Méiose
La méiose (du grec μείωσις, meiōsis, « petit », « amoindrissement », « diminution »[1]), est un processus de double division cellulaire découvert par Edouard Van Beneden (1846-1910) et qui prend place dans les cellules (diploïdes) de la lignée germinale pour former les gamètes (haploïdes).
Pour la figure de style, voir Méiose (figure de style).
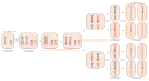
Généralités
Il existe deux types de divisions cellulaires chez les eucaryotes : la mitose, qui concerne les cellules somatiques et assure la naissance de cellules identiques à la cellule mère lors de la multiplication asexuée (elle conserve donc l'information génétique), et la méiose, qui aboutit à la production de cellules sexuelles ou gamètes pour la reproduction.
Chez les animaux, la méiose est un processus se déroulant durant la gamétogenèse (spermatogenèse ou ovogenèse), c'est-à-dire durant l'élaboration des gamètes (les spermatozoïdes chez le mâle et les ovules chez la femelle) chez les espèces dites diploïdes.
Chez les champignons, la méiose produit des spores, qui, par mitose, donneront une génération haploïde. Chez les angiospermes (plantes à fleurs), la méiose produit des cellules de grains de pollen. Elle donne des cellules haploïdes (cellules contenant n chromosomes) à partir de cellules diploïdes (cellule contenant 2n chromosomes — chez l'homme, une cellule normale contient 2n = 46 chromosomes (donc 23 paires) alors qu'un gamète contient n = 23 chromosomes au cours de deux divisions). Chez les espèces haploïdes (comme la Sordaria macrospora), la méiose intervient après la fécondation pour diviser la cellule-œuf (à 2n chromosomes). Mais en plus de ce rôle de division, la méiose a un rôle important dans le brassage génétique (mélange des gènes) et ce, grâce à deux mécanismes de brassage : le brassage interchromosomique et le brassage intrachromosomique.
Ainsi, durant la méiose, la quantité d'ADN au sein de la cellule évolue.
Chaque cellule divise son patrimoine génétique (contenu dans des chromosomes) en deux afin de n'en transmettre que la moitié aux cellules filles. Elle se déroule en plusieurs étapes formant un ensemble de deux divisions cellulaires, successives et inséparables. La première division méiotique est dite réductionnelle car elle permet de passer de 2n à n chromosomes doubles. La seconde est dite équationnelle car elle conserve le nombre de chromosomes, de n chromosomes doubles à n chromosomes simples. La méiose permet ainsi la formation de quatre cellules filles haploïdes (ou gamètes).
Par ailleurs le cycle de la méiose est étroitement lié au cycle du centrosome[2].
Schéma
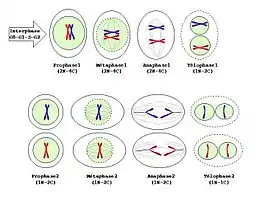
Les différentes étapes
Elle fait suite à la réplication de l'ADN. Elle permet le passage d'une cellule mère dite diploïde à 2n chromosomes homologues à deux chromatides à quatre cellules fille haploïdes à chromosomes simples.
Première division méiotique
Elle comporte les quatre phases de toute division cellulaire, mais présente des particularités.
Prophase I
L'enveloppe nucléaire disparaît. Les chromosomes bichromatidiens s’individualisent par condensation de leur ADN à partir de la chromatine du noyau. Ils apparaissent doubles car formés chacun de deux chromatides. Ils s’associent ensuite par paires de chromosomes homologues. Cet appariement donne des tétrades (car 4 chromatides) aussi appelées bivalents (car 2n chromosomes homologues). Cette phase est divisée en cinq étapes qui correspondent à cinq états caractéristiques de la chromatine : leptotène, zygotène, pachytène, diplotène et diacinèse.
- Leptotène : du grec lepto-, mince, et -tène, bandelette. Les chromosomes apparaissent très fins, très flexueux et très longs; Au ME les chromosomes sont bien dupliqués et donc la phase S a déjà eu lieu(ce sont les caractéristiques de ce stade) . Début de la condensation de la chromatine et attachement des télomères (extrémités des chromosomes) à l'enveloppe nucléaire par la plaque d'attache. Les chromatides s'associent par chromatides sœurs. Les deux brins provenant du père ensemble (bleu) et les deux brins provenant de la mère ensemble (rouge).
- Zygotène : du grec zygo-, liaison entre deux éléments. Début de l'appariement des chromosomes homologues (synapsis) par le complexe synaptonémal (ou synapton) et convergence depuis les télomères vers le centromère (un peu comme une fermeture éclair). Le complexe synaptonémal est une structure complexe constituée d'un élément central, SYCP1 qui forme un homodimère, relié à deux éléments latéraux. Les éléments latéraux sont en fait les cohésines SMC1, SMC3 formant un hétérodimère maintenu en place par mREC8 hHR21. Les cohésines se trouvent de part et d'autre par des filaments transverses ; à celles-ci se lie la chromatine de chaque zone des chromosomes impliqués dans le phénomène ultérieur d'enjambement (ou crossing-over). Il y a organisation « en bouquet » des chromosomes (on parle d'ikebana). L'ensemble des deux chromosomes homologues s'appelle une tétrade (car 4 chromatides) ou un bivalent (car 2 chromosomes).
- Pachytène (phase la plus longue, elle dure environ 2 semaines pour des spermatozoïdes humains) : du grec pachy-, épais. Appariement strict des chromosomes homologues et apparition des nodules de recombinaison (intra-chromosomique) et de nodules tardifs qui permettent les enjambements (échanges entre chromatides homologues; lesquelles échanges sont facilitées par la mise en place au préalable d'un chiasma). Cette phase a une importance considérable dans le brassage chromosomique (crossing-over).
- Diplotène : du grec diplo-, double. Désynapsis (séparation des chromosomes homologues), mais les chromosomes restent attachés en plusieurs points au niveau desquels deux des quatre chromatides semblent s'entrecroiser (recombinaison intra-chromosomique) (chiasma). Pour le bon déroulement de la méiose il en faut au minimum un par chromosome, en moyenne 2-3. Il y a décondensation de la chromatine et formation des grandes boucles permettant un fort taux de transcription. Cette étape de la prophase I peut durer plusieurs années chez l'ovocyte. En effet au cours de ce stade, l'ovocyte I augmente de volume, la décondensation des chromosomes permet la synthèse d'ARN messager et d'ARN ribosomiques qui seront stockés dans le cytoplasme et serviront de réserve pour le futur zygote lors des premières division de segmentation.
- Diacinèse : du grec dia-, à travers. Recondensation de la chromatine et détachement des télomères de l'enveloppe nucléaire. Glissement des chiasmata vers les télomères (terminalisation des chiasmata). À la fin, il y a disparition de l'enveloppe nucléaire.
Lors de cette phase, une séparation des centrosomes s'effectue. Ainsi, 2 centrosomes génèrent des fuseaux de fibres entre eux et s'éloignent l'un de l'autre.
Prométaphase I
Dans le cas de la première division de méiose, les microtubules kinétochoriens s'attachent aux chromosomes. Il y a deux kinétochores par chromosome, sur le centromère toujours, mais à la différence d'une mitose, ils ne sont pas diamétralement opposés, mais côte à côte (de manière à séparer uniquement les chromosomes homologues) lors de cette phase. Il y a alors un accrochage mon ou synthélique de chaque chromosome d'un bivalent, et donc un accrochage amphitélique de chaque bivalent. Il faut savoir que chez les mammifères, il n'y a pas juste un seul microtubule par kinétochore mais entre 15 et 40.
Métaphase I
Les paires de chromosomes homologues (bivalents) s'installent en vis-à-vis de part et d'autre du plan équatorial. Les chiasmas entre chromosomes homologues (et non les centromères comme en métaphase II) se placent de part et d'autre du plan équatorial. Leur orientation se fait de façon aléatoire : ce phénomène est appelé « ségrégation aléatoire ». Cette ségrégation engendre un brassage interchromosomique, lié à la répartition aléatoire des bivalents de part et d'autre du plan équatorial.
Durant cette phase, 2 kinétochores vont s'attacher à chaque centrosomes grâce à la formation d'un fuseau mitotique, les 2 kinétochores d'un même centrosomes sont dirigés vers le même pôle d'une cellule, et ceux du chromosome homologue vers l'autre pôle[3].
Anaphase I
La contraction de la fibre du fuseau achromatique entraîne la disjonction des deux chromosomes homologues de chaque paire et migrent aux pôles opposés, tirés par des microtubules kinétochoriens (microtubules accrochés à un kinétochore au niveau d'un centromère) dû à la dépolymérisation de tubuline. Il n'y a pas clivage des centromères, car SGO1 protège hRec8 de la séparase. Ainsi, en méiose, les chromatides homologues restent attachées au lieu de se séparer comme en mitose.
Durant cette phase, les microtubules du fuseau de division tirent les chromosomes homologues vers les pôles opposés de la cellule.
Télophase I et Cytodiérèse I
On observe une disparition du fuseau mitotique créée en métaphase, puis une séparation et individualisation en 2 cellules par cytodiérèse[4].
Deuxième division méiotique
Chaque chromosome étant resté dupliqué (n chromosomes à 2 chromatides), il n'y a pas de réplication de l'ADN et la division se déroule immédiatement. Cette division consiste en une simple mitose classique : pour chaque cellule, on passe de n chromosomes doubles à deux cellules à n chromosomes simples. L'étape Prométaphase II n'existe pas.
Prophase II
Le principe est le même que pour la mitose : les microtubules kinétochoriens s'attachent au kinétochores, à raison d'un kinétochore par chromatide, de part et d'autre du centromère (de manière à séparer les chromatides).
Métaphase II
Les chromosomes se placent sur la plaque métaphasique (et non plus équatoriale) par leur centromère. Leur condensation est maximale.
Durant cette phase, les centrosomes migrent aux pôles opposés de la cellule et les microtubules se fixent aux kinétochores, complexes protéiques préalablement attachés aux centromères des chromosomes.
Anaphase II
Les chromatides sœurs de chaque chromosome se séparent après rupture de leur centromère et migrent vers les pôles opposés de la cellule tirées par des fibres protéiques.
Durant cette phase, les microtubules du fuseau de division tirent les chromatides sœurs vers les pôles opposés de la cellule.
Télophase II
Les quatre cellules haploïdes issues de la méiose possèdent n chromosomes simples.
Durant cette phase, chaque centrosome se répartit dans les deux cellules filles formées.
Diversité des gamètes
Les gamètes créés par la méiose sont différents bien qu'ils descendent de la même cellule. Cette différenciation joue un rôle clef dans l'évolution des espèces et leur diversité génétique.
Brassage allélique par ségrégation indépendante des chromosomes homologues (brassage inter-chromosomique)
(cas des espèces à caryotype 2n)
Un premier facteur de diversité facile à comprendre provient de l'attribution aléatoire des allèles, c'est-à-dire de chacun des deux chromosomes d’une même paire (chromosomes homologues) vers les cellules filles haploïdes. Au moment de la métaphase I de la méiose, les chromosomes se disposent aléatoirement de part et d’autre du plan équatorial. Chaque chromosome (allèle) d'une paire migre ensuite vers un pôle (anaphase I), sans influencer les sens de migration des allèles des autres paires. Chaque cellule fille possédera donc un jeu de chromosomes (et donc de gènes) différent de celui de la cellule mère. Cette différenciation est appelée brassage inter-chromosomique.
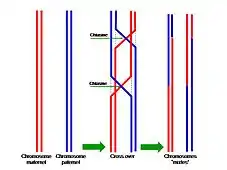
Échange d'allèles au sein d’une paire de chromosomes
À chaque méiose, sauf cas exceptionnels (drosophile mâle par exemple), il peut se produire un échange réciproque de fragments de chromatides appartenant à deux chromosomes homologues : c’est le phénomène d’enjambement, également appelé crossing over, qui survient pendant la prophase I (donc avant la séparation métaphasique des chromosomes homologues). Cet enjambement est provoqué par un module de recombinaison (complexe multi-enzymatique). Les chromatides recombinées se distinguent des chromatides d'origine ; on parle alors de brassage intra-chromosomique.
C'est aussi durant cette période que des échanges par crossing-over asymétriques de fragments inégaux de chromatine peuvent être réalisés et donner naissance à des gènes dupliqués sur l'une des chromatides, faisant apparaître une famille multigènique. Dans le cas de crossing over entre chromosomes non homologues, il y a alors transposition de gène.
Superposition des deux brassages alléliques
La superposition des deux brassages permet une diversité considérable des gamètes.
- S'il possède paires de chromosomes, le seul brassage interchromosomique permet arrangements possibles.
- Si l'individu possède gènes hétérozygotes, le seul brassage intrachromosomique permet arrangements possibles.




Anomalie de la méiose
- anomalie de nombre : non disjonction des chromosomes = aneuploïdie (par exemple la trisomie 21)
- anomalie de structure : translocation
- erreur de réplication de l'ADN : mutation
Reprise de la méiose
La reprise de la méiose (en) se produit dans le cadre de la méiose de l'ovocyte après un arrêt méiotique. Chez la femme, la méiose d'un ovocyte commence pendant l'embryogenèse et se termine après la puberté[5]. Un follicule primordial s'arrête, ce qui permet au follicule de croître en taille et de mûrir. La reprise de la méiose se fera après une poussée ovulatoire (ovulation) d'hormone lutéinisante (LH).
Mitose et méiose
La méiose et la mitose sont différentes en plusieurs points mais elles ont aussi de grandes similitudes. La mitose se produit au cours de la multiplication asexuée alors que la méiose a sa place dans la reproduction. Presque toutes les cellules peuvent subir une mitose alors que la méiose ne concerne que celles des organes de reproduction chez les espèces diploïdes (les ovogonies et les spermatogonies) ou la cellule-œuf chez les espèces haploïdes. À la fin de la mitose, il y a deux cellules génétiquement identiques alors qu'à la fin de la méiose il y a quatre cellules qui ne sont pas nécessairement génétiquement identiques.
Différences entre mitose et méiose
Ces deux types de division cellulaire interviennent dans la reproduction. La mitose est un mécanisme de la reproduction asexuée, et crée des clones à partir d'organismes unicellulaires. Ces clones sont par conséquent identiques sur le plan génétique (sauf dans le cas de mutations accidentelles). La méiose, quant à elle, crée quatre gamètes à l'identité génétique distincte. Ce procédé permet d'amplifier la diversité génétique au sein d'une espèce.
Ces deux types de reproduction ont des avantages comparés dépendants de l'environnement dans lequel les espèces évoluent. Ainsi, la mitose est plus avantageuse pour des espèces évoluant dans un milieu stable car, l'individu déjà adapté à son milieu créé des clones qui seront également adaptés. De plus ce type de reproduction est moins coûteux en énergie que la méiose. La méiose, elle, est plus avantageuse pour des espèces évoluant dans un environnement instable[6]. Elle est plus avantageuse car elle permet de produire une plus grande diversité. Cette dernière permet l'apparition d'individus dont les différences phénotypiques augmentent les possibilités de survie dans le milieu instable en question. Ce processus suit la théorie de l'évolution de Charles Darwin[7].
Il est particulièrement apparent dans les mécanismes de reproduction des levures, notamment les Saccharomyces cerevisiae, espèce capable de se reproduire par méiose comme par mitose. Ces levures se reproduisent ordinairement par mitose lorsqu'elles sont dans un environnement comportant assez de ressources pour survivre. Cependant, lorsqu'elles sont placées dans un environnement faible en nutriments (on parle alors de « sporulation media » ou SPO), les levures font alors de la méiose et créent quatre spores. Ce processus permet d'amplifier la diversité génétique, ce qui augmente les chances de survie de l'espèce.
Notes et références
- « Méiose », sur cnrtl.fr
- « Le cycle cellulaire : la mitose et la méiose », sur esna.csdccs.edu.on.ca, (consulté le )
- Cours de PACES Purpan (fac de Medecine Toulouse)
- Cours de PACES (injustement appelée 1ere année de médecine) de Purpan (Toulouse)
- (en) Laurinda A. Jaffe et Jeremy R. Egbert, « Regulation of Mammalian Oocyte Meiosis by Intercellular Communication Within the Ovarian Follicle », Annual Review of Physiology, vol. 79, no 1, , p. 237–260 (ISSN 0066-4278, PMID 27860834, PMCID 5305431, DOI 10.1146/annurev-physiol-022516-034102, lire en ligne)
- (en) Dusan Misevic, Charles Ofria et Richard E. Lenski, « Experiments with Digital Organisms on the Origin and Maintenance of Sex in Changing Environments », Journal of Heredity, (lire en ligne [PDF])
- (en) « Darwin Online », sur darwin-online.org.uk
Liens externes
- Animation en Flash de la méiose.
- Animation flash avec recombinaison génétique en prophase 1 de la méiose.
 Portail de la biologie cellulaire et moléculaire
Portail de la biologie cellulaire et moléculaire  Portail de la biologie
Portail de la biologie