Origine africaine de l'Homme moderne
En paléoanthropologie, l'origine africaine de l'Homme moderne est la théorie la plus communément admise pour décrire l'origine des populations humaines actuelles. Cette théorie porte aussi l'appellation anglaise Out of Africa II (deuxième sortie d'Afrique), pour la différencier de Out of Africa I, expression qui renvoie aux migrations d'espèces antérieures du genre Homo hors d'Afrique au cours du Paléolithique inférieur et au début du Paléolithique moyen. Dans les milieux scientifiques elle porte aussi les noms d'« hypothèse d'une origine unique récente » (OUR), « hypothèse du remplacement » (HR), ou modèle de l'« origine africaine récente » (OAR).
« Out of Africa II » redirige ici. Pour les autres significations, voir Out of Africa (homonymie).


Homo sapiens, espèce de l'Homme moderne, est apparu en Afrique[1] et y aurait vécu depuis environ 300 000 ans, avant de quitter l'Afrique il y a entre 60 000 et 50 000 ans, se répandant sur les autres continents, ce que confirment les études génétiques[2], et supplantant les espèces humaines antérieures, comme l'Homme de Néandertal en Europe et l'Homme de Denisova en Asie, avec des épisodes d'hybridation limitée entre espèces.
Origine de l'homme moderne : monogénisme versus polygénisme
Avec l'essor de l'anthropologie au début du XIXe siècle, un débat virulent opposa les tenants du monogénisme comme Johann Friedrich Blumenbach et James Cowles Pritchard, pour qui les différentes races humaines sont des variétés partageant une ascendance commune, et ceux du polygénisme tels que Louis Agassiz et Josiah C. Nott, qui soutenaient que les races humaines sont des espèces distinctes ou se sont développées comme espèces distinctes par transmutation à partir de singes, sans avoir d'ancêtres communs.
Vers le milieu du XXe siècle, de nombreux anthropologues s'étaient ralliés à la théorie du monogénisme, mais les partisans du polygénisme, comme Carleton Coon, demeuraient influents. Ce dernier émit en 1962 l'hypothèse d'une évolution indépendante et séparée d’Homo erectus vers Homo sapiens sur chacun des cinq continents[3].
Le polygénisme a ensuite cédé la place dans les années 1980 à la théorie de l'origine multirégionale de l'homme moderne, une version intermédiaire dans laquelle les cinq branches d'Homo erectus échangent des gènes tout au long de leur évolution avant de parvenir au stade final Homo sapiens.
Le développement de la génétique des populations dans les années 1980 et 1990 a permis de montrer l'origine commune et récente de toute l'humanité actuelle, avec un enracinement sur le continent africain, replaçant le monogénisme comme théorie centrale et quasi-consensuelle de l'origine de l'Homme moderne.
Premiers Homo sapiens
Homo sapiens est apparu en Afrique il y a au moins 300 000 ans, d'après les fossiles les plus anciens connus à ce jour, trouvés à Djebel Irhoud, au Maroc, et publiés en 2017 par Jean-Jacques Hublin[4],[5]. Homo sapiens aurait colonisé tout le continent avant de migrer hors d'Afrique.
Dans sa publication de 2017, Jean-Jacques Hublin défend l'idée d'une émergence d'Homo sapiens à l'échelle de l'ensemble du continent africain, selon une sorte de modèle multirégional limité à l'Afrique[4]. Selon lui, l'arbre phylogénétique de l'humanité est un « arbre dont il manque de nombreuses branches » et la surreprésentation de certaines régions et périodes serait due avant tout à l'abondance de fossiles en Afrique orientale, région dont les conditions ont été propices à la conservation des ossements[6].
Le passage de l'industrie lithique acheuléenne aux industries dites de mode 3, qui s'est produit en Afrique à partir d'environ 400 000 ans avant le présent, témoignerait peut-être de la transition entre des formes humaines archaïques et les premiers Homo sapiens.
- Fossiles d'Homo sapiens découverts en Afrique et datés de plus de 100 000 ans (non exhaustif)
- Homme de Djebel Irhoud : 300 ka, Maroc[4],[5]
- Homme de Florisbad : 260 ka, Afrique du Sud[7]
- Homme d'Eliye Springs : 200 à 300 ka, Kenya[8]
- Homme de Ngaloba (LH 18) : au moins 200 ka, Tanzanie[8]
- Homme de Kibish : 195 ka, Éthiopie[9]
- Homme de Guomde : plus de 180 ka, Kenya[10]
- Homme de Herto : 157 ka, Éthiopie[11]
- Homme de Singa : au moins 133 ka, Soudan[10]
En 2019, une étude des paléoanthropologues français Aurélien Mounier (CNRS-MNHN) et argentine Marta Mirazón Larh (université de Cambridge), publiée dans la revue Nature, a recherché, parmi les plus anciens fossiles africains connus attribués à l'espèce Homo sapiens, lesquels préfiguraient le mieux la morphologie finalement acquise par l'Homme moderne. Ayant analysé de nombreux crânes d'hommes modernes, issus des différentes populations de la planète, cette étude propose une morphologie virtuelle du dernier ancêtre commun de l'humanité actuelle, et la compare, en morphométrie 3D, aux 5 crânes africains les plus complets datés d'au moins 200 000 ans : Irhoud 1 (Maroc), Florisbad (Afrique du Sud), Eliye Springs (Kenya), Omo Kibish 2 (Éthiopie), et LH 18 (Tanzanie). Le crâne de Florisbad est jugé le plus proche de notre ancêtre virtuel, devant celui d'Eliye Springs. Les trois autres crânes fossiles représenteraient des stades plus archaïques de l'espèce Homo sapiens[8],[12].
Dans une synthèse publiée en 2016, le paléoanthropologue anglais Christopher Brian Stringer, du Musée d'histoire naturelle de Londres, rappelle l'existence, dans plusieurs régions d'Afrique, de fossiles humains récents qui ne sont peut-être pas attribuables à l'espèce Homo sapiens. Les fossiles d'Iwo Eleru (14 ka, Nigeria), de Lukenya Hill (22 ka, Kenya), et du lac Eyasi (7 fragments de crâne, environ 110 ka, Tanzanie), pourraient témoigner de l'existence de populations reliques en Afrique, tout comme l'Homme de Kabwe (environ 250 ka, Zambie), un peu plus ancien, holotype de l'espèce Homo rhodesiensis[10].
Ces deux études soulignent la grande diversité des morphologies relevées sur les différents fossiles africains datés sur une période allant du Pléistocène moyen tardif jusqu'au Pléistocène supérieur, ce qui pourrait refléter la coexistence de populations ou d'espèces morphologiquement distinctes en Afrique tout au long de cette période[10]. Le modèle d'évolution applicable au Paléolithique moyen africain resterait alors un buissonnement évolutif, ce qui contredirait la théorie de l'origine multirégionale de l'homme moderne à l'échelle du continent, telle que proposée par Jean-Jacques Hublin[4], si on donne à cette théorie le sens qu'elle a à l'échelle de la planète.
Il est aussi possible d'envisager cette théorie sous une version intermédiaire, consistant à imaginer un chemin évolutif se déroulant successivement dans plusieurs régions d'Afrique, avec de possibles apports génétiques de populations plus archaïques le long du chemin. Dans cette vision, l'Homme moderne n'émergerait pas d'un foyer régional unique, mais par accrétion successive de caractères acquis à différents moments et dans différentes régions du continent. Ce chemin évolutif laisserait derrière lui différentes populations ou espèces humaines archaïques ayant coexisté un certain temps avec la lignée pionnière, avant de finalement s'éteindre peu avant ou durant le Pléistocène supérieur[13].
Premières sorties d'Afrique
Un fragment de calotte crânienne fossile, noté Apidima 1, fut découvert en 1978 dans la Grotte d'Apidima, située dans le sud du Péloponnèse, en Grèce. Grâce à l'imagerie virtuelle par tomodensitométrie, l'analyse du fossile en morphométrie 3D, soulignant notamment la rondeur de l'os occipital et l'absence de fosse sus-iniaque, a permis en 2019 de l'attribuer à Homo sapiens, avec une datation de 210 000 ans (datation par l'uranium-thorium). Cette datation reculerait de quelque 160 000 ans l'âge de l'arrivée des premiers Homo sapiens en Europe[14],[15].
Homo sapiens n'a pas pu à l'époque se maintenir en Europe où il a probablement été supplanté par l'Homme de Néandertal, mieux adapté au froid des cycles glaciaires successifs, avant que l'Homme de Cro-Magnon réussisse bien plus tard son implantation, à partir de 48 000 ans avant le présent.
Le second plus ancien fossile d'Homo sapiens trouvé hors d'Afrique, un demi-maxillaire avec ses huit dents, a été découvert en 2002 dans la grotte de Misliya, en Israël. Il a été daté en 2018 d'environ 185 000 ans[16]. Des fossiles d'Homo sapiens avaient été mis au jour en Israël dès les années 1930, dans la grotte d'Es Skhul, datés d'environ 118 000 ans, et dans la grotte de Qafzeh, datés d'environ 92 000 ans. Sur les deux sites, les individus exhumés ont bénéficié de sépultures[17].
Ces premiers Homo sapiens trouvés hors d'Afrique n'auraient pas contribué au patrimoine génétique de l'humanité actuelle. Ils auraient profité d'une période interglaciaire pour s'étendre en dehors de l'Afrique, avant que le retour ultérieur d'une phase glaciaire conduise peut-être à leur retrait d'Eurasie, au profit de Néandertaliens venus du Nord.
- Fossiles d'Homo sapiens découverts hors d'Afrique et datés de plus de 80 000 ans (non exhaustif)
- Apidima 1 : 210 ka, Grèce[15]
- Homme de Misliya : 185 ka, Israël[16]
- Homme de Skhul : 118 ka, Israël
- Homme de Zhiren : 110 ka, Chine
- Homme de Qafzeh : 92 ka, Israël
- Homme de Fuyan : au moins 80 ka, Chine
Néanmoins, les études de paléogénétique ont montré que les fondateurs de toutes les populations vivantes non africaines se sont étendues à partir de l'Afrique il y a environ entre 65 000 à 45 000 ans. Ce modèle de « dispersion tardive » avait été remis en question par la découverte d'humains anatomiquement modernes (AMH) isolés dans des grottes du sud de la Chine suggérée dès vers 120 000 ans. Une équipe de chercheurs a évalué l'âge des premiers fossiles d'AMH provenant de cinq grottes de cette région à l'aide d'une analyse ADN ancienne et d'une stratégie de datation géologique à plusieurs méthodes. Ceux-ci constatent que les fossiles étaient beaucoup plus jeunes que ce qui avait été suggéré précédemment, certains vestiges datant de l'Holocène en raison de l'histoire complexe des dépôts dans ces grottes subtropicales. Selon ces chercheurs, les preuves actuelles montrent que les AMH se sont installés dans le sud de la Chine dans le délai fixé par des données moléculaires inférieures entre 50 000 à 45 000 ans et pas plus tôt[18].
Émergence de l'Homme moderne
Il y a environ 100 000 ans, des indices de technologies et d'artisanat plus sophistiqués commencent à apparaitre en Afrique du Sud, à Pinnacle Point puis à Blombos. À partir d'environ 50 000 ans avant le présent, les outils lithiques s'affinent et les outils en os et en bois de cervidé se développent[19],[20].
Génétique des populations
Deux parties du génome humain d'accès plus facile ont longtemps été seules exploitées pour déchiffrer l'histoire de l'Homme moderne. L'un est l'ADN mitochondrial, et l'autre le chromosome Y. Ce sont les deux seules parties du génome qui ne se recombinent pas au moment de la fécondation. L'ADN mitochondrial et le chromosome Y sont transmis respectivement le long des lignées féminine et masculine. Sur la base de ces éléments, la totalité de l'humanité actuelle aurait hérité son ADN mitochondrial d'une femme qui vivait en Afrique il y a environ 160 000 ans[21],[22], dénommée Ève mitochondriale. Tous les hommes d'aujourd'hui ont hérité leurs chromosomes Y d'un homme qui vivait à une date controversée en Afrique. Cet homme est le plus récent ancêtre patrilinéaire commun.
ADN mitochondrial

Le premier lignage issu du plus récent ancêtre matrilinéaire commun, surnommé « Ève mitochondriale », est l'haplogroupe L0. Cet haplogroupe se trouve en proportions élevées parmi les San d'Afrique australe et les Sandawe de Tanzanie. On trouve l'haplogroupe L1 (en) chez les Pygmées Binga[23],[24] et les haplogroupes L2 à L6 dans les autres régions d'Afrique. Les macro haplogroupes M et N, qui sont les lignages du reste du monde en dehors de l'Afrique, descendent du groupe L3, présent principalement dans l'est et le nord-est de l'Afrique.
ADN chromosomique Y
Les mutations définissant les macro-haplogroupes CT (tous les haplogroupes Y sauf A et B) datent d'avant la migration hors d'Afrique, son descendant, le macro-groupe DE, étant confiné à l'Afrique. Les mutations qui distinguent l'haplogroupe C de tous les autres descendants de CT sont apparus il y a environ 140 000 ans[25].
L'haplogroupe F est apparu il y a environ 45 000 ans, en Afrique du Nord (dans ce cas il montrerait une seconde vague de migration hors d'Afrique) ou en Asie du Sud. Plus de 90 % des hommes nés ailleurs qu'en Afrique descendent en ligne masculine du premier porteur de l'haplogroupe F.
ADN autosomal
L'ADN autosomal est la partie non sexuée du génome, qui est transmise par les deux parents et se recombine à chaque génération. Elle représente l'essentiel du génome (22 paires de chromosomes sur 23). En , une analyse des données sur les génomes SNP de l'International HapMap Project (Phase II) et du CEPH Human Genome Diversity Panel samples a été publiée[26]. Ces modèles furent prélevés sur 1 138 individus qui n'étaient pas liés[26]. Avant cette analyse, les généticiens de population s'attendaient à trouver des différences importantes entre les groupes ethniques, avec des allèles partagés entre ces groupes mais différents ou n'existant pas dans les autres groupes[27]. Au lieu de cela, l'étude de 53 populations à partir des données des HapMap et CEPH a révélé que les groupes de population étudiés comprenaient seulement trois groupes génétiques : les Africains, les Eurasiens (qui incluent les natifs d'Europe et du Moyen-Orient, jusqu'à l'ouest de l'actuel Pakistan), et les Asiatiques de l'Est, qui incluent les natifs d'Asie orientale, d'Asie du Sud, des Amériques et d'Océanie[27]. L'étude a montré que la plupart des différences des groupes ethniques pouvaient être attribuées à la dérive génétique, et que les populations africaines modernes ont une plus grande diversité génétique que les deux autres groupes génétiques[27],[28]. Ce dernier point s'explique aisément quand on considère que la diversité génétique des Eurasiens n'a démarré qu'après la sortie d'Afrique, alors que celle des Africains a pu s'accumuler sur une période plus longue.
Dernière sortie d'Afrique
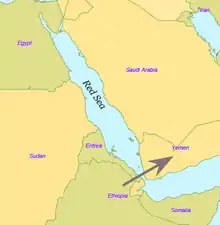
Il y a quelque 60 000 ans, un groupe de porteurs de l'haplogroupe mitochondrial L3 aurait migré d'Afrique orientale vers le Moyen-Orient.
Aujourd'hui le détroit de Bab-el-Mandeb, dans la mer Rouge, fait environ 20 kilomètres de largeur. Il y a 60 000 ans, lorsque le niveau de la mer était 70 mètres plus bas qu'aujourd'hui, il était beaucoup plus étroit. Bien que les détroits n'aient jamais été complètement fermés, il est possible qu'il y ait eu des iles qui pouvaient être atteintes avec de simples radeaux. Des tertres de coquilles datés de 125 000 ans ont été trouvés en Érythrée, indiquant que le régime alimentaire des premiers hommes incluait des fruits de mer obtenus par ramassage sur le rivage.
Certains chercheurs pensent que seules quelques personnes ont quitté l'Afrique dans le cadre d'une unique migration et qu'elles ont peuplé le reste du monde. Seul un petit groupe de près de 150 personnes aurait franchi la mer Rouge. C'est pourquoi, de tous les lignages présents en Afrique, seules les filles d'un seul lignage, L3, sont présentes hors d'Afrique. S'il y avait eu plusieurs migrations, on trouverait plus d'un lignage africain hors d'Afrique. Les filles du L3, les lignages M et N, sont peu fréquentes en Afrique subsaharienne (l'haplogroupe M1 est très ancien et diversifié en Afrique du Nord et en Afrique du Nord-Est) et semblent y être arrivées récemment. Une explication possible est que ces mutations se sont produites en Afrique de l'Est peu avant l'exode et, par effet fondateur, sont devenues les haplogroupes dominants après la sortie d'Afrique. Les mutations ont aussi pu se produire peu après la sortie d'Afrique.
D'autres chercheurs ont proposé un modèle de dispersion double selon lequel il y aurait eu deux sorties d'Afrique, dont l'une par la mer Rouge, qui se serait dirigée vers l'Inde en traversant les régions côtières (la route de la Côte), et qui serait représentée par l'Haplogroupe M. La seconde impliquerait un autre groupe, porteur de l'haplogroupe N, qui aurait suivi le Nil à partir de l'Afrique de l'Est, se dirigeant vers le nord et gagnant le Levant à travers le Sinaï. Puis, ce groupe se serait séparé dans plusieurs directions, certains allant en Europe et d'autres se dirigeant vers l'est, en Asie. Cette hypothèse tente d'expliquer pourquoi l'haplogroupe N est prédominant en Europe et pourquoi l'haplogroupe M y est absent. Cependant, une population de pionniers européens qui, initialement, possédait à la fois les haplogroupes M et N pourrait avoir perdu l'haplogroupe M en raison d'une dérive génétique accélérée par un éventuel goulet d'étranglement de population.
Les preuves d'une migration vers l'est par la côte d'Arabie ont pu être en partie détruites par la montée du niveau de la mer pendant l'Holocène[29],[30].
Une calotte crânienne fossile d'Homo sapiens, notée Manot 1, découverte en 2008 dans la grotte de Manot, en Galilée occidentale (Israël), publiée en 2015 avec une datation de 54 700 ans +/- 5 500 ans avant le présent, est le plus ancien fossile de morphologie moderne trouvé à ce jour au Levant. Sa datation semble indiquer un probable contact des hommes modernes avec les populations néandertaliennes contemporaines du Levant, représentées par les fossiles néandertaliens de la grotte de Kébara, datés d'environ 60 000 ans, et de la grotte d'Amud, datés d'environ 55 000 ans. Selon les études génétiques, cette période correspond à l'époque estimée de l'hybridation des Néandertaliens avec les hommes modernes, juste après leur sortie d'Afrique. Cette découverte tend à appuyer la théorie d'une dernière sortie d'Afrique par le Levant plutôt que par la mer Rouge[31].
L'hypothèse d'une sortie d'Afrique il y a 60 000 ans serait corroborée par l'analyse du crâne d'Hofmeyr[32], appartenant à un Homo sapiens d'Afrique du Sud, daté de 36 000 ans, et étonnamment semblable aux crânes d'Européens du Paléolithique supérieur[33]. Cette similitude suggère en effet que la population d’Afrique sub-saharienne dont l'Homme d'Hofmeyr était issu et les Européens du Paléolithique descendaient d'un ancêtre commun[32].
Références
- Joël Ignasse, « L'Homme moderne a bien une origine Africaine unique »
 , (consulté le )
, (consulté le ) - (en) Jérôme Goudet, François Balloux, Lori J Lawson Handley et Andrea Manica, « Going the distance: human population genetics in a clinal world. », revue scientifique, , p. 1 (lire en ligne )
- Jackson Jr., John P. (2001). "InWays Unacademical": The Reception of Carleton S. Coon's The Origin of Races
- (en) Jean-Jacques Hublin, Abdelouahed Ben-Ncer et al., « New fossils from Jebel Irhoud, Morocco and the pan-African origin of Homo sapiens », Nature, vol. 546, , p. 289-292 (DOI 10.1038/nature22336)
- (en) Daniel Richter et al., « The age of the hominin fossils from Jebel Irhoud, Morocco, and the origins of the Middle Stone Age », Nature, vol. 546, , p. 293-296 (DOI 10.1038/nature22335)
- Jean Jacques Hublin et Nicolas Martin, « Faut-il réécrire les débuts de l’histoire de l’Homme ? », France Culture, (lire en ligne, consulté le )
- (en) Rainer Grün, James S. Brink et al., « Direct dating of Florisbad hominid », Nature, vol. 382, no 6591, , p. 500–501 (DOI 10.1038/382500a0, lire en ligne)
- (en) Aurélien Mounier et Marta Mirazón Lahr, « Deciphering African late middle Pleistocene hominin diversity and the origin of our species », Nature, (lire en ligne)
- (en) Ian Mcdougall, F.H. Brown et John G. Fleagle, « Stratigraphic placement and age of modern humans from Kibish, Ethiopia », Nature, vol. 433, no 7027, , p. 733-736 (DOI 10.1038/nature03258)
- (en) Christopher Brian Stringer, « The origin and evolution of Homo sapiens », Philosophical Transactions of the Royal Society B - Biological Sciences, (lire en ligne)
- (en) Timothy White, Berhane Asfaw et al., « Pleistocene Homo sapiens from Middle Awash, Ethiopia », Nature, vol. 423, no 6491, , p. 742-747 (DOI 10.1038/nature01669, lire en ligne)
- Laure Cailloce, « À quoi ressemblait l’ancêtre d’Homo sapiens ? », Le Journal du CNRS, (lire en ligne)
- (en) Eleanor Scerri, Mark Thomas et al., « Did Our Species Evolve in Subdivided Populations across Africa, and Why Does It Matter ? », CellPress Trends in Ecology & Evolution, vol. 33, no 8, , p. 582-594 (DOI 10.1016/j.tree.2018.05.005, lire en ligne)
- (en) Eric Delson, « An early dispersal of modern humans from Africa to Greece », Nature, (DOI 10.1038/d41586-019-02075-9)
- (en) Katerina Harvati, Carolin Röding, Abel M. Bosman, Fotios A. Karakostis, Rainer Grün et al., « Apidima Cave fossils provide earliest evidence of Homo sapiens in Eurasia », Nature, (DOI 10.1038/s41586-019-1376-z)
- (en) Israel Hershkovitz et al., « The earliest modern humans outside Africa », Science, vol. 359, no 6374, , p. 456-459 (DOI 10.1126/science.aap8369)
- Bruno Maureille, Les premières sépultures, Paris, Le Pommier, Cité des sciences et de l'industrie, 2004, 123 p.
- (en) Xue-feng Sun et al., Ancient DNA and multimethod dating confirm the late arrival of anatomically modern humans in southern China, pnas.org, février 2021, doi.org/10.1073/pnas.2019158118
- (en) « Ancestral tools », Handprint.com, (consulté le )
- Middle to upper paleolithic transition
- Cann, R. L., Stoneking, M., Wilson, A. C. (1987), « Mitochondrial DNA and human evolution », Nature, vol. 325., pp. 31-36
- Vigilant, L., Stoneking, M., Harpending, H., Hawkes, K., Wilson, A. C. (1991) « African Populations and the Evolution of Human Mitochondrial DNA », Science, vol. 253, pp. 1503-1507
- Whole-mtDNA Genome Sequence Analysis of Ancient African Lineages
- mtDNA Variation in the South African Kung and Khwe
- (en) Fulvio Cruciani, Beniamino Trombetta, Andrea Massaia, Giovanni Destro-Bisol, Daniele Sellitto et Rosaria Scozzari, « A Revised Root for the Human Y Chromosomal Phylogenetic Tree : The Origin of Patrilineal Diversity in Africa », The American Journal of Human Genetics, (lire en ligne)
- (en) Coop G, Pickrell JK, Novembre J, Kudaravalli S, Li J, et al, « The Role of Geography in Human Adaptation », PLoS Genet, vol. 5, no 6, , p. 1–16 (DOI 10.1371/journal.pgen.1000500, lire en ligne)
- « Among Many Peoples, Little Genomic Variety », The Washington Post, (consulté le )
- « Geography And History Shape Genetic Differences In Humans », Science Daily, (consulté le )
- (en) Nicole Maca-Meyer, Ana M. González, José M. Larruga, Carlos Flores et Vicente M. Cabrera, « Major genomic mitochondrial lineages delineate early human expansions », BMC Genetics, vol. 2, no 13, (DOI 10.1186/1471-2156-2-13)
- (en) Marta Mirazón Lahr, Mike Petraglia et Stephen Stokes, « Searching for traces of the Southern Dispersal », University of Cambridge (consulté le )
- (en) Israel Hershkovitz, Ofer Marder et al., « Levantine cranium from Manot Cave (Israel) foreshadows the first European modern humans », Nature, vol. 520, no 7546, , p. 216–219 (PMID 25629628, DOI 10.1038/nature14134)
- « Sur les pas des premiers hommes modernes », sur Sciences et Avenir (consulté le )
- (en) John G. Fleagle, Primate Adaptation and Evolution, Academic Press, , 464 p. (ISBN 978-0-12-378633-3, lire en ligne), p. 386
Bibliographie
- (en) Aurélien Mounier et Marta Mirazón Lahr, « Deciphering African late middle Pleistocene hominin diversity and the origin of our species », Nature, (lire en ligne)
- (en) Jean-Jacques Hublin, Abdelouahed Ben-Ncer et al., « New fossils from Jebel Irhoud, Morocco and the pan-African origin of Homo sapiens », Nature, vol. 546, , p. 289-292 (DOI 10.1038/nature22336)
- (en) Christopher Brian Stringer, « The origin and evolution of Homo sapiens », Philosophical Transactions of the Royal Society B - Biological Sciences, (lire en ligne)
- (en) Tim J. Crow, Editor The Speciation of Modern Homo Sapiens, (ISBN 0-19-726311-9) (paperback) 2002.
- (en) Stephen Oppenheimer, The Real Eve : Modern Man's Journey Out of Africa, (ISBN 0-7867-1192-2) (Hardcover), 2003.
- (en) Nicholas Wade, Before the Dawn : Recovering the Lost History of Our Ancestors, 2006 (ISBN 1-59420-079-3)
- (en) Wells, Spencer, The Journey of Man : A Genetic Odyssey, 2003 (ISBN 0-691-11532-X)
- (en) Wells, Spencer, Deep Ancestry : Inside the Genographic Project, 2006 (ISBN 0-7922-6215-8)
- (en) "New Research Proves Single Origin Of Humans In Africa,", Science Daily,
- (en) Ancient African Megadroughts May Have Driven Human Evolution Out Of Africa,
- (en) DNA studies trace human migration from Africa,
Classiques
- Charles Darwin, La Filiation de l’Homme et la sélection liée au sexe, 1871, trad. sous la direction de Patrick Tort, coordonnée par M. Prum. Précédé de Patrick Tort, « L’anthropologie inattendue de Charles Darwin », Paris, Champion Classiques, 2013.
- Luigi Luca Cavalli-Sforza & Francesco Cavalli-Sforza, Qui sommes-nous ? Une histoire de la diversité humaine, éd. Albin Michel, 1994 (titre original : Chi Siamo : La Storia della Diversità Umana, 1993), (ISBN 0-201-44231-0).
Voir aussi
Articles connexes
- Origine multirégionale de l'homme moderne
- Premières migrations humaines hors d'Afrique
- Modernité comportementale
- Effet de pompe du Sahara
- Théorie de la catastrophe de Toba
- Expansion planétaire de l'Homme moderne
- Dispersion australe
- Dernier ancêtre commun
- Ève mitochondriale
- Plus récent ancêtre patrilinéaire commun
- Macrohaplogroupe L
- Identical ancestors point (en)
Liens externes
 Portail de la Préhistoire
Portail de la Préhistoire .svg.png.webp) Portail de l’Afrique
Portail de l’Afrique