consulté le )
- (en) D. vanEngelsdorp, J. Jr. Hayes, R. Underwood et J. Pettis, « A Survey of Honey Bee Colony Losses in the U.S., Fall 2007 to Spring 2008 », PLoS One, vol. 3, no 12, (DOI doi:10.1371/journal.pone.0004071, résumé)
- (en) (en) COST, « Action FA0803: Prevention of honeybee Colony Losses (COLOSS) », COST, (consulté le )
- Agence française de sécurité sanitaire des aliments, Mortalités, effondrements et affaiblissements des colonies d’abeilles, (présentation en ligne, lire en ligne)
- (en) Mariano Higes, « Honeybee colony collapse due to Nosema ceranae in professional apiaries », Environmental Microbiology Reports, Society for Applied Microbiology, vol. 1, no 2, , p. 110-113 (DOI 10.1111/j.1758-2229.2009.00014.x, résumé)
- Pascale Mollier et al., « Le déclin des abeilles, un casse-tête pour la recherche », INRA Magazine, Institut national de la recherche agronomique, , p. 13-24 (lire en ligne)
- (en) Dennis vanEngelsdorp, Jay D. Evans, Claude Saegerman, Chris Mullin et al., « Colony Collapse Disorder: A Descriptive Study », PLoS One, vol. 4, no 8, (DOI doi:10.1371/journal.pone.0006481, lire en ligne, consulté le )
- (en) Dennis vanEngelsdorp, Jerry Jr. Hayes, Robyn M. Underwood et Jeffery S. Pettis, « A survey of honey bee colony losses in the United States, fall 2008 to spring 2009 », Journal of Apicultural Research, International Bee Research Association, vol. 49, no 1, , p. 7-14 (ISSN 2078-6913, DOI 10.3896/IBRA.1.49.1.03, résumé, lire en ligne, consulté le )
- (en) Potts Simon G. et al., « Declines of managed honey bees and beekeepers in Europe », Journal of Apicultural Research, International Bee Research Association, vol. 49, no 1, , p. 15-22 (ISSN 2078-6913, DOI 10.3896/IBRA.1.49.1.02, résumé, lire en ligne, consulté le )
- (en) Cédric Alaux, François Ducloz, Didier Crauser et Yves Le Conte, « Diet effects on honeybee immunocompetence », Biology letters, Royal Society Publishing, vol. 10, no 4, , p. 562-565 (ISSN 1744-957X, DOI 10.1098/rsbl.2009.0986, lire en ligne, consulté le )
- (en) Dennis vanEngelsdorp, Jerry Jr. Hayes, Robyn M. Underwood, Dewey Caron et al., « A survey of managed honey bee colony losses in the USA, fall 2009 to winter 2010 », Journal of Apicultural Research, International Bee Research Association, vol. 5O, no 1, , p. 1-10 (ISSN 2078-6913, DOI 10.3896/IBRA.1.50.1.01, résumé, lire en ligne, consulté le )
- (en) Andrew Core et al., « A New Threat to Honey Bees, the Parasitic Phorid Fly Apocephalus borealis », PLoS One, vol. 7, no 1, (DOI doi:10.1371/journal.pone.0029639, lire en ligne, consulté le )
- (en) Romée van der Zee et al., « Managed honey bee colony losses in Canada, China, Europe, Israel and Turkey, for the winters of 2008-9 and 2009-10 », Journal of Apicultural Research, International Bee Research Association, vol. 51, no 1, , p. 100–114 (ISSN 2078-6913, DOI 10.3896/IBRA.1.51.1.12, résumé, lire en ligne, consulté le )
- (en) Dennis vanEngelsdorp, Dewey Caron, Jerry Jr. Hayes, Robyn M. Underwood et al., « A national survey of managed honey bee 2010-11 winter colony losses in the USA: results from the Bee Informed Partnership », Journal of Apicultural Research, International Bee Research Association, vol. 51, no 1, , p. 115-124 (ISSN 2078-6913, DOI 10.3896/IBRA.1.51.1.14, résumé, lire en ligne, consulté le )
- Service Presse INRA, « Les abeilles désorientées par une faible dose d’insecticide », sur Institut national de la recherche agronomique, Institut national de la recherche agronomique, (consulté le )
- (en) Mickaël Henry et al., « A Common Pesticide Decreases Foraging Success and Survival in Honey Bees », Science, American Association for the Advancement of Science, vol. 336, no 6079, , p. 348-350 (ISSN 0036-8075 et 1095-9203, DOI 10.1126/science.1215039, résumé)
- (en) Penelope R. Whitehorn et al., « Neonicotinoid Pesticide Reduces Bumble Bee Colony Growth and Queen Production », Science, American Association for the Advancement of Science, vol. 336, no 6079, , p. 351-352 (ISSN 0036-8075 et 1095-9203, DOI 10.1126/science.1215025, résumé)
- (en) Erik Stokstad, « Field Research on Bees Raises Concern About Low-Dose Pesticides », Science, American Association for the Advancement of Science, vol. 335, no 6076, , p. 1555 (ISSN 0036-8075 et 1095-9203, DOI 10.1126/science.335.6076.1555, résumé)
- Jean-Daniel Charrière et al., « Guide de la santé de l'abeille », ALP Forum, Agroscope, no 84f, (ISSN 1661-0814, lire en ligne)
- (en) Autorité européenne de sécurité des aliments, « Statement on the findings in recent studies investigating sub-lethal effects in bees of some neonicotinoids in consideration of the uses currently authorised in Europe », EFSA Journal, Autorité européenne de sécurité des aliments, vol. 10, no 6, (ISSN 1831-4732, DOI 10.2903/j.efsa.2012.2752, résumé, lire en ligne, consulté le )
- Agence nationale de sécurité sanitaire de l'alimentation, de l'environnement et du travail, AVIS de l’Agence nationale de sécurité sanitaire de l’alimentation, de l’environnement et du travail relatif à une demande d’appui scientifique et technique dans la perspective de la publication de l'article « A common pesticide decreases foraging success and survival in honey bees », (lire en ligne)
- (en) Stephen J. Martin, Andrea C. Highfield, Laura Brettell1, Ethel M. Villalobos, Giles E. Budge, Michelle Powell, Scott Nikaido et Declan C. Schroeder, « Global Honey Bee Viral Landscape Altered by a Parasitic Mite », Science, American Association for the Advancement of Science, vol. 336, no 6086, , p. 1304-1306 (ISSN 0036-8075 et 1095-9203, DOI 10.1126/science.1220941)
- (en) James E. Cresswell et al., « Comment on “A Common Pesticide Decreases Foraging Success and Survival in Honey Bees” », Science, American Association for the Advancement of Science, vol. 337, no 6101, , p. 1453 (ISSN 0036-8075 et 1095-9203, DOI 10.1126/science.1224618, résumé)
- (en) Département de l'Environnement, de l'Alimentation et des Affaires rurales, An assessment of key evidence about Neonicotinoids and bees, Gouvernement du Royaume-Uni, (lire en ligne)
Professionnels de l'apiculture
- (en) Roger A. Morse et Nicholas W. Calderone, « The Value of Honey Bees As Pollinisators of U.S. Crops in 2000 », Bee Culture, A. I. Root Company, (lire en ligne, consulté le )
- François Servel, « Etude de la Maladie de la Disparition aux USA », Syndicat des apiculteurs professionnels de Midi-Pyrénées, Traduction et adaptation d'un article de William T. Wilson et Diana M. Menapace dans le numéro de février-mars 1979 de l'American bee journal.
- (en) Maryann Frazier, Interview, Dramatic honey bee die-off worries beekeepers and crop producers, podcast, College of Agricultural Sciences, Université d'État de Pennsylvanie, (consulté le ).
- (en) Krishna Ramanujan, « Parasites, pathogens and pesticides called possible suspects in honeybee decimation », Cornell Chronicle, Université Cornell, , p. 1, 6 (lire en ligne)
- (en) D. vanEngelsdorp, R. Underwood, D. Caron et J. Jr. Hayes, « An Estimate of Managed Colony Losses in the Winter of 2006 – 2007: A Report Commissioned by the Apiary Inspectors of America », American Bee Journal, Dadant & Sons Inc, no 147, , p. 599-603 (résumé, lire en ligne, consulté le )
- Jean Haxaire, « Le frelon asiatique Vespa velutina, un nouveau prédateur de l’abeille ? », La Santé de l'Abeille, Fédération nationale des organisations sanitaires apicoles départementales, no 220, (lire en ligne, consulté le )
- Vincent Tardieu, L'étrange silence des abeilles : Enquête sur un déclin inquiétant, Paris, Éditions Belin, coll. « Regards », , 352 p. (ISBN 978-2-7011-4909-7, présentation en ligne)
- « Bourdonnement autour du déclin de l'abeille domestique », sur Portail de la recherche et du développement en Europe, Cordis, (consulté le )
- Céline Holzmann, Hivernage et pertes de colonies chez les apiculteurs professionnels français, Institut technique et scientifique de l’apiculture et de la pollinisation, (présentation en ligne, lire en ligne)
- « Les assolements agricoles, sources ponctuelles et massives de fleurs pour les pollinisateurs », sur Jachères apicoles pour la biodiversité, Réseau Biodiversité pour les Abeilles (consulté le )
Presse
- Cécile Dumas, « Disparitions massives d'abeilles aux Etats-Unis », Sciences & Avenir, Groupe Perdriel, (lire en ligne, consulté le )
- (en) Gunther Latsch, « Are GM Crops Killing Bees? », Spiegel International, Bertelsmann, (lire en ligne, consulté le )
- (en) Maurice Possley, « Missing bees create a buzz », The Chicago Tribune, Tribune Company, (lire en ligne, consulté le )
- (en) Craig Mackintosh, « European Bees Also Taking a Nosedive - Perhaps GM Crops? », Celsias, (lire en ligne, consulté le )
- Denis Lebioda, « Le téléphone portable responsable de la disparition des abeilles ? », sur www.contaminations-chimiques.info, (consulté le )
- (en) Vincent Valk, « Albert Einstein, Ecologist? », Gelf Magazine, (lire en ligne)
- (en) Jia-Rui Chong et Thomas H. Maugh, « Experts may have found what's bugging the bees; A fungus that hit hives in Europe and Asia may be partly to blame for wiping out colonies across the U.S. », Los Angeles Times, (lire en ligne)Cet article, citant des travaux d'expert, évoque le champignon unicellulaire Nosema ceranae, mais dont la responsabilité pourrait être partagée avec deux autres champignons parasites et une demi-douzaine de virus également retrouvés dans les corps des abeilles mortes. S'il était prouvé que le Nosema ceranae était responsable, un traitement fongicide à la fumagillin serait possible.
- (en) Susan Kuchinskas, « Are Bees Too Busy? », East Bay Express, (lire en ligne, consulté le )
- Gaëlle Dupont, « Les abeilles malades de l'homme », Le Monde, Groupe Le Monde, (lire en ligne, consulté le )
- « Un virus soupçonné de tuer les abeilles », sur Le Figaro, Société du Figaro, (consulté le )
- Olivier Rescanière, « Les ruches de l'est de la France se dépeuplent de façon inquiétante », Le Monde, Groupe Le Monde, (lire en ligne)
- Loïc Chauveau, « Les abeilles ont besoin d'un plan d'urgence », Sciences & Avenir, Groupe Perdriel, (lire en ligne, consulté le )
- Bruno Scala, « Devenues zombies, des abeilles parasitées sortent la nuit », Futura-Sciences, SARL Futura-Sciences, (lire en ligne, consulté le )
- « Les abeilles du rucher école de la CAHC n'ont pas résisté aux ondes de la ligne haute tension », La Voix du Nord, Groupe La Voix, (lire en ligne)
- Martine Valo, « Il faut interdire l'usage des pesticides néonicotinoïdes en Europe », Le Monde, Groupe Le Monde, (lire en ligne)
Vidéo
- La disparition des abeilles : la fin d’un mystère (documentaire de 52 min. Mona Lisa production/Mandarava productions avec la participation de France 5 / Télé-Québec / RTBF).
Liens externes
- « Synthèse officielle Données apicole 2005 en France » (consulté le )
- Synthèse scientifique publiée par Greenpeace
- Colloque sur les mortalités d’abeilles aux États-Unis : les experts confirment la piste des pathologies- Agriculture & Environnement, le 19 décembre 2007
- Les abeilles malades de l'homme (Article du Monde, 29.08.07).
- Un monde sans fruits ni légumes ? (Article du Monde, 13.10.07).
- Et si les abeilles disparaissaient ? par la Cité des sciences et de l'industrie
- Jean Weissenberger, « Les abeilles dans l'UE: un bilan de santé inquiétant », Library Briefing, Bibliothèque du Parlement européen (consulté le )
- Apis mellifera : maladies et parasites
 Portail de l’entomologie
Portail de l’entomologie