Glandes péribuccales
Les glandes péri-buccales des Araignées, appelées aussi glandes maxillo-labiales, sont des organes sécréteurs localisés à la surface des lames maxillaires et de la lèvre inférieure.
Introduction
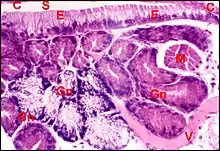
Désignées par Legendre (1953)[1], sous le nom de “plages glandulaires hypodermiques”, elles se présentent comme des épaississements tégumentaires localisés dans l'épiderme ("hypoderme") des lames maxillaires ou gnathocoxes et la lèvre inférieure ou labium dans le genreTegenaria. Ces structures existent en fait chez toutes les Araignées[2],[3]. Elles ne doivent pas être confondues avec les glandes gnathocoxales ou "salivaires" sous-jacentes (Fig.1).
Structure histologique
Ces épaississements ont un aspect en “bourrelet “ ou en “coussinet” dans les coupes histologiques ; ils s’y montrent constitués par de longues cellules épithéliales prismatiques se disposant en “palissade” (Fig.1) et recouvertes par une cuticule d’aspect strié.
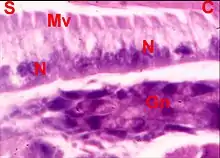
Les cellules sont pourvues de noyaux oblongs tous situés au même niveau et montrent un cytoplasme plus ou moins granuleux, vacuolaire (Fig.2), avec parfois des inclusions jaunâtres, et dont l’aspect évoque une activité glandulaire, sans permettre toutefois de l’affirmer. Les pôles apicaux sont pourvus d'une "brosse", faisceaux de microvilli vraisemblables.
Ultrastructure
Les premières études ultrastructurales du tégument gnathocoxo-labial[4], ont utilisé comme matériel Leptoneta microphthalma [2]et surtout Wendilgarda mustelina arnouxi[3] dont la glande labiosternale a fait l'objet d'une étude séparée.
L’aspect ultrastructural de l'épithélium gnathocoxo-labial est celui d’un épiderme sécréteur que surmonte un revêtement cuticulaire modifié.
Epithélium
Il est formé par des cellules toutes semblables, présentant des caractères d’adénocytes mais dépourvues de canalicules excréteurs comme celles de la glande rostrale. De ce fait, il ne comporte pas d’unités glandulaires structurées contrairement à la glande clypéale ou acronale.
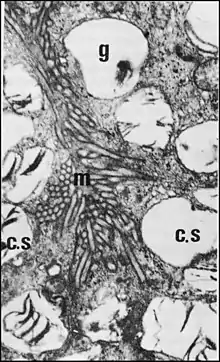
L’adénocyte est allongé et étroit.Son pôle basal est découpé par des invaginations du plasmalemme en fines languettes rappelant des prolongements podocytaires. Il repose sur une lamina continue, finement granulaire, pouvant être plaquée contre celle des acini de glandes gnathocoxales (“ glandes salivaires ”) adjacentes.
Son pôle apical (Fig.3) est fortement convexe et garni, sur toute sa surface, de microvillosités petites, irrégulières, enchevêtrées, renfermant des microfilaments et s’accolant, par place à celles d’adénocytes voisins ; elles bordent un espace extracellulaire séparant les adénocytes de la cuticule sus-jacente ou s’accolent à cette dernière. .
Les faces latérales sont à peu près planes, ne s’engrènent pas les unes dans les autres et remontent beaucoup moins haut que le sommet du pôle apical. Elles sont réunies entre elles par de petites jonctions.
Le noyau ovoïde siège dans le tiers basal de la cellule, contient un petit nucléole central d’aspect réticulé et une chromatine assez abondante, disposée en mottes irrégulières s’accolant à l’enveloppe nucléaire ou disséminées dans le nucléoplasme.
Les organites subcellulaires sont des mitochondries longues et flexueuses, des ribosomes libres, des rosettes de glycogène et surtout, un réticulum endoplasmique lisse extrêmement dense ainsi que des vésicules.
_et_cuticule.jpg.webp)
Ces dernières semblent prendre naissance à partir du réticulum lisse dilaté mais pourraient être aussi des éléments golgiens. Chez Leptoneta, elles nous ont paru arrondies, isolées ou confluentes, dépourvues de sécrétion ou contenant un matériel opaque marginal. En revanche, chez Wendilgarda, elles se présentent comme des grains de sécrétion nombreux, de contour irrégulier, plus ou moins anguleux et adoptent même parfois une forme en plaquette ou en bâtonnet. La membrane qui les délimite est fine et nettement dessinée. Le contenu du grain est un matériel peu dense, s’accolant à cette membrane et formant des lamelles superposées en piles plus ou moins disjointes (Fig.3). Il semble que les grains s’ouvrent parfois les uns dans les autres et libèrent leur contenu dans l’espace extracellulaire (Fig.4).
Ce dernier forme une cavité très réduite s’interposant entre la cuticule (cavité sous-cuticulaire) et les pôles apicaux des adénocytes qu’elle sépare également les uns des autres jusqu’à la hauteur des jonctions et a un contenu de même densité que celui des grains de sécrétion.
Cuticule
Elle est ajourée par un ensemble de fenestrations nombreuses responsables de son aspect “strié” dans les coupes histologiques. Ces fenestrations
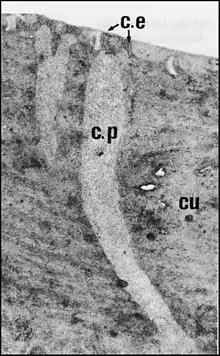
correspondent à des canaux poraires (pores-canaux) évoquant ceux décrits dans la cuticule des insectes. Ils sont assez mal limités chez Leptoneta, plus nets chez Wendilgarda (Fig.5), irréguliers, à peu prés parallèles mais plus ou moins ramifiés. Ils ne s’ouvrent pas directement à l’extérieur, semblent buter contre l’épicuticule et se résolvent, pour la traverser, en un faisceau de canalicules extrêmement grêles (15 nm) ("wax channels”)(Fig.5 : flèches). Leur contenu est identique à celui de l’espace extracellulaire (cavité sous-cuticulaire).
Commentaires
Caractères anatomiques
Sur le plan anatomique, l’étude ultrastructurale des coussinets “hypodermiques” gnathocoxaux montre qu’il s’agit bien de glandes tégumentaires. L’opinion de Legendre[1] basée sur la seule histologie et selon laquelle leurs “ cellules épithéliales présentent tous les caractères de cellules glandulaires exocrines ”se trouve ainsi confirmée.
De plus, ces mêmes éléments, recouverts par la cuticule comme des cellules “ hypodermiques” banales, peuvent être rattachés aux glandes tégumentaires arthropodiennes de classe 1 telles que Noirot et Quennedey (1974)[5] les ont définies chez les Insectes (Schéma)
L’adénocyte est recouvert par la cuticule et sa sécrétion doit traverser cette barrière pour être émise à l’extérieur. Chez les Araignées, d'autres glandes de classe 1 sont également connues dans les conduits de copulation de l'appareil génital femelle entélégyne[6], dans la spermathèque femelle et le tube séminifère mâle de Telema tenella[7],[8]. Il s'agit toujours d'organes simples en "coussin" ou formant une paroi de canal.
Les glandes de classe 1 sont beaucoup plus répandues dans la classe des Insectes où leur différenciation est d'ailleurs plus poussée (invagination fréquente, association avec la classe 3), par exemple dans la glande de Gilson des Phryganes [9], les glandes frontales et sternales des Termites[10], la glande sternale tubuleuse des Coléoptères Bathysciinae[11], les glandes à phéromones sexuelles de Bombyx mori[12], les glandes cirières d'autres Lépidoptères[13]
Cette sécrétion semble élaborée par le réticulum endoplasmique lisse. Toutefois, on sait par ailleurs que dans de nombreuses glandes exocrines d’Insectes (salivaires, dermiques, annexes du tractus génital), l’appareil de Golgi, non disposé en dictyosomes, paraît uniquement formé de vésicules. Il en est probablement de même pour les glandes péribuccales où les amas vésiculaires marqueraient l’emplacement de réseaux transgolgiens. La sécrétion y est concrétée en lamelles chez Wendilgarda mustelina arnouxi, peut-être aussi chez Leptoneta où l’aspect différent des vésicules ne traduirait qu’un autre stade du même processus sécrétoire.
Quoi qu’il en soit, elle est très différente des grains de pigment ou de guanine. De plus, l’abondance du réticulum lisse suggère que des lipides en forment au moins une partie. Elle est émise à l’extérieur par les canaux poraires et les canalicules épicuticulaires.
Sur le plan ultrastructural, les glandes des gnathocoxae et du labium s’apparentent donc à la l'organe rostral dont les adénocytes possèdent, eux aussi, un réticulum endoplasmique lisse abondant.
Caractères fonctionnels
Sur le plan fonctionnel, leur sécrétion pourrait jouer un rôle de lubrifiant dans des zones soumises aux frottements comme déjà envisagé chez les Tegenaria[1].
Elle pourrait intervenir aussi dans le nettoyage de la cavité buccale, dans la digestion extra-orale mais en aucun cas comme “ coussinet amortisseur ”[1], le cytosquelette des adénocytes étant trop peu développé, encore moins comme “ organe olfactif ”, hypothèse de Dahl[14]en 1885.
Bibliographie
- Legendre, R., 1953- Recherches sur les glandes prosomatiques des Araignées du genre Tegenaria. Ann. Univ. sarav.Sci 2, 4, p. 305-333.
- Lopez, A.avec M.Emerit, 1978 – Le dimorphisme sexuel gnathocoxal de Leptoneta microphthalma Simon (Araneae, Leptonetidae). Rev. Arachnol., (1), p. 1-15.
- Lopez, A., avec L. Juberthie-Jupeau et C. Ribera, 1980.- Les glandes gnathocoxales des Leptoneta (Araneae, Leptonetidae). Structure, ultrastructure et intérêt systématique. C.R.Ve Coll. Arachnol. express. franç., Barcelona, 1979, p. 163-178.
- Lopez, A. avec L. Juberthie-Jupeau), 1993 – Ultrastructure des glandes labio-sternales et péri-buccales chez Wendilgarda mustelina arnouxi Lopez & Emerit (Araneae, Theriiosomatidae). Rev. Arachnol., 10 (4), p. 75-92.
- Lopez, A. avec M. Emerit, 1985. Wendilgarda mustelina arnouxi n. ssp et la glande labio-sternale des Theridiosomatidae (Araneae). Mémoires de Biospéologie 12, p. 67–76
Notes et références
- Legendre, R., « Recherches sur les glandes prosomatiques des Araignées du genreTegenaria », Ann. Univ. sarav.Sci. 2, 4: 305-333.,
- Lopez, A., Juberthie-Jupeau,L. et C. Ribera, « Les glandes gnathocoxales des Leptoneta (Araneae, Leptonetidae). Structure, ultrastructure et intérêt systématique. », C.R.Ve Coll. Arachnol. express. franç., Barcelona, 1979, p. 163-178.,
- Lopez, A. & L. Juberthie-Jupeau, « Ultrastructure des glandes labio-sternales et péri-buccales chez Wendilgarda mustelina arnouxi Lopez & Emerit (Araneae, Theriiosomatidae). », Rev. Arachnol., 10 (4), p. 75-92.,
- Lopez, 1980,1993
- (en) Noirot, Ch. & A.Quennedey,, « Fine structure of insect epidermal glands. », Ann.Rev.Entomol.,19, p.61-80.,
- Kovoor,J., « Une source probable de phéromones sexuelles : les glandes tégumentaires de la région génitale des femelles d' Araignées. », C.R. VIème Coll. Arachnol. express. franç., Modena-Pisa 1981 Atti Soc. Tosc. Sci. Nat., Mem., ser. B, 88, suppl. (1981). p.1-14.,
- Lopez,A.& L.Juberthie-Jupeau, « Structure et ultrastructure de la spermathèque chez Telema tenella Simon (Araneae, Telemidae). », Mem. Biospéol., 10, p. 413-418.,
- Lopez,A. & L.Juberthie-Jupeau, « Ultrastructure comparée du tube séminifère chez les mâles d’Araignées. », Mem. Biospéol., 12, p. 97-109.,
- Quennedey, A., « Les glandes de Gilson des larves de Phryganea varia Fab. (Insecta, Trichoptera). Etude histochimique et ultrastructurale. », Journal Microsc., 8, p. 479-496.,
- Quennedey, A., « Les glandes exocrines des termites : ultrastructure comparée des glandes sternales et frontales. », Thèse de doctorat, Biologie, Dijon, Université de Bourgogne, 1978, 254 pp.,
- Juberthie-Jupeau,L. et M.Cazals, « Etude ultrastructurale de la glande sternale tubuleuse chez les Bathysciinae souterrains. », Mém.Biospéol.,10, p. 439-444.,
- (de) Steinbrecht, R.A., « Feinstruktur und Histochemie der Sexualduftdrüse des Seidenspinners Bombyx mori L. », Z. Zellforsch.und Mikrosk. Anat., 64, p.227–261.,
- (en) Locke,M., « The Cuticle and Wax Secretion in Calpodes Ethlius (Lepidoptera, Hesperidae). », Quart.J.Microsc.Sci.,101, p.333-338.,
- (de) Dahl,F., « , - Das Gehör und Geruchsorgan der Spinnen. », Arch. mikr. Anat. 24, 1–11.,
 Portail de l’arachnologie
Portail de l’arachnologie  Portail de la biologie cellulaire et moléculaire
Portail de la biologie cellulaire et moléculaire