Investissement parental
Dans la biologie de l'évolution, Robert Trivers[1] fut le premier à définir le terme d'investissement parental (IP) comme tout investissement par le parent dans une descendance individuelle qui en augmente la chance de survie (et par là la réussite de la reproduction), mais en ôtant au parent des possibilités d'investir dans une autre progéniture. En d'autres termes, cela représente ce que les parents dépensent (en temps, énergie, etc.) et qui profite à leur progéniture mais les empêche d'investir dans d'autres composants de leur valeur sélective, comme le bien-être de leur descendance déjà existante et future et leur propre bien-être[2],[1]. Ceux-ci[3] comprennent le bien-être de leur descendance déjà existante et future et leur propre bien-être[4],[5].
Le biologiste Ronald Aylmer Fisher évoque le concept de dépense parentale (c'est-à-dire la part de ressource parentale, temps ou énergie, investie dans les soins parentaux apportés à la progéniture) dans son ouvrage Théorie génétique de la sélection naturelle publié (en) publié en 1930, Trivers introduisant en 1972 le terme d’investissement parental comme unité fondamentale pour mesurer toutes les ressources qu’un parent investit sur sa progéniture. Le biologiste Robert David Siegel (en) évoque le même concept sous le nom d’effort parental[6].
Dans la théorie des histoires de vie, les investissements reproducteurs peuvent être compris en termes de compromis pour l'allocation de ressources entre la reproduction et l'investissement parental. Trivers observe une asymétrie d'investissement entre les mâles et les femelles, ces dernières, chez de nombreuses espèces animales, investissant davantage de ressources dans leur descendance que les mâles. Cette différence d’investissement est due à l'anisogamie (investissement initial disproportionné dans peu d'ovules mais bien plus gros que les nombreux petits spermatozoïdes produits pas les mâles), et au fait que, bien souvent, les femelles participent davantage aux soins aux jeunes que les mâles. Ces derniers augmentent leur succès reproducteur via une augmentation de l'accès aux femelles réceptives (sélection intersexuelle et sélection intrasexuelle)[7].
Diversité de l'investissement parental
Diversité de forme
Les soins parentaux peuvent être fournis à tout stade de vie de la progéniture : soins prénataux, y compris des comportements comme la surveillance des œufs, la préparation du nid, le port du couvain, l'incubation et la nourriture chez les mammifères placentaires, ainsi que soins postnataux, qui comprennent l'approvisionnement en nourriture et la protection des petits.
Soins parentaux au sein du vivant
« Les soins parentaux incluent au sens large la préparation des nids et des terriers, la production d'œufs garnis de substances de réserve, les soins apportés aux œufs et aux jeunes à la fois à l'intérieur et à l'extérieur du corps du géniteur, l'alimentation des jeunes avant et après la naissance, et les soins éventuellement apportés après l'indépendance nutritionnelle des jeunes[8] ».
Ces soins se retrouvent dans un vaste éventail de groupes taxonomiques comprenant à la fois des espèces ectothermiques (invertébrés, poissons, amphibiens et reptiles) et endothermiques (oiseaux et mammifères). Les biologistes observent une large diversité de situations chez les amphibiens et les invertébrés. L'histoire évolutive chez les autres groupes taxonomiques montre des situations plus marquées, avec de réelles différences d’investissement entre mâles et femelles.
Répartition des soins parentaux chez différents groupes :
- Invertébrés : Rare, mais lorsque cela se produit ce sont souvent les femelles seules. Toutefois, il existe un certain nombre de cas de soins menés par le mâle, comme chez les Belostomatidae[9],[10].
- Poissons : Le ratio mâle seulement/Biparental/Femelle seulement est de 9/3/1[11].
- Amphibiens : Femelles seules et Mâles seuls tout aussi communs. Faible fréquence d’un soin biparental[12],[13].
- Reptiles : Soit les femelles seules soit les deux parents[11].
- Oiseaux : Dans 90 % des espèces, il y a un soin biparental (dont 9 % avec une aide au soin d’apparentés), mais les femelles investissent souvent plus dans le soin. Dans les espèces restantes il y a un soin de la part des femelles seules, les cas de mâles prodiguant seuls les soins étant plus rares[14],[15].
- Mammifères : Soin des femelles dans toutes les espèces. Dans 95 % des espèces, les femelles fournissent seules le soin. Dans 5 % des cas les mâles aident aux soins. Dans aucun cas les mâles sont seuls à investir dans le soin[16]. Chez les mammifères, les soins biparentaux sont associés à la monogamie sociale[17].
Valeur sélective de l'investissement parental

Coûts et bénéfices liés à l'investissement parental
La reproduction est un processus coûteux en matière de temps, de ressources et d'énergie. L'investissement parental peut se faire au détriment de la condition et de la survie future d'un individu ainsi que de sa capacité de reproduction à venir (et donc de la valeur reproductive du parent). Il peut aussi engendrer des conflits comme le conflit parents-progéniture[18].
Pour exemple, chez le Bruant des prés, le nombre de jeunes bénéficiant de soins parentaux par leurs parents et le temps consacré aux soins affectent significativement le taux de survie des parents. Il existe donc une corrélation négative entre l’ampleur des soins parentaux (fonction du nombre de jeunes de la portée et donc du temps consacré aux soins parentaux) et le taux de survie des parents : on parle de trade-off. À noter que le taux de survie parental n’est pas affecté par le sexe du jeune auquel le parent a apporté ses soins[19].
Chez les espèces itéropares, où les mâles et les femelles connaissent au cours de leur vie plusieurs épisodes de reproduction, les parents sacrifient les avantages de l'investissement parental dans la descendance présente au profit de la reproduction à venir. En particulier, les parents ont besoin d'équilibrer les exigences de leur progéniture avec les leurs pour ne pas négliger leur propre survie.
Toutefois, les bénéfices de l'investissement parental en faveur de leur descendance sont importants et associés à une amélioration de la condition, de la croissance, de la survie et finalement, du succès reproducteur de la progéniture.
La sélection tend donc à maximiser chez les parents la différence entre les bénéfices et les coûts, et les soins parentaux auront tendance à exister lorsque les premiers sont beaucoup plus importants que les seconds. Globalement, l'investissement parental est sélectionné quand les avantages qui lui sont associés sont supérieurs aux coûts engendrés.
La théorie des jeux appliquée à l'investissement parental
En 1973, Maynard Smith & Price[20] appliquent la théorie des jeux à l’écologie comportementale et à l’évolution en introduisant le concept de stratégie évolutivement stable(ESS). Une stratégie est dite ESS si, lorsque toute la population l’a adoptée, aucune stratégie « mutante » ne peut envahir la population sous l’effet de la sélection naturelle.
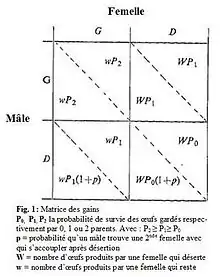
Maynard Smith (1977)[21] utilise ce concept afin de modéliser les différentes stratégies sexuelles que l’on peut rencontrer dans l’investissement parental. Notons qu’il se base sur la définition de Trivers de l’investissement parental (1972)et s’inspire de la critique apportée par Dawkins & Carlisle (1976)[22], selon laquelle la sélection aurait tendance à favoriser la stratégie qui maximise le nombre de progénitures qui sera produit par un individu dans le futur, indépendamment de ce que celui-ci a investi dans le passé. Pour chaque parent, il y a deux stratégies possibles : prendre soin de ses jeunes (G=Garder) ou les abandonner (D=Déserter). La décision de s’investir ou non dans les progénitures dépend des bénéfices et des coûts que ce choix entraine ainsi que des stratégies employées par les autres.
Prenons pour exemple, l’un des trois modèles que présente Maynard-Smith pour illustrer le concept de l’ESS. Le succès de la paire varie avec l’investissement pré-copulatoire (venant uniquement de la femelle) et post-copulatoire. Une femelle qui produit beaucoup d’œufs sera moins apte à en prendre soin et vice-versa. On obtient la matrice des gains suivante (Fig.1), avec quatre types d’ESS possibles.
Si le gain de l’investissement est supérieur à celui de déserter, le parent reste (G).Si le gain de l’investissement est inférieur à celui de désertion, le parent déserte (D). Pour déserter, un parent doit être presque aussi efficace que les deux parents réunis pour prendre soin des enfants et le parent déserteur doit avoir de bonnes chances de trouver un nouveau partenaire.
Notes et références
- (en) R.L. Trivers (dir.), Sexual selection and the descent of man, 1871-1971, Chicago, IL: Aldine, , 378 p. (ISBN 0-435-62157-2 et 9780435621575), « Parental investment and sexual selection »
- . (en) Tim H. Clutton-Brock, The Evolution of Parental Care, Princeton University Press, , 368 p. (ISBN 0-691-08730-X, 9780691087306 et 0691025169, OCLC 21975626)
- (en) John Beatty (dir.), Keywords in Evolutionary Biology, Cambridge (Mass.)/London, Harvard University Press, (réimpr. 1994), 432 p. (ISBN 0-674-50312-0 et 9780674503120, OCLC 25510478), « Fitness: theoretical contexts », p. 115-119
- (en) William Donald Hamilton, « The genetical evolution of social behaviour, I », Journal of Theoretical Biology, vol. 7, no 1, , p. 1-16 (ISSN 0022-5193, DOI 10.1016/0022-5193(64)90038-4)
- (en) William Donald Hamilton, « The genetical evolution of social behaviour, II », Journal of Theoretical Biology, vol. 7, no 1, , p. 17-52 (ISSN 0022-5193, DOI DOI:10.1016/0022-5193(64)90039-6)
- (en) Siegel RB, Weathers WW, Beissinger SR., « Assessing parental effort in a Neotropical parrot: a comparison of methods », Anim Behav., vol. 57, no 1, , p. 73-79 (DOI 10.1006/anbe.1998.0949).
- Étienne Danchin, Luc-Alain Giraldeau, Frank Cézilly, Écologie comportementale, Dunod, (lire en ligne), p. 241-324.
- Étienne Danchin, Luc-Alain Giraldeau, Frank Cézilly, Écologie comportementale, Dunod, (lire en ligne), p. 318.
- David W. Zeh et Robert L. Smith, « Paternal Investment by Terrestrial Arthropods », American Society of Zoologists, vol. 25, no 3, , p. 785–805 (DOI 10.1093/icb/25.3.785)
- Douglas W. Tallamy, « Sexual selection and the evolution of exclusive paternal care in arthropods », Journal of Evolutionary Biology, vol. 60, no 5, , p. 559–567 (DOI 10.1006/anbe.2000.1507)
- J.D. Reynolds, N.B. Goodwin et R.P. Freckleton, « Evolutionary transitions in parental care and live bearing in vertebrates », Philosophical Transactions of the Royal Society of London, vol. 357, , p. 269-281 (DOI 10.1098/rstb.2001.0930)
- C.W. Beck, « Mode of fertilization and parental care in anurans », Animal Behavior, vol. 55, , p. 439–449 (DOI 10.1006/anbe.1997.0619)
- K. Summers, C.S. McKeon et H. Heying, « The evolution of parental care and egg size: a comparative analysis in frogs », Proceedings of the Royal Society of London, vol. 273, , p. 687–692 (DOI 10.1098/rspb.2005.3368)
- D. Lack, « Ecological Adaptations for Breeding in Birds », Science, vol. 163, no 3872, , p. 1185-1187 (DOI 10.1126/science.163.3872.1185)
- A. Cockburn, « Prevalence of different modes of parental care in birds », Proceedings of the Royal Society of London, Series B, vol. 273, , p. 1375-1383 (DOI 10.1126/science.163.3872.1185)
- T. H. Clutton-Brock, « The Evolution of Parental Care », Journal of Evolutionary Biology, vol. 5, no 4, , p. 719–721 (DOI 10.1046/j.1420-9101.1992.5040719.x)
- (en) Ulrich H. Reichard et Christophe Boesch, Monogamy. Mating Strategies and Partnerships in Birds, Humans and Other Mammals, Cambridge University Press, (lire en ligne), p. 109.
- M. Kölliker & H. Richner. Parent-offspring conflict and the genetics of offspring solicitation and parental response dans Animal Behaviour 2001, 62: 395-407
- N.T. Wheelwright, K.A. Tice & C. R. Freeman-Gallant. ‘’Postfledging Parental Care in Savannah Sparrows: Sex, Size and Survival’’ dans ‘’Animal Behaviour’, 2003, 65 pp 433-445
- Maynard Smith, J. and Price, G.R. The logic of animal conflict, Nature 246:15-18 doi:10.1038/246015a0 (1973)
- John Maynard Smith,Parental investment: a prospective analysis. Animal Behaviour 25: 1–9.(1977)
- Richard Dawkins ; Carlisle, T.R. (1976). Parental investment, mate desertion and a fallacy. Nature 262: 131–133. doi:10.1038/262131a0
Annexes
Articles connexes
Bibliographie
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Parental investment » (voir la liste des auteurs).
- (en) J.R. Krebs (dir.), N.B. Davies (dir.), Tim H. Clutton-Brock et C. Godfray, Behavioural ecology : an evolutionary approach, Blackwell Science, (réimpr. 1984, 1991, 1996), 464 p. (ISBN 0-86542-731-3 et 9780865427310, OCLC 181161242), « Parental investment », p. 234-262
- (en) R.L. Trivers (dir.), Sexuel Selection and the Descent of Man, Blackwell Science, , 307 p., « Parental Investment and Sexual Selection », p. 136-179
- (en) Hanna Kokko et Michael D. Jennions, « Parental investment, sexual selection and sex ratios », Journal of evolutionary biology, vol. 21, no 4, , p. 919-948 (ISSN 1010-061X, DOI 10.1111/j.1420-9101.2008.01540.x)
- (en) John Maynard Smith, « Parental investment: a prospective analysis. », Animal Behaviour, vol. 25, no 1, , p. 1-9
- (en) Richard Dawkins et T.R. Carlisle, « Parental investment, mate desertion and a fallacy », Nature, vol. 262, , p. 131-133
- (en) Kolliker et Richner, « Parent–offspring conflict and the genetics of offspring solicitation and parental response », Animal Behaviour, vol. 62, , p. 395–407
 Portail origine et évolution du vivant
Portail origine et évolution du vivant