Lymphocyte NK
Les cellules tueuses naturelles, ou lymphocytes NK, pour Natural Killer, sont des cellules du système immunitaire inné cytotoxique qui n'éliminent pas directement les agents infectieux notamment en raison de leurs récepteurs invariants.
Pour les articles homonymes, voir NK.
.jpg.webp)
Les cellules NK représentent environ 5 à 16 % des lymphocytes humains[1] Ce sont des cellules provenant de la lignée lymphocytaire proche des cellules des lymphocytes T, mais ne possédant pas de récepteur membranaire aux antigènes comme les lymphocytes. Ce sont de grands lymphocytes granuleux (par opposition aux « petits lymphocytes »), non T (CD3-) non B (CD19-), caractérisés chez l'humain par les marqueurs CD56, CD16 et NK. Les récepteurs du lymphocyte NK sont synthétisés au cours du développement et de la maturation de la cellule puis ceux-ci ne subiront plus de changement[2].
Les cellules NK utilisent des récepteurs inhibiteurs (récepteur tueur d'immunoglobuline et Ly49) pour se développer, mûrir et reconnaître le « soi » du « non-soi ». Les cellules tueuses naturelles éliminent les cellules dont la fonction est altérée, comme les cellules tumorales ou les cellules infectées par un virus. Par exemple, les cellules tueuses naturelles peuvent reconnaître les cellules n'exprimant plus le CMH de classe I. Cette situation est observée durant une infection virale, certains virus pouvant induire la diminution du CMH de classe I pour éviter la reconnaissance par d'autres cellules immunitaires comme les lymphocytes T CD8[3].
Découvertes dans les années 1960, ces cellules tirent leur nom du fait qu'elles n'ont pas besoin de l'activation d'un lymphocyte CD4+ pour avoir une action cytotoxique[4],[5]. L'existence de lymphocytes cytotoxiques naturels ou lymphocytes NK (sigle de l'anglais Natural Killer, signifiant « tueur naturel ») aussi appelés cellules tueuses naturelles ayant des propriétés anti-tumorales intrinsèques et innées a été découverte lors d'expérience avec des lymphocytes T[6]. L'activité des cellules NK a d'abord été observée dans les cellules mononucléaires du sang périphérique humain[7],[8]; cependant, ses gros lymphocytes granuleux résident dans plusieurs tissus lymphoïdes et non lymphoïdes, notamment la moelle osseuse , les ganglions lymphatiques , la peau, l'intestin, les amygdales, le foie et les poumons[9].
En matière de cytotoxicité, la différence fondamentale entre les lymphocytes T CD8+ et les lymphocytes NK est que la cellule T CD8+ nécessite une activation par le T CD4+ auxiliaire pour exprimer sa cytotoxicité entraînant une synthèse de récepteurs spécifiques d'une espèce microbienne par recombinaison somatique. Comme leur nom l'indique, les cellules NK sont constitutivement cytotoxiques et, contrairement aux cellules T cytotoxiques, ne nécessitent pas d'exposition préalable à l'antigène pour médier leurs effets anti-tumoraux[10],[11].
Découverte
Les lymphocytes NK ont été découverts dans les années 1960[4],[12] et, en 10 ans, les chercheurs ont commencé à explorer une population de lymphocytes innés auparavant inconnue, et désignées aujourd'hui sous le nom de cellules tueuses naturelles (NK)[10],[13],[14],[11].
Développement et maturation fonctionnelle des cellules NK
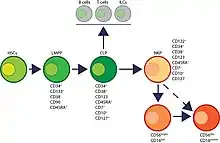
On pensait initialement que les cellules NK se développaient exclusivement dans la moelle osseuse. Cependant, des preuves récentes chez l'homme et la souris suggèrent qu'elles peuvent également se développer et mûrir dans les tissus lymphoïdes secondaires y compris les amygdales, la rate et les ganglions lymphatiques[15]. Les progéniteurs cellulaires et les populations intermédiaires qui donnent naissance aux cellules NK sont définis par l'expression différentielle de marqueurs de surface spécifiques à la lignée[16]. Les cellules tueuses naturelles représentent 5 à 20% des lymphocytes circulants chez l'homme[17].Chez l'homme, des sous-ensembles de cellules NK expriment le récepteur Fc activateur, CD16 et la plupart expriment CD56 [molécule d'adhésion des cellules neurales (NCAM) ou Leu-19][18],[19].
Différentes étapes
Les cellules souches hématopoïétiques multipotentes donnent naissance à tous les leucocytes et globules rouges dont une branche constitue le progéniteur lymphoïde commun. Les progéniteurs lymphoïdes communs donnent naissance à des cellules lymphoïdes innées Pro-B, Pre-T, et des lignées Pre-NKP[20]. Des travaux récents ont permis de délimiter un total de six étapes du développement des cellules NK humaines en fonction de leur développement à la fois moelle osseuse et ganglion lymphatique[15],[21] :
- stade 2a : les cellules CD3ε- CD7+ CD127+ marquent le stade le plus précoce des pré-NK ;
- stade 2b : est défini par l'expression de l'IL-1R, un récepteur de l'IL-1β ;
- stade 3 : l'expression des récepteurs d'activation, y compris NKG2D (CD314, lectine de type C, KLRK1), CD335 (récepteur de cytotoxicité naturelle, NCR1, NKp46) et CD337 marque la transition des cellules NK du stade 2b au stade 3.
- stade 4 : il est subdivisé en deux parties en fonction de l'expression du récepteur activateur NKP80[22],[23]. La principale distinction des cellules NK au stade 4a est qu'elles expriment des quantités abondantes de CD56 (CD56 bright). Ces cellules NK sont NKP80 - et expriment les niveaux maximaux de NKG2D, CD335, CD337, inhibiteur NKG2A et CD161 Au stade 4b, les cellules NK humaines deviennent positives pour NKP80 et conservent leur statut CD56 bright.

On distingue deux types de cellules NK chez l'être humain, regroupées selon la densité des marqueurs membranaires CD56 et CD16 :
- La régulation négative de l'expression de CD56bright pour devenir CD56dim dans la plupart des cas et l'expression du membre de la superfamille CD16 d'immunoglobulines (FcγRIII) dans un sous-ensemble de cellules NK définissent le stade 5. La plupart des cellules NK humaines dans le sang périphérique sont CD56dim. Les niveaux d'expression de CD56 fournissent une classification fonctionnelle des cellules NK humaines. La plupart des cellules NK humaines dans le sang périphérique sont CD56dim. Les cellules CD56bright NK sont considérées comme moins matures et résident principalement dans les tissus lymphoïdes secondaires tandis que le sous-ensemble CD56dim représente la majorité des cellules NK en circulation.
La plupart des cellules population immature de NK se transforment en une population CD56bright mineure (~5 %) qui se transforme en population CD56dim majeure (> 90 %). La régulation négative du CD56 pendant la maturation des cellules NK humaines est fortement associée à l'acquisition d'une cytotoxicité anti-tumorale car les cellules N56 CD56bright sont de puissants producteurs de cytokines inflammatoires, tandis que la fonction cytolytique des cellules NK humaines réside principalement dans la population CD56 dim[24],[25],[26].
Rôle des cytokines dans le développement des cellules NK
Les cytokines sont des médiateurs inflammatoires essentiels qui contrôlent de multiples aspects de la biologie des cellules NK. Les cellules NK expriment les récepteurs des cytokines au début de leur développement[27] et nécessitent une signalisation à travers la chaîne gamma commune (γc) pour leur développement, leur homéostasie et leur fonction[28]. La chaîne γc (CD132) est une glycoprotéine transmembranaire qui sert de sous-unité de signalisation pour IL-2, IL-4, IL-7, IL-9, IL-15 et IL-21[29]. Bien que ces cytokines présentent une certaine redondance fonctionnelle, leurs fonctions spécifiques aux cellules au cours d'une réponse immunitaire sont déterminées par l'expression de récepteurs distincts[30],[31]
.jpg.webp)
L'interleukine-2 et l'IL-15 sont des membres fonctionnellement apparentés de la famille γc des cytokines en ce qui concerne leurs interactions avec les récepteurs[32], .L'IL-2Rα (CD25) est exprimée sur les cellules NK activées et augmente considérablement leur affinité pour l'IL-2 qui stimule leur prolifération et la production de molécules lytiques telles que la perforine et le granzyme B[33]. Étant donné que les cellules NK se trouvent près des zones de cellules T dans les tissus lymphoïdes secondaires[9], l'IL-2 dérivée des cellules T peut faciliter un échange efficace vitale entre les lymphocytes innés et adaptatifs pendant une infection[34].
l'IL-15 est unique à cet égard. L'IL-15 interagit avec les cellules T Pour que cela se produise, l'IL-15 soluble se lie à l'IL-15Rα sur la surface des cellules qui présentent ce complexe aux cellules NK exprimant les hétérodimères IL2-Rβ / γc[35]. L'IL-15 peut être présentée par les cellules dendritiques et les macrophages ainsi que par les cellules non hématopoïétiques, y compris les cellules stromales et les cellules épithéliales[36]. IL-21 synergise avec IL-2 pour augmenter l'expression de NKG2A, CD25, CD86, CD69, Perforin et Granzyme B et ainsi augmenter la cytotoxicité[37]. Ces cytokines qui utilisent les récepteurs à base de γc sont le lien obligatoire entre les cellules NK et les cellules qui les produisent. Par exemple, les cellules T auxiliaires qui produisent de l'IL-21 peuvent réguler les niveaux d'expression des récepteurs d'activation ou du contenu cytolytique dans les cellules NK. De même, les CD qui produisent IL-15 jouent un rôle essentiel dans la prolifération et l'amorçage des cellules NK
Rôle des récepteurs d'activation codés par la lignée germinale
La forte cytotoxicité anti-tumorale est le résultat de sécrétion de quantité importante de cytokines pro-inflammatoires[38]. La cellule NK possède multiples récepteurs d'activation des cellules NK codés par la lignée germinale tels que NKG2D, NCR1, NCR2, NCR3, NKG2C, CD244, Ly49D et Ly49H. L'expression de plus d'un récepteurs d'activation qui reconnaissent les ligands du non soi ou pathogènes confère aux cellules NK des capacités innées de médiation des fonctions effectrices. En raison de l'expression de multiples récepteurs d'activation, les cellules NK doivent suivre un programme de développement distinct pour éviter une mauvaise reconnaissance de «soi» conduisant à des réponses auto-immunes. La nature variée des récepteurs d'activation et l'absence de domaines de signalisation dans leurs parties cytoplasmiques nécessitent l'association et le recrutement de co récepteurs pour la transduction du signal[39]. Les molécules coréceptrices qui propagent la signalisation des récepteurs NK comprennent FcεRIγ, CD3ζ et le DAP12 qui signalent via des motifs d'activation à base de tyrosine (ITAM) contenus dans leurs domaines cytoplasmiques. Les récepteurs d'activation qui utilisent ces corécepteurs incluent CD16, NCR1, Ly49D, Ly49H et NKG2D[40],[41],[42],[43] . Cependant, Ly49H et NKG2D peuvent également signaler via le motif YINM présent dans le corécepteur, DAP10[44],[45],[46] L'activation des cellules NK à travers ces corécepteurs se produit en interagissant avec des ligands cellulaires et étrangers distincts présents sur les cellules malades et forme la base de la réponse immunitaire médiée par les cellules NK dans de multiples contextes.
Fonctions des cellules NK
Les cellules tueuses naturelles médient leurs effets grâce à deux fonctions effectrices essentielles. Premièrement, les cellules NK sont des lymphocytes cytotoxiques qui peuvent directement lyser des cellules qui ont subi une transformation maligne ou qui ont été infectées par un virus ou un autre pathogène intracellulaire[47]. La fonction cytolytique des cellules NK peut s'initier à travers une variété de processus, y compris la dégranulation et la stimulation des récepteurs de la mort, et est essentielle pour l'élimination des cellules malades et dysfonctionnelles[48],[49]. Deuxièmement, les cellules NK peuvent produire une variété de cytokines inflammatoires en réponse à la stimulation des récepteurs et l'activation induite par les cytokines inflammatoires[50],[51]. Ces fonctions effectrices des cellules NK sont des composantes essentielles de la réponse immunitaire et sont les principaux mécanismes par lesquels les cellules NK assurent l'immunité .
Rôles cytotoxiques par activation des récepteurs NK
Il existe sur la membrane cellulaire des NK des récepteurs activateurs (portant des séquences « ITAM ») : immunoreceptor tyrosine-based activation motif) ou inhibiteurs (portant des séquences « ITIM » : immunoreceptor tyrosine-based inhibition motif). Lorsqu'un NK rencontre une autre cellule, la lyse de cette cellule ne se produira que si les signaux d'activation surpassent les signaux d'inhibition. Le principal signal inhibiteur est produit par les récepteurs KIR (acronyme de l'anglais « killer cell Ig-like receptor »), portés par le NK, qui reconnaissent les molécules du CMH de classe I. L'activation d'un seul type de récepteur KIR suffit à empêcher l'activation du NK alors qu'il faut toujours plusieurs signaux activateurs différents pour provoquer la dégranulation du NK et la mort de la cellule non reconnue. Les signaux d'activation sont variés, et comportent notamment des protéines produites par des cellules stressées, par exemple lors d'une infection. Ce système d'équilibre dynamique activation / inhibition permet en pratique aux cellules NK de lyser toutes cellules dépourvues des molécules du CMH de classe I (dont théoriquement tous parasites extracellulaires) ou cellules infectées par des virus ou des bactéries tout en épargnant les cellules saines. La lyse des cellules cibles se fait principalement par les voies perforine / granzyme et l'interféron gamma.
Cytotoxicité cellulaire dépendante des anticorps
L'ADCC ou antibody-dependent cell-mediated cytotoxicity, est permise par l'expression du marqueur CD16 à la surface des cellules NK. Le CD16, nommé aussi FcγRIII, est un récepteur de fragment constant d'immunoglobulines telles que l'IgG1 et l'IgG3. Il semblerait que les cellules CD56bright, qui expriment peu de CD16, soient moins efficaces dans les mécanismes d'ADCC que les cellules CD56dim.
Une autre voie par laquelle les cellules NK reconnaissent les cellules cibles potentielles dépend du fait que des cellules tumorales et des cellules infectées par certains virus exposent des antigènes contre lesquels le système immunitaire a développé une réponse anticorps, de telle façon que des anticorps antitumoraux ou antiviraux soient liés à leur surface. Étant donné que les cellules NK expriment le CD16, qui est un récepteur membranaire pour l'extrémité carboxy-terminale de la molécule d'IgG, appelée Fc (cf. anticorps), elles peuvent fixer à ces anticorps et, par la suite, lyser les cellules ainsi marquées. Ceci est un exemple d'un processus connu sous le nom de cytotoxicité cellulaire dépendante des anticorps (ADCC, antibody-dependent cell-mediated cytotoxicity).
Sécrétion de cytokines
Les cellules tueuses naturelles sont de puissants producteurs de cytokines pro-inflammatoires et immunosuppressives. Cependant, la libération de cytokines inflammatoires est distincte de la sécrétion de granules cytotoxiques[52] et les cellules NK utilisent des voies de signalisation induits par activation pour réguler ces deux fonctions[53] . Bien que les cellules NK puissent produire une large gamme de cytokines en fonction de l'environnement inflammatoire , les cellules NK produisent principalement des cytokines de type produit par le Th1 lorsqu'elles répondent aux ligands tumoraux et aux agents pathogènes intracellulaires[54],[55] Il s'agit notamment de l'IFN-γ, du TNF et du facteur de stimulation des colonies de granulocytes / monocytes (GM-CSF) qui facilitent l'activation des cellules T les cellules dendritiques, les macrophages et les neutrophiles[56],[57]. Les cellules NK produisent également des cytokines chimiotactiques (chimiokines), notamment CCL3 (MIP-1α), CCl4 (MIP-1β), CCL5 (RANTES), XCL1 (lymphotoxine) et CXCL8 (IL-8) qui peuvent attirer les lymphocytes effecteurs et les cellules myéloïdes vers un site d'inflammation[58].

L'activation des voies de transcription des molécules cytolytiques et des cytokines inflammatoires est un processus hautement régulé médié par une variété de régulateurs dans les cellules NK. Beaucoup de ces facteurs de transcription, tels que le T-bet, définissent la lignée et s'activent tôt dans le développement des cellules NK[15]. L'activation induite par les cytokines de facteurs de transcription, tels que les transducteurs de signal et les voies de transcription (STAT) 4 et 5, se produit en réponse à la signalisation IL-12 et IL-2 + IL-15, respectivement[59]. Les récepteurs de la cellule NK lancent également des voies de transcription inflammatoire lors de l'activation. Ceux-ci comprennent l'hétérodimère c-Fos et c-Jun, AP-1, le facteur nucléaire kappa-amplificateur de chaîne légère des cellules B activées (NF-κB) et le facteur nucléaire des cellules T activées[60],[61] qui se lient aux régions promotrices et favorisent la transcription du gène de la cytokine inflammatoire[62],[63]
Les cellules NK sécréteraient également des protéines FasR afin d'induire la mort de cellules tumorales[64].
Les cellules NK dans la santé et la maladie
Cellules NK dans la maladie

Rôle des cellules NK dans l'infection virale ou bactérienne
Il a été montré que les cellules NK pouvaient être activées par une interaction directe entre les peptides viraux (présents en surface du virus) et des récepteurs membranaires retrouvés sur la cellule NK. Pour exemple, le récepteur NKp46 avec l’hémagglutinine du virus influenza (virus de la grippe) ou l’hémagglutinine-neuramidase du virus Sendai[65]. De la même manière, l’interaction du récepteur NKp44 avec la glycoprotéine E du DENV pourrait activer les cellules NK[66].
Bien avant les cellules T CD8, les cellules NK sont activées dans la réponse immune antivirale. Dans les quelques heures suivant l'infection virale, un pic de cytokines telles que l'IL-12 et les interférons de type I est retrouvé dans la sérologie[67]. En effet, la présence d'interférons est spécifique d'une infection virale : ce sont des cytokines antivirales synthétisables par divers types cellulaires dans l'organisme. Les IFN I induisent l'activation de voies de signalisation permettant en amont la synthèse de protéines antivirales. De plus, les IFN I ont un rôle important dans l'activation des cellules NK (notamment en stimulant leur synthèse de protéines cytotoxiques) et leur prolifération.
Les cellules NK sont aussi activées par l’IL-12 et le TNF-α produits par les macrophages au début de nombreuses infections et qui induisent une production d'autres cytokines, majoritairement l’IFN-γ[68]. Cette activation des cellules NK durant le début de l’infection virale permet de bloquer ou tout au moins de contenir l'infection, pendant qu'une réponse immune adaptative, plus spécifique au type de virus, se met en place.
Les cellules dendritiques sont aussi connues pour activer les cellules NK dans certains cas. Elles sécrètent l'IL-12, l'IL-18 et l'IFN-I qui activent les fonctions des cellules NK[69].
Une fois activée, la cellule NK agit en trois phases majeures que l'on peut découper de la manière qui suit :
1. la production de cytokines ;
2. le relargage de granules cytotoxiques (granzymes et perforines, essentiellement) ;
3. la lyse de la cellule cible.
Rôle des cellules NK dans la lymphohistiocytose hémophagocytaire
La biologie des cellules NK est d'un intérêt particulier dans la lymphohistiocytose hémophagocytaire primaire (pHLH pour les anglophones) car tous les défauts génétiques associés à ce trouble entraînent une diminution de la capacité cytotoxique des cellules NK et des lymphocytes T ; des tests de destruction des cellules NK sont utilisés cliniquement pour le diagnostic de HLH. L'importance des altérations de la fonction des cellules NK semble liée à la pathogenèse de la lymphohistiocytose hémophagocytaire[70].
Traitement et cellule NK
Les lymphocytes NK CAR sont des lymphocytes NK génétiquement modifiés pour cibler des antigènes spécifiques situés à la surface des cellules cancéreuses[71]. C'est un axe de recherche en immunothérapie. Cette approche semble présenter moins d'effets secondaires indésirables que le traitement par lymphocytes T CAR[72],[73],[74].
Pathologies en rapport avec les cellules NK
Déficits en lymphocytes NK
Certaines maladies comportent une diminution du nombre global de ces cellules. D'autres ont une déficience de leur fonction tout en conservant un nombre normal. Toutes entraînent un déficit immunitaire.
La mutation du gène MCM4 entraîne un syndrome dont l'un des éléments est un déficit en cellules NK[75]. Celle des gènes GATA2[76] et RTEL1[77] comporte également une lymphopénie concernant ces cellules. D'autres mutations, par exemple concernant le CD16[78] ou l'IRF8[79] , sont responsables d'un déficit fonctionnel.
Une étude japonaise démontre que de brefs séjours en forêt permettent d'augmenter très sensiblement le nombre de lymphocytes NK[80].
Références
- http://acces.ens-lyon.fr/biotic/biomol/enjeux/TGS/html/nk.htm#
- Kiessling R, Klein E, Pross H, Wigzell H. (1975a). 'Natural' killer cells in the mouse. II. Cytotoxic cells with specificity for mouse Moloney leukemia cells. Characteristics of the killer cell. Eur J Immunol 5: 117–121
- (en) Janeway CA, Jr. et al., Immunobiology., Garland Science, , 6e éd. (ISBN 0-443-07310-4)
- (en) W. Rosenau et HD. Moon, « Lysis of homologous cells by sensitized lymphocytes in tissue culture », J Natl Cancer Inst, no 27, , p.471–83.
- Smith HJ. Antigenicity of carcinogen-induced and spontaneous tumours in inbred mice. Br J Cancer (1966) 20:831–7. doi:10.1038/bjc.1966.95
- (en) Robert K. Oldham, « Natural killer cells: Artifact to reality:: An odyssey in biology », CANCER AND METASTASIS REVIEW, vol. 2, no 4, , p. 323–336 (ISSN 0167-7659 et 1573-7233, DOI 10.1007/BF00048565, lire en ligne, consulté le )
- Oldham RK, Siwarski D, McCoy JL, Plata EJ, Herberman RB. Evaluation of a cell-mediated cytotoxicity assay utilizing 125 iododeoxyuridine-labeled tissue-culture target cells. Natl Cancer Inst Monogr (1973) 37:49–58.
- Pross HF, Jondal M. Cytotoxic lymphocytes from normal donors. A functional marker of human non-T lymphocytes. Clin Exp Immunol (1975) 21:226–35.
- Paolo Carrega et Guido Ferlazzo, « Natural killer cell distribution and trafficking in human tissues », Frontiers in Immunology, vol. 3, (ISSN 1664-3224, PMID 23230434, PMCID PMC3515878, DOI 10.3389/fimmu.2012.00347, lire en ligne, consulté le )
- (en) R. Kiessling, Eva Klein, H. Pross et H. Wigzell, « „Natural” killer cells in the mouse. II. Cytotoxic cells with specificity for mouse Moloney leukemia cells. Characteristics of the killer cell », European Journal of Immunology, vol. 5, no 2, , p. 117–121 (DOI 10.1002/eji.1830050209, lire en ligne, consulté le )
- (en) R. Kiessling, Eva Klein et H. Wigzell, « „Natural” killer cells in the mouse. I. Cytotoxic cells with specificity for mouse Moloney leukemia cells. Specificity and distribution according to genotype », European Journal of Immunology, vol. 5, no 2, , p. 112–117 (DOI 10.1002/eji.1830050208, lire en ligne, consulté le )
- (en) H J Smith, « Antigenicity of carcinogen-induced and spontaneous tumours in inbred mice. », British Journal of Cancer, vol. 20, no 4, , p. 831–837 (ISSN 0007-0920 et 1532-1827, PMID 5964614, PMCID PMC2008147, DOI 10.1038/bjc.1966.95, lire en ligne, consulté le )
- Herberman RB, Nunn ME, Holden HT, Lavrin DH. Natural cytotoxic reactivity of mouse lymphoid cells against syngeneic and allogeneic tumors. II. Characterization of effector cells. Int J Cancer (1975) 16:230–9.
- (en) Ronald B. Herberman, Myrthel E. Nunn et David H. Lavrin, « Natural cytotoxic reactivity of mouse lymphoid cells against syngeneic and allogeneic tumors. I. Distribution of reactivity and specificity », International Journal of Cancer, vol. 16, no 2, , p. 216–229 (DOI 10.1002/ijc.2910160204, lire en ligne, consulté le )
- Steven D. Scoville, Aharon G. Freud et Michael A. Caligiuri, « Modeling Human Natural Killer Cell Development in the Era of Innate Lymphoid Cells », Frontiers in Immunology, vol. 8, (ISSN 1664-3224, PMID 28396671, PMCID PMC5366880, DOI 10.3389/fimmu.2017.00360, lire en ligne, consulté le )
- (en) Joseph C. Sun et Lewis L. Lanier, « NK cell development, homeostasis and function: parallels with CD8+ T cells », Nature Reviews Immunology, vol. 11, no 10, , p. 645–657 (ISSN 1474-1733 et 1474-1741, PMID 21869816, PMCID PMC4408539, DOI 10.1038/nri3044, lire en ligne, consulté le )
- (en) Nathalie Jacobs, Langers, Renoux et Thiry, « Natural killer cells: role in local tumor growth and metastasis », Biologics: Targets and Therapy, , p. 73 (ISSN 1177-5475, PMID 22532775, PMCID PMC3333822, DOI 10.2147/BTT.S23976, lire en ligne, consulté le )
- (en) L L Lanier, R Testi, J Bindl et J H Phillips, « Identity of Leu-19 (CD56) leukocyte differentiation antigen and neural cell adhesion molecule. », The Journal of Experimental Medicine, vol. 169, no 6, , p. 2233–2238 (ISSN 0022-1007 et 1540-9538, PMID 2471777, PMCID PMC2189344, DOI 10.1084/jem.169.6.2233, lire en ligne, consulté le )
- Lanier LL, Phillips JH, Hackett J Jr, Tutt M, Kumar V. Natural killer cells: definition of a cell type rather than a function. J Immunol (1986) 137:2735–9.
- (en) Motonari Kondo, Irving L. Weissman et Koichi Akashi, « Identification of Clonogenic Common Lymphoid Progenitors in Mouse Bone Marrow », Cell, vol. 91, no 5, , p. 661–672 (DOI 10.1016/S0092-8674(00)80453-5, lire en ligne, consulté le )
- (en) Jianhua Yu, Aharon G. Freud et Michael A. Caligiuri, « Location and cellular stages of natural killer cell development », Trends in Immunology, vol. 34, no 12, , p. 573–582 (PMID 24055329, PMCID PMC3852183, DOI 10.1016/j.it.2013.07.005, lire en ligne, consulté le )
- (en) Massimo Vitale, Michela Falco, Roberta Castriconi et Silvia Parolini, « Identification of NKp80, a novel triggering molecule expressed by human NK cells », European Journal of Immunology, vol. 31, no 1, , p. 233–242 (ISSN 1521-4141, DOI 10.1002/1521-4141(200101)31:13.0.CO;2-4, lire en ligne, consulté le )
- (en) Aharon G. Freud, Karen A. Keller, Steven D. Scoville et Bethany L. Mundy-Bosse, « NKp80 Defines a Critical Step during Human Natural Killer Cell Development », Cell Reports, vol. 16, no 2, , p. 379–391 (PMID 27373165, PMCID PMC4970225, DOI 10.1016/j.celrep.2016.05.095, lire en ligne, consulté le )
- Cooper MA1, Fehniger TA, Caligiuri MA. The biology of human natural killer-cell subsets. Trends Immunol. 2001 Nov;22(11):633-40
- (en) Megan A. Cooper, Todd A. Fehniger, Sarah C. Turner et Kenneth S. Chen, « Human natural killer cells: a unique innate immunoregulatory role for the CD56bright subset », Blood, vol. 97, no 10, , p. 3146–3151 (ISSN 1528-0020 et 0006-4971, DOI 10.1182/blood.V97.10.3146, lire en ligne, consulté le )
- (en) Roland Jacobs, Gabriele Hintzen, Almut Kemper et Katrin Beul, « CD56bright cells differ in their KIR repertoire and cytotoxic features from CD56dim NK cells », European Journal of Immunology, vol. 31, no 10, , p. 3121–3126 (ISSN 1521-4141, DOI 10.1002/1521-4141(2001010)31:103.0.CO;2-4, lire en ligne, consulté le )
- (en) Eleftheria E. Rosmaraki, Iyadh Douagi, Claude Roth et Francesco Colucci, « Identification of committed NK cell progenitors in adult murine bone marrow », European Journal of Immunology, vol. 31, no 6, , p. 1900–1909 (ISSN 1521-4141, DOI 10.1002/1521-4141(200106)31:63.0.CO;2-M, lire en ligne, consulté le )
- (en) Raffaella Meazza, Bruno Azzarone, Anna Maria Orengo et Silvano Ferrini, « Role of Common-Gamma Chain Cytokines in NK Cell Development and Function: Perspectives for Immunotherapy », Journal of Biomedicine and Biotechnology, vol. 2011, , p. 1–16 (ISSN 1110-7243 et 1110-7251, PMID 21716670, PMCID PMC3118299, DOI 10.1155/2011/861920, lire en ligne, consulté le )
- (en) Martin J. Boulanger et K.Christopher Garcia, « Shared Cytokine Signaling Receptors: Structural Insights from the Gp130 System », dans Advances in Protein Chemistry, vol. 68, Elsevier, (ISBN 978-0-12-034268-6, DOI 10.1016/s0065-3233(04)68004-1, lire en ligne), p. 107–146
- (en) Kamran Ghoreschi, Arian Laurence et John J. O’Shea, « Janus kinases in immune cell signaling », Immunological Reviews, vol. 228, no 1, , p. 273–287 (PMID 19290934, PMCID PMC2782696, DOI 10.1111/j.1600-065X.2008.00754.x, lire en ligne, consulté le )
- (en) Kotaro Suzuki, Hiroshi Nakajima, Yasushi Saito et Takashi Saito, « Janus kinase 3 (Jak3) is essential for common cytokine receptor γ chain (γc)-dependent signaling: comparative analysis of γc, Jak3, and γc and Jak3 double-deficient mice », International Immunology, vol. 12, no 2, , p. 123–132 (ISSN 1460-2377 et 0953-8178, DOI 10.1093/intimm/12.2.123, lire en ligne, consulté le )
- (en) Geoffrey A Smith, Kenji Uchida, Arthur Weiss et Jack Taunton, « Essential biphasic role for JAK3 catalytic activity in IL-2 receptor signaling », Nature Chemical Biology, vol. 12, no 5, , p. 373–379 (ISSN 1552-4450 et 1552-4469, PMID 27018889, PMCID PMC4837022, DOI 10.1038/nchembio.2056, lire en ligne, consulté le )
- (en) Georg Gasteiger, Saskia Hemmers, Matthew A. Firth et Audrey Le Floc’h, « IL-2–dependent tuning of NK cell sensitivity for target cells is controlled by regulatory T cells », The Journal of Experimental Medicine, vol. 210, no 6, , p. 1167–1178 (ISSN 1540-9538 et 0022-1007, PMID 23650441, PMCID PMC3674692, DOI 10.1084/jem.20122462, lire en ligne, consulté le )
- (en) Zeguang Wu, Giada Frascaroli, Carina Bayer et Tatjana Schmal, « Interleukin-2 from Adaptive T Cells Enhances Natural Killer Cell Activity against Human Cytomegalovirus-Infected Macrophages », Journal of Virology, vol. 89, no 12, , p. 6435–6441 (ISSN 0022-538X et 1098-5514, PMID 25855747, PMCID PMC4474314, DOI 10.1128/JVI.00435-15, lire en ligne, consulté le )
- (en) Sigrid Dubois, Jennifer Mariner, Thomas A Waldmann et Yutaka Tagaya, « IL-15Rα Recycles and Presents IL-15 In trans to Neighboring Cells », Immunity, vol. 17, no 5, , p. 537–547 (DOI 10.1016/S1074-7613(02)00429-6, lire en ligne, consulté le )
- (en) Erwan Mortier, Jérôme Bernard, Ariane Plet et Yannick Jacques, « Natural, Proteolytic Release of a Soluble Form of Human IL-15 Receptor α-Chain That Behaves as a Specific, High Affinity IL-15 Antagonist », The Journal of Immunology, vol. 173, no 3, , p. 1681–1688 (ISSN 0022-1767 et 1550-6606, DOI 10.4049/jimmunol.173.3.1681, lire en ligne, consulté le )
- (en) Kresten Skak, Klaus Stensgaard Frederiksen et Dorthe Lundsgaard, « Interleukin-21 activates human natural killer cells and modulates their surface receptor expression », Immunology, vol. 123, no 4, , p. 575–583 (ISSN 0019-2805 et 1365-2567, PMID 18005035, PMCID PMC2433320, DOI 10.1111/j.1365-2567.2007.02730.x, lire en ligne, consulté le )
- Kamalakannan Rajasekaran, Matthew J. Riese, Sridhar Rao et Li Wang, « Signaling in Effector Lymphocytes: Insights toward Safer Immunotherapy », Frontiers in Immunology, vol. 7, (ISSN 1664-3224, PMID 27242783, PMCID PMC4863891, DOI 10.3389/fimmu.2016.00176, lire en ligne, consulté le )
- (en) Lewis L Lanier, « Up on the tightrope: natural killer cell activation and inhibition », Nature Immunology, vol. 9, no 5, , p. 495–502 (ISSN 1529-2908 et 1529-2916, PMID 18425106, PMCID PMC2669298, DOI 10.1038/ni1581, lire en ligne, consulté le )
- (en) Hisashi Arase, Tadahiro Suenaga, Noriko Arase et Yoshimitsu Kimura, « Negative Regulation of Expression and Function of FcγRIII by CD3ζ in Murine NK Cells », The Journal of Immunology, vol. 166, no 1, , p. 21–25 (ISSN 0022-1767 et 1550-6606, DOI 10.4049/jimmunol.166.1.21, lire en ligne, consulté le )
- Raffaella Augugliaro, Silvia Parolini, Roberta Castriconi et Emanuela Marcenaro, « Selective cross-talk among natural cytotoxicity receptors in human natural killer cells », European Journal of Immunology, vol. 33, no 5, , p. 1235–1241 (DOI 10.1002/eji.200323896, lire en ligne, consulté le )
- (en) Rebecca M. May, Mariko Okumura, Chin-Jung Hsu et Hamid Bassiri, « Murine natural killer immunoreceptors use distinct proximal signaling complexes to direct cell function », Blood, vol. 121, no 16, , p. 3135–3146 (ISSN 0006-4971 et 1528-0020, PMID 23407547, PMCID PMC3630829, DOI 10.1182/blood-2012-12-474361, lire en ligne, consulté le )
- (en) David B. Rosen, Manabu Araki, Jessica A. Hamerman et Taian Chen, « A Structural Basis for the Association of DAP12 with Mouse, but Not Human, NKG2D », The Journal of Immunology, vol. 173, no 4, , p. 2470–2478 (ISSN 0022-1767 et 1550-6606, DOI 10.4049/jimmunol.173.4.2470, lire en ligne, consulté le )
- (en) Alejandro López-Soto, Leticia Huergo-Zapico, Andrea Acebes-Huerta et Mónica Villa-Alvarez, « NKG2D signaling in cancer immunosurveillance: NKG2D signaling », International Journal of Cancer, vol. 136, no 8, , p. 1741–1750 (DOI 10.1002/ijc.28775, lire en ligne, consulté le )
- Smith KM, Wu J, Bakker AB, Phillips JH, Lanier LL. Ly-49D and Ly-49H associate with mouse DAP12 and form activating receptors. J Immunol (1998) 161:7–10.
- (en) Mark T. Orr, Joseph C. Sun, David G.T. Hesslein et Hisashi Arase, « Ly49H signaling through DAP10 is essential for optimal natural killer cell responses to mouse cytomegalovirus infection », Journal of Experimental Medicine, vol. 206, no 4, , p. 807–817 (ISSN 1540-9538 et 0022-1007, PMID 19332875, PMCID PMC2715124, DOI 10.1084/jem.20090168, lire en ligne, consulté le )
- Yuxia Zhang et Bing Huang, « The Development and Diversity of ILCs, NK Cells and Their Relevance in Health and Diseases », dans Regulation of Inflammatory Signaling in Health and Disease, vol. 1024, Springer Singapore, (ISBN 978-981-10-5986-5, DOI 10.1007/978-981-10-5987-2_11, lire en ligne), p. 225–244
- Helena Stabile, Cinzia Fionda, Angela Gismondi et Angela Santoni, « Role of Distinct Natural Killer Cell Subsets in Anticancer Response », Frontiers in Immunology, vol. 8, (ISSN 1664-3224, PMID 28360915, PMCID PMC5352654, DOI 10.3389/fimmu.2017.00293, lire en ligne, consulté le )
- (en) Mark J. Smyth, Erika Cretney, Janice M. Kelly et Jennifer A. Westwood, « Activation of NK cell cytotoxicity », Molecular Immunology, vol. 42, no 4, , p. 501–510 (DOI 10.1016/j.molimm.2004.07.034, lire en ligne, consulté le )
- (en) Cyril Fauriat, Eric O. Long, Hans-Gustaf Ljunggren et Yenan T. Bryceson, « Regulation of human NK-cell cytokine and chemokine production by target cell recognition », Blood, vol. 115, no 11, , p. 2167–2176 (ISSN 0006-4971 et 1528-0020, PMID 19965656, PMCID PMC2844017, DOI 10.1182/blood-2009-08-238469, lire en ligne, consulté le )
- (en) Bailey E. Freeman, Hans-Peter Raué, Ann B. Hill et Mark K. Slifka, « Cytokine-Mediated Activation of NK Cells during Viral Infection », Journal of Virology, vol. 89, no 15, , p. 7922–7931 (ISSN 0022-538X et 1098-5514, PMID 25995253, PMCID PMC4505636, DOI 10.1128/JVI.00199-15, lire en ligne, consulté le )
- (en) Esther Reefman, Jason G. Kay, Stephanie M. Wood et Carolin Offenhäuser, « Cytokine Secretion Is Distinct from Secretion of Cytotoxic Granules in NK Cells », The Journal of Immunology, vol. 184, no 9, , p. 4852–4862 (ISSN 0022-1767 et 1550-6606, DOI 10.4049/jimmunol.0803954, lire en ligne, consulté le )
- (en) Kamalakannan Rajasekaran, Pawan Kumar, Kristina M Schuldt et Erik J Peterson, « Signaling by Fyn-ADAP via the Carma1–Bcl-10–MAP3K7 signalosome exclusively regulates inflammatory cytokine production in NK cells », Nature Immunology, vol. 14, no 11, , p. 1127–1136 (ISSN 1529-2908 et 1529-2916, PMID 24036998, PMCID PMC3855032, DOI 10.1038/ni.2708, lire en ligne, consulté le )
- (en) E. Vivier, D. H. Raulet, A. Moretta et M. A. Caligiuri, « Innate or Adaptive Immunity? The Example of Natural Killer Cells », Science, vol. 331, no 6013, , p. 44–49 (ISSN 0036-8075 et 1095-9203, PMID 21212348, PMCID PMC3089969, DOI 10.1126/science.1198687, lire en ligne, consulté le )
- (en) Kevin D. Cook, Stephen N. Waggoner et Jason K. Whitmire, « NK Cells and Their Ability to Modulate T Cells during Virus Infections », Critical Reviews in Immunology, vol. 34, no 5, , p. 359–388 (ISSN 1040-8401, DOI 10.1615/CritRevImmunol.2014010604, lire en ligne, consulté le )
- Blanchard DK, Michelini-Norris MB, Djeu JY. Production of granulocyte-macrophage colony-stimulating factor by large granular lymphocytes stimulated with Candida albicans: role in activation of human neutrophil function. Blood (1991) 77:2259–65.
- (en) G. Bosch, F. Preijers, A. Vreugdenhil et J. Hendriks, « Granulocyte-macrophage colony-stimulating factor (GM-CSF) counteracts the inhibiting effect of monocytes on natural killer (NK) cells », Clinical & Experimental Immunology, vol. 101, no 3, , p. 515–520 (PMID 7664499, PMCID PMC1553231, DOI 10.1111/j.1365-2249.1995.tb03143.x, lire en ligne, consulté le )
- (en) Thierry Walzer, Marc Dalod, Scott H. Robbins et Laurence Zitvogel, « Natural-killer cells and dendritic cells: “l'union fait la force” », Blood, vol. 106, no 7, , p. 2252–2258 (ISSN 0006-4971 et 1528-0020, DOI 10.1182/blood-2005-03-1154, lire en ligne, consulté le )
- Dagmar Gotthardt et Veronika Sexl, « STATs in NK-Cells: The Good, the Bad, and the Ugly », Frontiers in Immunology, vol. 7, (ISSN 1664-3224, PMID 28149296, PMCID PMC5241313, DOI 10.3389/fimmu.2016.00694, lire en ligne, consulté le )
- (en) Cristina M. Tato, Alejandro Villarino, Jorge H. Caamaño et Mark Boothby, « Inhibition of NF-κB Activity in T and NK Cells Results in Defective Effector Cell Expansion and Production of IFN-γ Required for Resistance to Toxoplasma gondii », The Journal of Immunology, vol. 170, no 6, , p. 3139–3146 (ISSN 0022-1767 et 1550-6606, DOI 10.4049/jimmunol.170.6.3139, lire en ligne, consulté le )
- (en) Ilaria Tassi, Marina Cella, Rachel Presti et Angela Colucci, « NK cell–activating receptors require PKC-θ for sustained signaling, transcriptional activation, and IFN-γ secretion », Blood, vol. 112, no 10, , p. 4109–4116 (ISSN 0006-4971 et 1528-0020, PMID 18784374, PMCID PMC2581989, DOI 10.1182/blood-2008-02-139527, lire en ligne, consulté le )
- (en) Antonio Sica, Linda Dorman, Vincenzo Viggiano et Marco Cippitelli, « Interaction of NF-κB and NFAT with the Interferon-γ Promoter », Journal of Biological Chemistry, vol. 272, no 48, , p. 30412–30420 (ISSN 0021-9258 et 1083-351X, DOI 10.1074/jbc.272.48.30412, lire en ligne, consulté le )
- Zhang F, Wang DZ, Boothby M, Penix L, Flavell RA, Aune TM. Regulation of the activity of IFN-gamma promoter elements during Th cell differentiation. J Immunol (1998) 161:6105–12.
- Zamai L, Ahmad M, Bennett IM, Azzoni L, Alnemri ES, Perussia B. (1998). Natural killer (NK) cell-mediated cytotoxicity: differential use of TRAIL and Fas ligand by immature and mature primary human NK cells. J Exp Med 188: 2375–2380
- d'après Mandelboim O et al., 2001
- Hershkovitz O et al.2009
- Samuel CE et al. 2001
- Nguyen KB et al, 2002
- Banchereau J et al., 1998
- (en) Anthony R. French et Megan A. Cooper, « Immunology of Cytokine Storm Syndromes: Natural Killer Cells », dans Cytokine Storm Syndrome, Springer International Publishing, (ISBN 978-3-030-22094-5, DOI 10.1007/978-3-030-22094-5_10, lire en ligne), p. 163–181
- La Recherche n° 542, décembre 2018, p. 46.
- Mitch Leslie New cancer-fighting cells enter trials, 2018
- Rohtesh S. Mehta, Katayoun Rezvani, Chimeric Antigen Receptor Expressing Natural Killer Cells for the Immunotherapy of Cancer, 2018
- Li Y, Hermanson DL, Moriarity BS, Kaufman DS, Human iPSC-Derived Natural Killer Cells Engineered with Chimeric Antigen Receptors Enhance Anti-tumor Activity, 2018
- Gineau L, Cognet C, Kara N et al. Partial MCM4 deficiency in patients with growth retardation, adrenal insufficiency, and natural killer cell deficiency, J Clin Invest, 2012;122:821–832
- Mace EM, Hsu AP, Monaco-Shawver L et al. Mutations in GATA2 cause human NK cell deficiency with specific loss of the CD56(bright) subset, Blood, 2013;121:2669–2677
- Hanna S, Béziat V, Jouanguy E, Casanova JL, Etzioni A, A homozygous mutation of RTEL1 in a child presenting with an apparently isolated natural killer cell deficiency, J Allergy Clin Immunol, 2015;136:1113–1114
- Grier JT, Forbes LR, Monaco-Shawver L et al. Human immunodeficiency-causing mutation defines CD16 in spontaneous NK cell cytotoxicity, J Clin Invest, 2012;122:3769–3780
- Mace EM, Bigley V, Gunesch JT et al. Biallelic mutations in IRF8 impair human NK cell maturation and function, J Clin Investig, 2017;127:306-320
- (en) Q. Li et al., « Visiting a Forest, but Not a City, Increases Human Natural Killer Activity and Expression of Anti-Cancer Proteins », International Journal of Immunopathology and Pharmacology, (DOI 10.1177/039463200802100113, lire en ligne, consulté le )
Voir aussi
Bibliographie
- Richard A. Goldsby, Thomas J. Kindt, Barbara A. Osborne et Serge Weinman. Immunologie : le cours de Janis Kuby. Dunod, Paris, 2003. (ISBN 2-10-007396-6)
Article connexe
 Portail de la biologie
Portail de la biologie  Portail de la biologie cellulaire et moléculaire
Portail de la biologie cellulaire et moléculaire  Portail de l’hématologie
Portail de l’hématologie  Portail de la zoologie
Portail de la zoologie  Portail de la médecine
Portail de la médecine