Hyphe
Hyphe[n 1] est un élément végétatif filamenteux, souvent à plusieurs noyaux cellulaires (multinucléaire), caractéristique des champignons, de certaines algues et de certains protistes végétaux. Elle peut mesurer plusieurs centimètres de long mais n'avoir que quelques microns de diamètre et donc, à l'état isolé, être invisible à l’œil nu. En mycologie, c’est le constituant essentiel du sporophore et du mycélium[1]. Ces filaments deviennent visibles lorsqu'ils sont réunis en cordons suffisamment gros ; on parle alors de mycélium.


Le mot hyphe reflète davantage une ressemblance superficielle qu'une réalité morphologique commune à des taxa très éloignés phylogénétiquement, et doit donc être utilisé avec précaution.
L’hyphosphère est la région du sol directement formée et influencée par les hyphes fongiques et les micro-organismes associés qui jouent un rôle important dans la croissance des champignons et dans leur capacité à former des mycorhizes (bactéries auxiliaires de la mycorhization (en)).
Étymologie
Le terme hyphe vient du latin scientifique hypha, dérivé du grec ancien ὑφή / huphḗ signifiant « tissu »[2].
Les Groupes et espèces produisant des hyphes
Plusieurs espèces, appartenant à des lignées différentes, produisent des hyphes :
- beaucoup chez les « vrais » champignons (Eumycota) et pseudochampignons[n 2] (Pseudomycota) apparentés aux algues[3], comme les mildious ;
- chez certaines bactéries filamenteuses comme l'Actinomyces et Streptomyces ;
- chez des algues (le terme « hyphe » désigne alors les filaments cloisonnés internes qui constituent par exemple la moelle des laminaires).
Les hyphes peuvent être « cloisonnées » et elles sont alors dites « septées », ou « non cloisonnées » et elles sont dites « siphonnées » ou « coenocytiques ». L'éventuel cloisonnement ne constitue alors pas nécessairement des cellules, au sens où plusieurs noyaux distincts peuvent être contenus entre deux cloisons (ou « septes »).
Caractéristiques
Chez les champignons et pseudochampignons
On peut distinguer :
- les hyphes dites fondamentales qui forment l'armature du tissu fongique, d'assez fort diamètre : 8-10-15 µm, voire davantage.
- Les hyphes connectives, plus grêles : vers 2-4-6 µm, reliant les uns aux autres les divers éléments hyphiques.
- Les hyphes primordiales (sensu Fayod) désignent des hyphes les plus superficielles de la cuticule.
- Les hyphes dites excrétrices, ou mieux nommées sécrétrices : se subdivisent en hyphes lacticifères et hyphes oléifères, encore dites vasiformes.
Chez la plupart des fungi et chez des pseudochampignons multicellulaires, l'appareil végétatif de nutrition se compose d'éléments filamenteux de base appelés hyphes.
D'autres espèces, appelées levures, sont constituées de cellules uniques formant des colonies. Il existe aussi des cas où le thalle peut passer à divers moments de son cycle de vie sous les formes filamenteuse ou unicellulaire[3]. Enfin, il existe des formes trophiques particulières comme les cellules à rhizoïdes, plasmodes, protoplastes, etc.
Chez la plupart des espèces, quand une spore germe, elle produit un tube germinatif qui en croissant donne une hyphe filamenteuse. La ramification et l'entremêlement des hyphes donnent un mycélium.
Une des caractéristiques qui différencient les hyphes est la présence ou l'absence de parois transversales appelées septa (septum au singulier).
Généralement, les Oomycètes, les chytridiomycètes et les Zygomycètes ont des hyphes non cloisonnées ou siphons. Les filaments ont alors une structure cénocytique, c'est-à-dire qu'ils contiennent de nombreux noyaux non séparés les uns des autres[4].

Par contre, les Ascomycètes, les Basidiomycètes et leurs états asexués associés ont des hyphes cloisonnées (ou septées). Chaque compartiment comporte alors un ou deux (ou plus) noyaux. Ces noyaux peuvent d'ailleurs être génétiquement différents dans le cas d'anastomose (fusion) de filaments génétiquement différents. Le mycélium est alors dit hétérocaryotique ; sinon, c'est-à-dire si les noyaux sont tous issus de la germination de la même spore, il est dit homocaryotique[5]. Ainsi les cellules des hyphes de beaucoup de Basidiomycètes contiennent deux noyaux génétiquement différents.
Dans certains cas, ces hyphes s'accolent étroitement et s'organisent en faux-tissus appelés plectenchymes, constituants essentiels des sporophores ou de la partie fongique des lichens, par exemple. À l'extrême inverse, certaines hyphes sont réduites évolutivement à des cellules isolées entourées d'une paroi (cas des levures).
Des hyphes de champignons émergent des spores. Les hyphes caractérisent notamment les Deutéromycètes (catégorie qui rassemble les champignons dits « imparfaits » (Fungi imperfecti) dont les Penicillium, Fusarium, et Aspergillus).
Chez les bactéries filamenteuses
Comme les champignons filamenteux, les bactéries filamenteuses forment un réseau d'hyphes, mais ces hyphes sont de petite taille et les colonies formées dépassent rarement le centimètre. Le réseau d'hyphes n'est pas visible à l’œil nu (une observation au grossissement x10 est en général nécessaire pour voir l'aspect filamenteux de la périphérie de la colonie).
Chez les plantes
Le mot hyphe est chez les botanistes parfois synonyme de thalle (appareil végétatif des plantes dites inférieures ou primitives)
Physiologie de la croissance
Chez les champignons et pseudochampignons
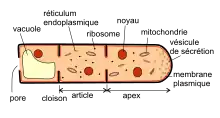
Les hyphes s'allongent par leur apex. Contrairement aux cellules animales ou végétales, la « cellule fongique » ne se divise pas[3]. La croissance se fait uniquement à son sommet. Elle peut s'accompagner de la formation de « cloisons » délimitant une structure ressemblant à une cellule et qu'on appelle article. Chez certaines espèces, ces cloisons sont percées d'un trou, appelé pore. Ils assurent une connexion entre les articles voisins.
Les études de l'ultrastructure (au microscope électronique) ont révélé la présence de beaucoup d'organelles suivant un fort gradient de concentration vers l'apex. Une intense activité biosynthétique produit des vésicules de sécrétion qui sont dirigés vers l'apex où elles fusionnent avec la membrane plasmique. C'est par ce moyen que la croissance polarisée de l'hyphe se fait. De plus ces vésicules contiennent de nombreux enzymes qui sont relâchés dans le milieu et qui vont permettre au champignon de digérer les matières organiques qui l'entourent. Croissance et assimilation des nutriments vont donc de pair.
Dans les cellules plus âgées des vacuoles apparaissent et peuvent envahir la totalité de l'article[3].
Hyphes fongiques et fixation du carbone et du calcium
Au moins depuis les années 1970, il est constaté que des aiguilles de calcite, microstructures fines et fragiles, sont fréquemment trouvées dans le sol. Elles sont présentes en grande quantité, dans la zone d'interface sol-système mycélien, en zone tempérée comme dans divers milieux tropicaux carbonatés[6],[7] ou arides[8] ou dans les sols organiquement très riche à forte activité biologique.
Ces mousses d’aiguilles microscopiques intriguaient les minéralogistes. Constituées d’aiguilles de calcite (non aragonite), elles étaient bien connues dans les milieux dits « supergènes » (sols ou roches altérées en milieu non saturé, dans la couche également prospectée par les racines de plantes et les hyphes fongiques) mais jusqu’au début des années 1980 les minéralogistes les considéraient comme d’origine abiotique, les classant comme simples espèces minérales (lublinite), dites « wiskers » par certains spécialistes des karsts. Certains auteurs, Comme Durand dans le nord-Est de la France (en 1978)[9] ou comme M. Pouget en 1980 en Algérie [10] ou encore comme Regaya en 1983[11] ont noté qu’elles étaient particulièrement présentes dans les encroûtements calcaires. Cette même année 1983, Callot et al. constatent que souvent ces genèse d’entrelacs de microaiguilles calcaires sont « associées à des structures racinaires en cours de minéralogenèse »[12].
En 1982 V. Verges avait montré que ces entrelacs de cristaux en en forme d’aiguilles sont, en fait, des concentrations calcitiques (et non aragonitiques), à caractère monocristallin[13]. Elles étaient alors encore considérées « des structures hyphanitiques appartenant au plasma du sol, par suite de leur très faible dimension (diamètre de 0,1 1pm à 1 gm), telles que L. Bal les définissait en 1975[14]. Leurs qualitificatifs évoquaient cependant des formes vivantes puisqu’on parlait de « pseudomycéliums », de « structure hyphanitique » à « croissance dendritique », etc...
Au milieu des années 1980, trois agronome français (Gabriel Callot, André Guyon et Daniel Mousain) ont étudié avec attention ces aiguilles (à divers grossissements et en micro-dissection), confirmant qu’elles provenaient d’une concentration de calcaire (et d’un peu de silice) à l’intérieur des hyphes de Basidiomycètes. Au sein d’éboulis caillouteux constitués de fragments de calcaire très poreux, là où les eaux carbonatées se concentrent, Callot et ses collègues ont noté la formation (à partir des macropores de la roche) de calcaire blanchâtre, cotonneux, pulvérulent, toujours à la face inférieure des morceaux de roche calcaire [15]. Ce matériau, partiellement biogénique est un entrelac d'aiguilles de calcite entrelacées à des hyphes mycéliens (blancs ou bruns). Il peut former une couche atteignant 1 à 3 cm d’épaisseur [15].
Callot et ses collègues, en 1985, avaient déjà expérimentalement prouvé que du carbonate de calcium peut se former à l’extérieur d’hyphes de champignon[16]. Cette nouvelle étude montre que dans la nature, il peut aussi se former à l’intérieur de l’hyphe (peut être en lien avec la présence de granules de polyphosphates dans les hyphes, selon S Trullu et ses collègues en 1981[17]). Ceci montre que les champignons peuvent aussi, modestement, contribuer aux puits de carbone basés sur le CaCO3. Ces microsystèmes ont une importance écologique encore mal mesurée, mais ils constituent de fait à la fois un piège à calcium et un puits de carbone, et il est possible qu’il contribue à mieux conserver l’eau de condensation (rosée) et de ruissellement dans certains cailloutis ou sols superficiels sans cela très drainants.
- On parle de « calcitisation » des structures fongiques quand les micro-aiguilles calcaires se forment rapidement par concentration et cristallisation des sels à la suite du dessèchement de l’hyphe fongique ; les aiguilles sont alors droites (alors qu’en laboratoire sur des fragments de calcaire maintenus constamment humide par un goutte à goutte, elles sont courbes)[15].
- La déshydratation de l'hyphe est généralement estivale, mais peut aussi intervenir quand l’hyphe est présent dans un vide ou courant d’air qui l’expose plus facilement à la déshydratation. Ce qui « pourrait expliquer en partie la présence des aiguilles de calcite dans les macro-vides du sol où, par ailleurs, leur élongation n’est nullement freinée par des contraintes physiques » [15].
Articles connexes
Liens externes
Notes
- Une ou un hyphe, les deux genres sont possibles. Voir hyphe dans le Wiktionnaire ou sur le site du CNRTL. Le Larousse choisit le féminin.
- Les pseudochampignons sont des organismes fongiformes qui étaient classés autrefois parmi les champignons et qui se retrouvent maintenant dispersés, avec l’avènement de la classification phylogénétique, dans plusieurs clades non monophylétiques avec les Fungi (Silar et al, 2013).
Références
- Josserand, Marcel., La description des champignons supérieurs (basidiomycètes charnus) : technique descriptive, vocabulaire raisonné du descripteur, Paris, Editions Lechevalier, , 392 p. (ISBN 2-7205-0507-2 et 978-2-7205-0507-2, OCLC 11710544, lire en ligne)
- Définitions lexicographiques et étymologiques de « hyphe » dans le Trésor de la langue française informatisé, sur le site du Centre national de ressources textuelles et lexicales
- Philippe Silar et Fabienne Malagnac, Les champignons redécouverts., Belin,
- P. Bouchet, J.-L. Guignard, Y.-F. Pouchus, J. Villard, Les champignons, Mycologie fondamentale et appliquée, Masson (2e édition),
- John Webster, Introduction to Fungi, Cambridge, Cambridge University Press, , 841 p. (ISBN 978-0-521-01483-0 et 0-521-01483-2, lire en ligne)
- Bocquier G (1973) « Genèse et évolution de deux toposéquences de sols tropicaux du Tchad : interprétation biogéodynamique ». Orstom. Thèse, 350p |URL : http://horizon.documentation.ird.fr/exl-doc/pleins_textes/cahiers/PTP/18441.PDF
- Nahon D (1976) Cuirasses ferrugineuses et encroûtements calcaires au Sénégal occidental et en Mauritanie. Systèmes évolutifs: géochimie, structure, relais et coexistence (Vol. 44, No. 1). Thèse de Doctorat ès Scienc, Fac. Sci. Tech. St-Jérôme, Marseille, Mém. C.N.R.S. 44, 232 p |Persée-Portail des revues scientifiques en SHS|URL : https://www.persee.fr/doc/sgeol_0302-2684_1976_mon_44_1_2404
- Sehgal, J. L., & Stoops, G. (1972). Pedogenic calcite accumulation in arid and semi-arid regions of the Indo-Gangetic alluvial plain of erstwhile Punjab (India)—Their morphology and origin. Geoderma, 8(1), 59-72. 5R ésumé
- Durand D (1978) La pédogenèse en pays de craie dans le Nord-Est de la France. Thèse Doctorat ès Sciences, Institut de Géologie, Strasbourg, 175 p.
- Pouget M (1980) « Les relations sol végétation dans les steppes algéroises ». Thèse Doct. ès Sci., Paris, Travaux et documents ORSTOM, 116, 555 p (lien IRD)
- Regaya K (1983) Etude géologique de la formation des limons de Matmata (Sud Tunisien). Thèse de 3ème cycle, Marseille St-Jérôme, 121 p.
- Callot G, Chamayou H, Maertens C & Salsac L (1983) Interactions sol racine et incidence sur la nutrition minérale. I.N.R.A., Paris, 320 p.
- Verges V (1982) « Contribution à l'analyse et à la représentation cartographique des formations pédologiques en moyenne montagne calcaire ». Thèse 3 e cycle, Univ. Paris VII, Pedol. Aménagement des Sols, 223 p.
- Bal L (1975) « Carbonate in soil : A theorical consideration on and proposal for its fabric analysis. 1. - Crystic, calcic and fibrous plasmic fabric ». Neth. J. agric. Sci., 23, 18-35 et 163-176.
- Callot G, Guyon A & Mousain D (1985) « Inter-relations entre aiguilles de calcite et hyphes mycéliens ». Agronomie, 5(3), 209-216
- Callot G, Mousain D & Plassard C (1985) « Concentrations de carbonate de calcium sur les parois des hyphes mycéliens ». Agronomie. 5 (2) |URL : https://hal.archives-ouvertes.fr/hal-00884743/document
- Strullu D.G, Gourret J.P & Garrec J.P (1981) « Microanalyse des granules vacuolaires des ectomycorhizes, endomycorhizes et endomycothalles ». Physiol. Vég., 19, 367-378.
 Portail de la mycologie
Portail de la mycologie  Portail de la phycologie
Portail de la phycologie