Neuropeptide
Un neuropeptide est un type d'hormone, plus précisément un peptide sécrété par un neurone et ayant essentiellement une fonction de neuromodulateur. Ce sont des polypeptides ou de petites protéines formés de quelques acides aminés jusqu'à plusieurs centaines (191 pour l'hormone de croissance).
Ils se lient généralement à un récepteur couplé à une protéine G capable, lorsqu'elle est activée, de déclencher une chaîne de réactions mettant en œuvre plusieurs effecteurs pouvant moduler finement la transmission du message. La transmission de l'information est plus lente que via un récepteur-canal qui, lui, peut rapidement déclencher un potentiel d'action postsynaptique (comme l'acétylcholine ou le glutamate) ou l'inhiber (GABA).
Plus de cent neuropeptides ont été décrits. Ils interviennent dans la régulation du sommeil, de l'éveil, de l'émotion, de la récompense, de la douleur, de l'apprentissage et de la mémoire[1]. Leur étude est relativement récente et beaucoup de choses restent à découvrir à leur égard.
Famille des neuropeptides
Plus d'une centaine de neurotransmetteurs neuropeptidiques ont été décrits. Voici les principaux classés par Nestler et al.[1] :
| Famille de la calcitonine | Tachykinine |
| PRGC (peptide relié au gène de la calcitonine) |
Neurokinine A Neurokinine B Neuropeptide K Substance P |
| Hormones libératrices/inhibitrices de l'hypothalamus | |
| Corticolibérine CRH (corticotropin-releasing hormone) GnRH (Gonadotropin releasing hormone) gonadolibérine GHRH (growth hormone-releasing hormone), somatolibérine Somatostatine TRH thyrotrophin-releasing hormone, hormone thyréotrope | |
| Hormones hypothalamiques | Famille VIP-glucagon |
| Ocytocine Vasopressine |
GLP-1 glucagon-like peptide PACAP pitutary adenylate cyclase-activating peptide VIP vasoactive intestinal polypeptide, peptide vasoactif intestinal |
| Famille du neuropeptide Y | Peptides opioïdes |
| Neuropeptide Y (NPY) PYY neuropeptide YY PP polypeptide pancréatique |
dynorphine leu-enképhaline met-enképhaline |
| Autres peptides | |
| ARP agouti-related peptide bradykinine cholécystokinine galanine |
MCH melanin-concentrating hormone, Hormone de mélano-concentration neurotensine orexine orphanine FQ |
Localisation

Les neuropeptides se trouvent dans le système nerveux central et périphérique, système sympathique et parasympathique inclus.
Outre leur fonction de neuromodulateur, les neuropeptides peuvent agir comme des hormones. C'est par exemple le cas de l'ocytocine et de la vasopressine, deux polypeptides qui sont synthétisés par les neurones de l'hypothalamus et libérés dans la circulation sanguine au niveau de l'hypophyse. Cette ocytocine circulante intervient comme hormone pour stimuler la contraction des muscles utérins lors de l’accouchement ou pour stimuler la contraction des glandes mammaires pour éjecter le lait lors de la tétée.
L'ocytocine ne pouvant franchir la barrière hémato-encéphalique, comment se fait-il qu'on la retrouve dans le cerveau ? Pour pouvoir jouer le rôle de neurotransmetteur, l'ocytocine doit être produite par les neurones hypothalamiques et être libérée dans des synapses à l'extrémité de leurs axones. L'ocytocine se lie alors à des récepteurs qui sont exprimés par des neurones de l'amygdale, l'hypothalamus ventromédial, du septum, du noyau accumbens et du tronc cérébral.
Synthèse des neuropeptides
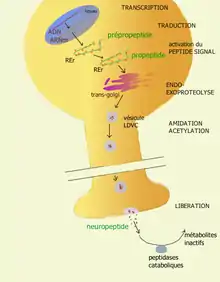
À la différence des neurotransmetteurs à petites molécules, la synthèse des neuropeptides, comme celle des protéines, part d'une structure codée dans l'ADN et se réalise à travers un long processus comportant plusieurs étapes susceptibles d'être régulées différemment suivant les tissus.
Le processus se déroule en plusieurs étapes :
- Transcription en ARNm d'une séquence d'ADN qui code un prépropeptide dans le noyau ;
- Ce transcrit primaire subit alors dans le noyau un processus de maturation au cours duquel des séquences (appelées introns) sont éliminées par excision et les séquences codantes sont raboutées (par épissage) pour former un ARNm mature. Lorsque les excisions et les épissages sont effectués de plusieurs manières, un même transcrit primaire peut donner plusieurs ARNm (par épissage alternatif) ;
- Ces séquences d'ARNm sont ensuite exportées dans le cytoplasme jusqu'aux ribosomes situés dans le réticulum endoplasmique rugueux où elles sont traduites en séquences d'acides aminés, nommées prépropeptides ;
- Cette séquence peptidique comporte en tête une séquence de signalisation (appelée le peptide signal) qui sert à diriger le peptide en formation vers la lumière du réticulum endoplasmique rugueux. Une fois sa fonction accomplie, cette séquence de tête est éliminée par excision pour donner un propeptide (sous l'action de la peptidase signal) ;
- Celui-ci est ensuite transféré dans l'appareil de Golgi et empaqueté dans une grosse vésicule de stockage spécial, la LDCV (large dense cored vesicle) où il subit des modifications post-traductionnelles comme des clivages, des amidations, sulfatations et autres modifications covalentes pour donner le peptide final actif.
La génération de plusieurs peptides actifs à partir d'un seul gène peut s'effectuer par deux grands processus :
Épissage alternatif
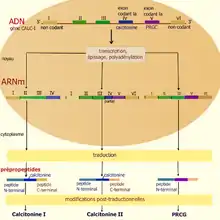
La génération de diversité par épissage alternatif peut s'illustrer avec la calcitonine et le peptide relié au gène calcitonine. Le gène CALC I donne par transcription un grand transcrit primaire comportant 6 exons, qui après épissage alternatif donne trois ARNm, deux spécifiques de la calcitonine et un du peptide relié au gène calcitonine (PRGC). Les ARNm mature de la calcitonine sont composés des exons I-II-III-IV et I-II-III-IV'-V-VI, celui du PRGC correspond aux exons I-II-III-V-VI. Les différents épissages possibles du transcrit primaire dépendent de facteurs tissu-spécifiques[2].
Après traduction, le premier ARNm donne un précurseur de la calcitonine, formé d'une région N-terminale (contenant le peptide signal) et d'une région C-terminale (CCP-I) encadrant la calcitonine. Le traitement post-traductionnel clivera ces peptides pour ne laisser que le peptide actif, la calcitonine. Le second précurseur de la calcitonine diffère du premier par son peptide C-terminal CCP-II. De même, l'ARNm du PRGC, donne par traduction le PRCG-I mature encadré par des peptides qui seront éliminés.
Chez l'homme, les plus fortes concentrations de calcitonine sont observées dans les cellules C parafolliculaires de la thyroïde mais aussi dans la parathyroïde et le thymus. Elle joue un rôle central dans le métabolisme du calcium et du phosphore. Le PRGC est produit dans les neurones du système nerveux central et périphérique et a pour fonction, entre autres, d'être un puissant vasodilatateur.
Modifications post-traductionnelles
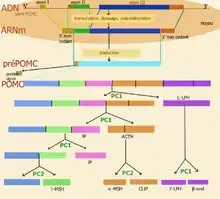
Les prépropeptides synthétisés dans le réticulum endoplasmique rugueux sont généralement plus grands que leurs produits actifs peptidiques et ils peuvent donner plusieurs neuropeptides[3]. Le traitement de post-traduction est une étape qui peut s'accomplir différemment suivant le tissu et donner une grande diversité de peptides. C'est le cas de la proopiomélanocortine POMC, un peptide qui, une fois traduit, va subir des transformations conduisant à plusieurs peptides plus petits comme l'ACTH, la mélanocortine (ou α-MSH) et la β-endorphine. Une fois le peptide signal éliminé, le propeptide subit une série de protéolyses effectuées par les prohormones convertases 1 et 2 (PC1 et PC2). La PC1 intervient au début du processus pour libérer l'ACTH, la β-endorphine et la γ-LPH. La convertase PC2 clive l'ACTH en CLIP et α-MSH ainsi que la γ-MSH de la portion N-terminale du propeptide[1].
La POMC est codée par un gène principalement exprimé de façon normale dans les cellules corticotropes de l'hypophyse antérieure, certaines cellules de l'hypothalamus (cellules du noyau arqué, et quelques neurones dorsomédians), et dans les mélanocytes. Les sites de clivage de la POMC varient d'un tissu à l'autre. Par exemple, dans les cellules corticotrophes de l'hypophyse antérieure, les clivages se font sur seulement 4 sites de clivage, tous de type Lys-Arg. Dans les mélanocytes ou l'hypothalamus, les clivages se produisent sur d'autres sites.
Stockage, libération et diffusion
Les neuropeptides sont stockés dans de grosses vésicules à cœur dense LDCV[4], (de 100 à 300 nm de diamètre) assemblées par l'appareil de Golgi puis transportées vers la synapse. Ces vésicules sont de taille supérieure aux petites vésicules claires (SSV small clear vesicle, de 50 nm) qui assurent le stockage des neurotransmetteurs à petites molécules (glutamate, GABA). Les neurotransmetteurs aminergiques (catécholamine, sérotonine, histamine) sont préférentiellement mais pas exclusivement stockées dans les grosses LDCV. Dans de nombreux cas, les grosses vésicules LDCV chargées de neuropeptides coexistent dans la même cellule avec les petites vésicules SSV chargées de petits neurotransmetteurs. Voici un tableau des colocalisations proposé par Nestler et coll. :
| Exemples de neuropeptides et de neurotransmetteurs à petites molécules colocalisés dans les mêmes terminaisons nerveuses (Nestler et coll[1] 2009) | ||
| Neuropeptides | Petites molécules | Sites de colocalisation |
| Neuropeptide Y | Noradrénaline | neurones du locus cerulus et préganglionnaires sympathiques |
| VIP | Acétylcholine | neurones préganglionnaires parasympathiques |
| CGRP | Acétylcholine | neurones moteur spinaux |
| Neurotensine, cholécystokininne | Dopamine | neurones de la substance noire |
| TRH, Substance P, enképhaline | Sérotonine | neurones des noyaux du raphé |
| Enképhaline | GABA | neurones du striatum se projetant sur le globus pallidus |
| Dynorphine, substance P | GABA | neurones du striatum se projetant sur la substance noire pars reticulata |
L'exocytose s'accomplit aussi de manière différente pour ces deux types de vésicules. L'exocytose des petites SSV s'opère rapidement en réponse à un seul potentiel d'action, alors que la libération des neuropeptides enfermés dans la grosse vésicule LDCV se fait lentement en réponse à une stimulation soutenue[4].
Les petites vésicules SSV contiennent des neurotransmetteurs capables d'activer des récepteurs-canaux ioniques alors que les grosses vésicules LDCV contiennent des neuropeptides, des hormones et des neurotransmetteurs qui activent des récepteurs couplés aux protéines G. C'est pourquoi l'exocytose des LDCV est avant tout associée avec la modulation de l'activité synaptique et ne peut pas induire par elle-même une activité synaptique propre.
Une autre différence concerne les sites de libération : les petites vésicules libèrent leurs neurotransmetteurs directement dans la fente synaptique, les grosses LDCV loin des synapses.
Une fois émises dans la synapse, les petites molécules de neurotransmetteurs disparaissent rapidement si elles n'ont pas pu se lier à un récepteur. Elles sont métabolisées par des enzymes ou recaptées très rapidement. C'est l'inverse pour les neuropeptides qui ne sont pas recaptés et ont le temps de diffuser loin de leur site de libération. Ils peuvent être quand même détruits par des peptidases.
Neuropeptide Y
Le neuropeptide Y (NPY) fait partie de la famille du même nom : les neuropeptides Y, qui comportent dans cette famille en plus du NPY, le polypeptide Y (PYY) et le polypeptide pancréatique (PP). Ce sont des neurotransmetteurs peptidiques qui interviennent dans la régulation de la prise de nourriture, des fonctions sexuelles, de la température corporelle, la régulation de la pression artérielle…
Définition
Le neuropeptide Y a été découvert par Tatemoto en 1982. Il est constitué de 36 acides aminés, ceux-ci restent très conservés à travers le règne animal : on observe une différence d'un seul acide aminé entre le NPY de la poule et celui de l'humain. On retrouve le NPY dans les neurones du système nerveux central et périphérique, dans le sang, ainsi que dans les plexus nerveux périvasculaires de nombreux organes. Il est localisé aussi bien dans le système sympathique que parasympathique. Le NPY dispose de nombreux récepteurs métabotropiques récepteurs métabotropiques, tels que Y1, 2, 3 4, 5 et 6. Au niveau cérébral, il est synthétisé dans le noyau arqué de l'hypothalamus.
Diminution de la thermogenèse
La thermogenèse est due à la vasoconstriction des vaisseaux au niveau de la peau, permettant ainsi à l'organisme de maintenir sa température corporelle lorsque les conditions le nécessitent (lorsque la température extérieure est très basse par exemple). Or le neuropeptide Y entraîne la vasoconstriction des vaisseaux, il joue donc un rôle dans la thermogenèse.
Développement de l'obésité
Celle-ci est généralement due à un excès de prise alimentaire. Or, on observe chez les sujets obèses, un taux élevé de NPY, s'accompagnant d'une augmentation de la prise alimentaire et d'une hypersécrétion d'insuline, malgré un degré d'adiposité élevé. Cet excès de NPY peut avoir deux origines : soit le tissu adipeux ne sécrète pas de leptine (inhibant la sécrétion de NPY), soit les récepteurs du noyau arqué sont insensibles à la leptine, à la suite par exemple d'une mutation de ces récepteurs. Mais le rôle du NPY dans l'obésité n'est pas seulement dû à l'augmentation des quantités ingérées. Il semble, en effet, qu'il engendre des modifications hormonales à l'origine d'une augmentation du taux d'insuline, d'acétylcholine, et corticostérone dans le sang, favorisant ainsi le stockage des nutriments et une résistance à l'insuline au niveau des muscles.
Grâce à une surrénalectomie, on a aussi pu établir un lien entre l’action centrale du NPY et les glucocorticoïdes. En effet, en l’absence de ces derniers, le NPY perdrait ses effets. Ainsi, on pourrait aller jusqu’à déduire que le stress, agissant sur les glucocorticoïdes, favoriserait la prise de poids.
Effets anxiolytique et sédatif
On a remarqué une augmentation de NPY, au niveau cérébral, après traitement aux antidépresseurs. On a aussi constaté un taux plus faible de NPY dans le cerveau et le liquide cérébrospinal de personnes suicidées, le NPY serait donc également un anxiolytique.
Le processus de mémorisation serait aussi facilité par le NPY grâce aux récepteurs Y2. Si un agoniste venait à être développé, il pourrait peut-être aider au traitement de la maladie d’Alzheimer.
Notes et références
- (en) Eric J. Nestler, Steven E. Hyman, Robert C. Malenka, Molecular neuropharmacology : A foundation for clinical neuroscience (2e édition), McGraw-Hill, 2001, 2009
- R. Cohen, K.L. Becker, A. Julienne, « Calcitonine et peptidesapparentés », EMC-endocrinologie, vol. 1, , p. 200-213
- Purves, Augustine, Fitzpatrick, Hall, LaMantia, McNamara, Williams, Neurosciences, De Boeck, 2005 (3e éd.), 811 p.
- Yongsoo Park, Kyong-Tai Kim, « Short-term plasticity of small synaptic vesicle (SSV) and large dense-core vesicle (LDCV) exocytosis », Cellular signalling, vol. 21, , p. 1465-1470
Voir aussi
Bibliographie
- J. S. Lacroix, 1995, Le neuropeptide Y sur orbite : Fonctions multiples et potentialité thérapeutique, Médecine & Hygiène, n° 2066, pp. 779 à 784
- J. S. Lacroix et B.L Mosimann et F Correia et A. Ricchetti, 1995, Neuropeptide Y et Rhume des foins, Médecine & Hygiène, n° 2091, pp. 2192 à 2194
- Purves, Augustine, Fitzpatrick, Hall, Lamantia, McNamara, Williams, Neurosciences, DeBoeck, Neurosciences & Cognition, pp. 155 à 160. (ISBN 28041-4797-5)
- Jean Claude Orsini, Neurobiologie du comportement alimentaire, collection neurobiologie, édition scientifique. (ISBN 2-84703-018-2) (BNF 39117021)
- I. Cusin et F. Rohner-Jeanrenaud, 1998, Boucle régulatrice entre le neuropeptide Y et la leptine et son altération chez le rongeur obèse, Médecine sciences, n° 8-9, vol. 14, pp. 907-913
Articles connexes
- Neuropeptide Y
- Secrétoneurine, un autre neuropeptide
Liens externes
 Portail des neurosciences
Portail des neurosciences  Portail de la biochimie
Portail de la biochimie  Portail de la médecine
Portail de la médecine