Édouard Van Beneden
Édouard Joseph Louis Marie Van Beneden, né à Louvain le et mort à Liège le , est un zoologiste et embryologiste belge, professeur à l'Université de Liège, découvreur de la méiose.
Pour les articles homonymes, voir Van Beneden.

| Naissance |
Louvain (Belgique) |
|---|---|
| Décès |
Liège (Belgique) |
| Nationalité |
|
| Domaines | zoologiste, embryologiste |
|---|---|
| Institutions | Université de Liège |
| Renommé pour | Découverte de la méiose |
| Distinctions | Prix quinquennal des Sciences en 1871, 1887 et 1891 |
Il était le fils du paléontologue Pierre-Joseph Van Beneden (1809-1894).
Biographie
Les informations de cette section sont issues en majeure partie du livre de Gabriel Hamoir[1].
Jeunesse
Édouard van Beneden est fils de Rosalie Valcke et de Pierre-Joseph van Beneden, professeur de zoologie, d'anatomie comparée et de paléontologie à l'Université Catholique de Louvain depuis 1836. Il a quatre sœurs. Il fait sa scolarité au collège du Petit Séminaire de Saint-Trond. Édouard termine sa rhétorique en 1863 avec un 1er prix d'excellence au classement général. Il suit brillamment pendant deux ans le double cursus de licence en Sciences Naturelles et en Sciences appliquées. Il hésite sur la voie à suivre. En 1865, il jette un coup d'œil dans un microscope sur des vers parasites, des Cysticerques, que son père étudiait. Il semble que cette expérience l'ait définitivement motivé pour la biologie plutôt que pour l'ingénierie. En 1865-1866, il suit les cours d'anatomie comparée de son père. En 1867, il obtient le titre de docteur en Sciences Naturelles avec la plus grande distinction. Édouard van Beneden est plus intéressé par la biologie fondamentale que par la systématique. Il se lie d'amitié avec un confrère et ami de son père, Theodor Schwann, qui avait été professeur à Louvain mais qui était à ce moment professeur à l'Université de Liège.
En 1868, alors qu'il n'a que 22 ans, l'Académie Royale lui décerne le premier prix et une médaille d'or pour sa réponse à la question « Faire connaître la composition anatomique de l'œuf dans les différentes classes du règne animal, son mode de développement et la signification des différentes parties qui la constitue ». Sa réponse, intitulée « Sur la composition et la signification de l'œuf », démontre, en s'appuyant sur de multiples exemples tirés de la riche collection zoologique de son père, que tout œuf est une cellule composée d'un noyau, d'un protoplasme et d'une enveloppe, et que les différences observées entre espèces sont d'importance secondaire. Il affirme :
- « Rien de ce qui vit ne prend naissance par génération spontanée ; toute unité vitale procède d'une unité vitale antérieure ; l'organisation et la vie ne commencent ni ne se terminent jamais ; leur durée est à la fois continue et indéfinie ; elles sont continues en ce que le seul mode de reproduction des unités vivantes est la division ; indéfinie, grâce à la syncytose (fusion de deux ou plusieurs cellules) ; la doctrine de l'évolution est donc seule conforme aux principes des sciences positives ; elle nous apparaît comme la conséquence des lois de la propagation des êtres. »
Cette déclaration est très conforme aux vues de Theodor Schwann sur l'œuf. Édouard continue à travailler, spécialement, dans le petit laboratoire de biologie marine créé à Ostende par son père à qui il soumet ses découvertes. Grâce aux contacts de son père et de Schwann, Édouard fait un grand voyage en Europe en 1868-1869. En Allemagne il visite Max Schultze à Bonn, Hermann von Helmholtz à Heidelberg, Karl von Siebold à Münich, et Karl Leuckart à Giessen. À Wurzbourg, il rencontre le Suisse Albert von Kölliker, père de l'histologie et embryologiste. Il séjourne aussi trois mois à Concarneau où il étudie les Copépodes, parasites des poissons. Il va aussi à Londres où il rencontre un grand ami, Ray Lankester.
Professeur
Van Beneden se porte candidat à un poste de professeur de physiologie à l'Université de Louvain. Il n'est pas retenu mais l'Université de Liège lui propose la succession de Théodore Lacordaire. Il est nommé chargé de cours de zoologie et d'anatomie comparée à Liège en 1870, professeur associé en 1871 et, finalement, professeur ordinaire en 1874 pour les cours de zoologie, d'anatomie et physiologie comparées et enfin d'embryologie. Voici ce qu'écrit Édouard à ses parents le après avoir donné ses deux premiers cours (cité par Hamoir[1] page 34) :
- « Très chers parents,
- Je viens de terminer ma seconde leçon que je considère plutôt comme mon début. J'en suis très satisfait. J'ai toute confiance maintenant dans l'avenir et je suis tranquille. J'ai parlé pendant 1 h 1/2 sans aucune note. Je me trouvais aussi à l'aise qu'à ma chambre et je n'ai pas perdu du tout le fil de mes idées. J'ai parlé assez lentement pour pouvoir leur permettre de prendre des notes et tous en prennent depuis le commencement jusqu'à la fin de la leçon. L'attention s'est soutenue jusqu'à la fin et je n'ai qu'à me louer de l'attitude de mon auditoire. J'avais plus de 50 auditeurs. Mais je suis très fatigué.Une heure et demie, c'est trop long. Je vous donnerai tous les détails samedi...
- Spring a dit hier à Vanlair qu'il avait appris que les élèves avaient été très satisfaits de mon début.
- Je vais me reposer aujourd'hui... Adieu, à samedi.
- Votre tout dévoué Édouard »
De leur côté, les étudiants seront toujours fascinés par son enseignement. Albert Brachet qui a suivi les leçons d'Édouard van Beneden entre 1887 et 1890 témoigne[2] :
- « Dès sa première leçon, il s'imposait à ce milieu complexe, formé pourtant d'une grande majorité d'indifférents auxquels ne se mêlaient que quelques esprits curieux vraiment désireux d'apprendre et de comprendre. Sans jamais s'aider de notes, il parlait assez lentement pour que ses auditeurs puissent prendre un résumé complet de la leçon et reproduire les nombreux croquis dont il accompagnait son exposé. Malgré les complications progressives d'un cours étendu, l'attention ne se relâchait pas. C'est parce qu'un lien solide et continu enchaînait tous les faits particuliers qu'il exposait et que ce lien ressortait, se dégageait peu à peu, et l'on apercevait alors qu'il était fait de deux grandes idées, bases fondamentales de la biologie morphologique, dont il voulait avant tout que ses élèves fussent imprégnés : le transformisme et la théorie cellulaire. » Il faut noter qu'Édouard van Beneden enseignait l'évolutionnisme (Darwin) plutôt que le transformisme (Lamarck), théories que Brachet semble confondre.
Quelques années plus tard, Hans von Winiwarter suit les cours de Van Beneden pendant sa première année d'études de médecine (cité par Hamoir[1] pages 36-37) :
- « Je me souviendrai toujours de ma première leçon de zoologie qui fut aussi le premier cours auquel j'assistais à l'Université. Ce fut une véritable révélation, mais aussi ce jour-là mon destin fut fixé (...) J'ai la conviction que bien d'autres que moi ressentirent un pareil coup de foudre qui modifia peut-être leur carrière ultérieure ou influença leurs recherches.
- De plus Van Beneden possédait un extraordinaire talent d'exposition. Les problèmes les plus complexes comme les questions les plus ardues étaient présentés en phrases simples et claires composées de mots courants, renforcés de temps en temps par des comparaisons imagées mais tout aussi banales. Sa démonstration se déroulait tout naturellement sans difficulté apparente ; et tout à coup, la conclusion s'imposait d'elle-même, et, chose plus surprenante, se gravait dans la mémoire au point qu'il n'était plus nécessaire de revoir les notes prises en cours...
- Au cours de zoologie, le maître s'attachait aux principes généraux, à l'étude de la cellule et des groupes d'animaux inférieurs, ne rappelant que les grandes lignes des classes les plus élevées. D'aucuns n'ont pas compris la portée d'une telle délimitation. Pour le futur médecin cependant, les animaux inférieurs, surtout les protozoaires, prennent de jour en jour une importance plus grande et la connaissance approfondie de l'organisme élémentaire, la cellule, est indispensable pour la juste compréhension de l'histologie, de l'embryologie et de l'anatomie...
- Quant au cours d'embryologie, il constituait un exemple vraiment unique en son genre car Van Beneden l'avait pour ainsi dire bâti de toutes pièces sur des recherches personnelles. »
Après s'être installé à Liège et avoir achevé sa première année universitaire, en juillet et , il retourne en Allemagne. À Iéna, il rencontre Anton Dohrn, au moment où ce dernier cherche à créer une station zoologique à Naples, ce qui sera réalisé en . Il rencontre également Karl Gegenbaur, grand morphologiste, et Ernst Haeckel, grand évolutionniste.
Van Beneden est admis à l'Académie Royale des Sciences, des Lettres et des Beaux-Arts de Belgique, comme membre associé le et comme membre titulaire le .
En 1872, à 30 ans, il effectue un voyage en bateau au Brésil, voyage qui évoque celui du jeune Charles Darwin à bord du Beagle quarante ans auparavant. Il observe méthodiquement la faune aquatique au large de Rio de Janeiro. Il découvre une nouvelle espèce de dauphin, Sotalia Brasilensis.
À son retour en 1873, il crée son propre laboratoire à l'Université de Liège. Il obtient, en particulier la création d'un laboratoire de microcopie qui permet à ses assistants de travailler et aux étudiants qui le désirent d’accéder aux observations microscopiques.

En 1880, il fonde, avec Charles Bambeke, les Archives de Biologie.
Vers 1880, une nouvelle loi introduit l'enseignement des sciences naturelles à l'école primaire et préconise de développer le sens de l'observation des enfants. Les instituteurs de Wallonie sollicite Édouard van Beneden pour leur apporter les connaissances indispensables pour remplir leur nouvelle mission. Van Beneden répond avec enthousiasme. Du 1er au , il réunit les instituteurs et leur prodigue 10 leçons et 7 séances de travaux pratiques. Il rédige un texte de 159 pages illustré de 70 figures. Les années suivantes, le cours d'été est enrichi et donné par Julien Fraipont et Charles Julin, assistants de van Beneden.
Grâce à son beau-père, Pierre-Joseph van Beneden avait installé, à ses frais, un laboratoire et un aquarium à Ostende en 1843 pour initier les recherches de biologie marine en Mer du Nord. En 1883, Édouard van Beneden et Charles Bambeke installent les bases d'une station de biologie marine publique à Ostende dans un bâtiment appartenant à l'État. Ils se procurent un petit chalutier. Van Beneden obtient le soutien financier de l'Université de Liège pour développer la station.
De 1885 à 1888, les professeurs Édouard Van Beneden, Constant-François Vanlair et Voltaire Masius élaborent avec l'architecte Lambert Noppius les plans du grand Institut de Zoologie de l'Université de Liège qui est construit sur la rive droite de la Meuse, sur le Quai Édouard van Beneden.
Édouard van Beneden meurt à Liège le à l'âge de 64 ans.
Son élève Paul Cerfontaine publia ses derniers travaux et poursuivit son œuvre après sa mort.
L'influence de Van Beneden fut considérable à Liège, où une importante école d'embryologie comparée et expérimentale se constitue autour et après lui.
Famille

Le , à 36 ans, Édouard van Beneden épouse Berthe Dequesne, fille d'un magistrat de Bruxelles. Ils auront trois filles : Juliette en 1884, Hélène (Nelly) en 1885 et Marie-Louise (Amy) en 1886.
Édouard Van Beneden loue ensuite le château de Ramelot comme résidence secondaire afin de veiller à l’épanouissement de la famille.
Cependant, à trois ans, Juliette est atteinte d’une paralysie infantile et doit être appareillé au niveau des jambes. Un drame survient ensuite en : Hélène est atteinte d’une maladie respiratoire et malgré les efforts pour la sauver, elle meurt quelques jours plus tard dans les bras de son père. De plus, Berthe et ses deux filles sont contraintes de s’isoler dans une auberge non loin afin de prévenir toute contagion. Cette disparition affectera grandement Berthe et Édouard qui décident par conséquent de quitter le domaine de Ramelot.
En , il fait l’acquisition du château de Résimont lors d’une vente publique et décide d’y entreprendre une série de travaux de réaménagement qui dureront plusieurs années et qui transformeront intégralement le domaine. À partir de là, Résimont fut considéré comme un havre de paix pour la famille et son entourage. À l’aube de ses soixante ans, cet amoureux du Condroz délaisse peu à peu la rigueur scientifique qu’il a pratiqué toute sa vie au profit de la chasse et des promenades dans la campagne avoisinant le château.
Lorsqu’il meurt en 1910, un nouveau chapitre commence à Résimont. Marie-Louise, sa fille cadette, épouse le de la même année, le baron Robert Elie-Lefebvre, originaire du Havre en Haute-Normandie. Ils eurent tous deux 3 filles et 3 garçons. Ces nombreux arrivants dans la famille permirent à la veuve d’Édouard Van Beneden, fervente catholique et continuellement vêtue de noir, de jouer pleinement son rôle de grand-mère jusqu’à sa mort, 27 ans après le décès de son mari.
Juliette Van Beneden se maria avec Paul Elie-Lefebvre, le frère de Robert, le à Evelette. Ils n’eurent pas d’enfants.
Travaux
Présentation générale
- « L'œuvre scientifique d'Édouard van Beneden est très considérable par son étendue comme par l'importance des faits qui y sont relatés. Son analyse est cependant relativement aisée parce qu'elle offre un remarquable caractère d'homogénéité[2]. »
L'œuvre d'Édouard van Beneden est marquée, d'une part, par la poursuite de la recherche sur l'ovogenèse, la fécondation et le développement embryonnaire qu'il a initié en 1868, à 22 ans, en répondant à la question posée par l'Académie Royale de Bruxelles et, d'autre part, par la continuité avec l'œuvre de son père, éminent zoologiste systématicien dont il hérite deux intérêts majeurs : pour les animaux marins dont il poursuit l'étude avec la création de la station de biologie marine d'Ostende, officiellement rattachée à l'Université de Liège, et pour les parasites dont Pierre-Joseph van Beneden était un spécialiste reconnu.
| Position des taxons étudiés par Édouard van Beneden | |
La liste des publications d'Édouard van Beneden a été dressée par Albert Brachet[2]. Il en existe aussi une version accessible en ligne[3]. On peut les regrouper selon trois axes majeurs :
- La recherche sur les animaux marins. De 1868 à 1874, van Beneden travaille essentiellement sur les Arthropodes : Crustacés (7 publications) et Limulides (2 publications). Il décrit de nouvelles espèces et explore l'anatomie et le développement des Grégarines (6 publications) qui sont des Protistes parasites des Arthropodes. Entre 1881 et 1887, il publie 7 articles sur les Tuniciers et les Ascidies. À partir de 1890, il s'intéresse aux Anthozoaires (5 articles).
- La recherche sur l'ovogenèse et l'embryogenèse des Mammifères. Entre 1868 et 1874, il publie 4 exposés généraux sur l'œuf et la fécondation. En 1875, il décrit « La maturation de l'œuf, la fécondation et les premières phases du développement embryonnaire des Mammifères d'après des recherches faites chez le Lapin »[4], ce qui amorce une nouvelle voie de recherche qu'il complétera avec des études sur « le Murin », c'est-à-dire sur les Chiroptères (Chauves-souris), ce qui donnera 14 publications jusqu'en 1901 et 2 publications posthumes en 1911 et 1912.
- La recherche sur l'ovogenèse et la spermatogenèse du vers Ascaris. Au début de sa carrière, il publie 3 articles sur les Trématodes et Nématodes. En 1883-1884 et 1887, il explore l'ovogenèse, la spermatogenèse et l'embryogenèse chez Ascaris megalocephala, un parasite de l'intestin de Cheval, appelé maintenant Parascaris equorum, qui se prête particulièrement bien à l'étude de la maturation ovocytaire et de la fécondation. Il découvre les deux phases de la méiose et interprète correctement la réduction chromatique et la fécondation[5] (3 publications).
Édouard van Beneden cesse de publier après 1901 bien qu'il ait plusieurs sujets en état de rédaction avancée qui seront publiés après sa mort par Albert Brachet, Paul Cerfontaine et M. de Sélys.
Méiose : maturation et fécondation de l'ovocyte
_1980%252C_MiNr_2537.jpg.webp)
Édouard van Beneden est crédité de la découverte des principales étapes de la méiose et de leur signification. Sa méthode est essentiellement visuelle : observer les cellules à travers un microscope. Mais pour voir ce qu'il se passe, il lui faut :
- disposer de microscopes de bonne qualité, ce qui est le cas vers 1880. Van Beneden emploie des microscopes Hartnack et Zeiss. Edmund Hartnack a mis au point l'objectif à immersion en 1859, ce qui est un progrès considérable dans la résolution. Les optiques de Carl Zeiss ont bénéficié des compétences optiques d'Ernst Abbe à partir de 1866, puis de celles d'Otto Schott, chimiste spécialiste du verre, qui ont permis de commercialiser à partir de 1886 les objectifs apochromatiques, supprimant totalement les irisations colorées dues à la dispersion de la lumière blanche.
- éclairer les échantillons. L'éclairage est un problème majeur tant que l'lampe électrique à incandescence n'est pas en usage courant (commercialisée par Thomas Edison à partir de 1880). Les fabricants de microscopes ont introduit des loupes sur la platine et des miroirs et des condenseurs sous la platine pour focaliser la lumière. « Cette organisation de la cellule, dont Brüke affirmait l'existence (1861) sans pouvoir la dévoiler, l'on a commencé à l'analyser. Grâce au perfectionnement de la technique microscopique, grâce à des objectifs et des appareils d'éclairage plus parfaits, l'on entrevoit la complexité qui se cache sous une uniformité apparente. »
- détecter des objets transparents dans des cellules transparentes en notant les changements de réfringence. En effet, l'objectif à contraste de phase ne sera inventé qu'en 1930. Van Beneden dessine à la main en utilisant une chambre claire. Il publiera, avec Albert Neyt en 1887, les premières photographies prises au microscope.
- interpréter les images en 3D. Les cellules ne sont pas sectionnées en couches minces. Il faut donc constamment reconstituer l'objet observé à partir de l'image en 2D.
- tenir compte des artefacts introduits lorsque les cellules sont fixées (acide nitrique 3 % ou acide osmique 1 %), déshydratées (alcool à 40 puis 70 %) et colorées (carmin boracique).
- disposer du bon modèle biologique. Les chercheurs de l'époque étudient la fécondation sur des nématodes, des gastéropodes, des oursins, des étoiles de mer, des mammifères... De façon surprenante, ils observent des mécanismes semblables à travers les règnes animal et végétal. Cependant certaines espèces présentent des caractéristiques plus observables ou plus manipulables que d'autres.
- décrire et nommer les images découvertes souvent sans savoir ce que sont les objets... qui eux-mêmes changent d'aspect. Il est très difficile de voir les phénomènes quand l'observateur ne sait pas ce qu'il voit, n'a pas de mot pour désigner les choses. Il faut ensuite s'accorder entre collègues pour adopter le même vocabulaire, avec la difficulté de faire correspondre les mots créés en allemands avec les mots créés en français et, par la suite, en anglais. « La cellule nous apparaît aujourd'hui comme un organisme complexe dont les appareils nous sont inconnus ; nous devons commencer par débrouiller l'organisation cellulaire et étudier les modifications morphologiques qui accompagnent les actes de la vie... Nous sommes exposés, dans nos tâtonnements pour découvrir la route qui doit nous conduire au but dans ces régions inexplorées, à suivre souvent des chemins sans issues : l'essentiel se confondra pour nous avec l'accessoire: car la portée des faits nous échappe et nous en sommes réduits à rechercher par voie d'analyse les matériaux qui devront servir un jour à édifier le schéma de l'organisation élémentaire. »
Premier travaux
Dès ses premiers travaux (1868-1870), van Beneden s'est attaché à décrire les caractéristiques de l'œuf, de la fécondation et des premières étapes de l'embryogenèse. Il cherche à prouver que l'œuf, la cellule-œuf comme il l'appelle, est le germe initial de tout organisme. Même quand plusieurs cellules contribuent à la formation de l'œuf, il existe une seule cellule germinale. Les autres cellules ne sont que des accessoires. Pendant les 10-15 ans qui ont suivi, les observations se sont multipliées et les concepts se sont précisés.
Dans son ouvrage de 1883, Recherches sur la maturation de l'oeuf, la fécondation, et la division cellulaire, van Beneden indique que « le mérite d'avoir inauguré une ère nouvelle dans l'histoire de la fécondation revient avant tout à Otto Bütschli, l'éminent professeur de zoologie à Heidelberg. » Bütschli, dans ses publications de 1872-1876, démontre que :
- deux noyaux prennent naissance dans le vitellus de l'œuf fécondé, qu'ils se confondent pour donner naissance au noyau de la première cellule de l'embryon. Ce que Leolpold Auerbach découvre indépendamment à son tour en 1874. Mais Bütschli n'a pas élucidé l'origine des deux noyaux qu'il a observés dans l'ovocyte fécondé. Van Beneden signale que, dès 1850, Warneck a décrit deux noyaux dans des ovocytes fécondés de gastéropodes.
- il se forme un fuseau autour des éléments du noyau.
- ce fuseau intervient dans la formation des globules polaires (qu'il appelle corpuscules de rebut).
Les travaux de Bütschli sur la fécondation animale (nématodes) sont complétés par les observations d'Eduard Strasburger dans le domaine végétal (gymnospermes). En étudiant la fécondation chez le lapin en 1875, van Beneden constate que le déclenchement de la division cellulaire est provoqué par la présence de spermatozoïdes à proximité des ovocytes mais il n'observe jamais de spermatozoïdes pénétrer dans l'ovocyte[4]. Cette observation conforte la théorie du contact : la fécondation serait provoquée par la diffusion à travers la membrane de l'œuf d'un produit issu du spermatozoïde et qui provoque l'apparition d'un pronucleus périphérique. Ensuite, le pronucleus périphérique s'unit au pronucleus central pour former le noyau de l'œuf. En 1876, Oskar Hertwig porte un coup fatal à cette vieille théorie du contact[6]. Il démontre avec les œufs d'oursins que la fécondation consiste en la fusion de deux cellules germinales, ovocyte et spermatozoïde. Mais il considère que le nucléole de l'ovule fusionne avec le noyau du spermatozoïde tandis que le noyau de l'ovule disparait. Cette description ne satisfait pas van Beneden car dans ses propres observations menées sur des étoiles de mer, le nucléole disparait avant le noyau. D'autre part, beaucoup d’événements restent inexpliqués, comme la formation des globules polaires que l'on attribuait à la gamète mâle[7].
En 1876-1877, Hermann Fol décrit la fécondation chez l'étoile de mer Asteria glacialis et met en évidence :
- un système de filaments organisés en double étoile qu'il appelle amphiaster et qui se développe près de la surface de l'ovocyte.
- une étoile donne naissance à un premier globule polaire qui peut éventuellement se diviser.
- l'étoile qui reste dans l'ovocyte donne immédiatement un nouvel amphiaster qui se divise en deux : d'une part un deuxième globule polaire et d'autre part un pronucleus.
- le rapprochement du pronucleus femelle du pronucleus mâle.
Ascaris megalocephala
En 1884, van Beneden publie un travail remarquable sur la maturation des œufs d'Ascaris megalocephala[5], un vers parasite de l'intestin de cheval.
- « J'ai trouvé, dans l'Ascaride du cheval, un matériel admirable. Je suis convaincu que les œufs de ce nématode deviendront bientôt l'objet classique pour l'étude et la démonstration des phénomènes qui se rattachent à la fécondation. »
- « L'utérus n'a pas moins de 15 à 20 cm de longueur chez une femelle adulte. Dès le moment où ils y pénètrent, les œufs arrivent en contact avec les zoospermes. Quand ils sont arrivés dans le vagin, ils sont munis chacun de deux pronuclei ; l'expulsion des globules polaires s'est accomplie ; mais la segmentation n'a pas commencé. Je ne crois pas que l'on puisse désirer un matériel se prêtant plus merveilleusement à l'étude de la maturation et de la fécondation de l'œuf. »
Les avantages sont nombreux[8] :
- Les femelles peuvent être disséquées très facilement pour mettre à nu les organes sexuels. Ils sont constitués de deux ovaires tubulaires (~7 cm), prolongés chacun d'un oviducte (~3,5 cm) et d'un utérus (~18 cm) qui aboutissent dans le vagin (~1 cm)[9].
- Les ovocytes de chaque section d'oviducte, d'utérus ou de vagin sont mis en suspension dans une goutte de sérum physiologique et transférés sur une lame de microscope où il est possible de les observer soit vivants, soit fixés et colorés. En coupant des sections d'un 1/2cm, on obtient des milliers d'œufs au même stade de développement.
- Les phénomènes se déroulent lentement permettant de décrire les différents stades de maturation qui se déroulent simultanément le long du tractus génital.
- Les œufs peuvent être rendus transparents.
- Les spermatozoïdes d'Ascaris sont de forme conique et n'ont pas de flagelle. Ils se déplacent comme les amibes. Ils sont 10 fois plus gros que ceux de l'oursin et clairement visibles au microscope sans coloration.
- De plus, la chromatine du noyau, au moment de la mitose des cellules diploïdes, s'organise en un nombre constant de filaments ou « corpuscules chromatiques », auxquels von Waldeyer-Hartz donnera le nom de chromosomes en 1888. Van Beneden découvre que, chez Ascaris megalocephala, ce nombre est de quatre.

Bas : Planche XI, diverses formes des spermatozoïdes utérins. La nomenclature utilisée (spermatogonie, spermatocyte, spermatogemme, cytophore) est celle utilisée par van Beneden.
Formation des spermatozoïdes
- L'appareil sexuel mâle « est formé chez les Ascaris d'un tube unique dont le diamètre croît insensiblement de son extrémité aveugle vers son embouchure. Il suffit d'en examiner le contenu, aux divers points de sa longueur, pour établir la succession des phénomènes évolutifs. » Ce tube est constitué de quatre organes identifiables : le testicule, le canal déférent, la vésicule séminale et le canal éjaculateur[10].
- La formation des spermatozoïdes se déroule en trois phases :
- Phase de multiplication cellulaire - Dans la région haute du testicule se forment ce que van Beneden appelle les spermatomères qui à leur dernière division donnent des spermatogonies.
- Phase d'accroissement - Dans la région moyenne du testicule, il ne se forment plus de nouvelles spermatogonies ; mais ces cellules, très réduites d'abord, y grandissent considérablement.
- Phase de maturation - Les spermatogonies subissent une maturation en deux étapes, si bien que, « dans la région inférieure de l'utérus, chaque spermatogonie se résout en quatre spermatocytes qui restent unis entre eux de façon à constituer ensemble un spermatogemme. Chaque spermatocyte engendre ensuite une portion cytophorale, vers le centre du spermatogemme et les quatre parties cytophorales constituent ensemble un cytophore. Au moment de pénétrer dans le canal déférent, les spermatozoïdes se séparent du cytophore; ils deviennent libres et ils se modifient de façon à prendre peu à peu les caractères qu'ils affectent dans la vésicule séminale. »[10] À ce stade, les spermatozoïdes ont une forme sphérique.
- « C'est à (Karl Bogislaus) Reichert (1849) que l'on doit la découverte de ce fait capital de la formation constante, chez certains nématodes, de quatre spermatozoïdes aux dépens d'une même cellule mère. Reichert a fait ses observations chez deux petits Nématodes parasites de la Grenouille (Strongylus auricularis et Ascaris acuminata). (Hermann) Munk (1858) et après lui Rudolf Leuckart reconnurent que les grands Ascarides (A. megalocephala et A. lumbricoïdes) se comportent exactement comme les petits Nématodes des Batraciens... Nous ne faisons donc que confirmer sur ce point les données de nos prédécesseurs. »[10]

Bas: Planches XIV et XV, au centre, début de la formation du fuseau ; à gauche, figure ypsiliforme.
Pénétration des spermatozoïdes
- Les spermatozoïdes remontent du vagin vers la partie supérieure de l'utérus. Durant ce trajet ils changent de forme. De sphéroïdes, ils s'allongent, prennent la forme de poires, puis de cônes.
- Ils pénètrent les ovocytes (de forme ovoïde) situés dans le tiers supérieur de l'utérus et n'ont pas accès à l'oviducte. Dans les deux tiers inférieurs de l'utérus, ils sont en présence d'ovocytes déjà fécondés en cours de maturation.
- Un seul spermatozoïde pénètre un ovocyte. Il est extrêmement rare de trouver deux spermatozoïdes dans un même ovocyte. Il y a donc un mécanisme qui empêche la dispermie. Cette observation met un terme au débat qui agitait les chercheurs : certains avançaient qu'il fallait plusieurs spermatozoïdes pour féconder un ovocyte.
- Van Beneden insiste sur le fait que la pénétration du spermatozoïde n'est pas la fécondation. En effet « le zoosperme se maintient au milieu du vitellus sans se confondre avec lui... Les rapports entre les deux éléments cellulaires, avant la formation des globules polaires, peuvent être comparés à ceux qui relient un parasite à son hôte. Le résultat de la fécondation, c'est la formation d'une cellule capable de se diviser et de donner naissance à un individu semblable aux progéniteurs. Ce n'est qu'après l'expulsion du second globule polaire et la formation de la seconde couche périvitelline que le spermatozoïde subit les modifications qui annoncent l'imminence de la fécondation. »[5]
- Remarque : il faut garder à l'esprit que, chez Ascaris, à la différence de ce qui se passe dans l'ovogenèse des mammifères, la pénétration du spermatozoïde déclenche la maturation de l'ovocyte I et les deux phases de la méiose qui s’enchaînent sans diapause (voir le tableau dans l'article ovogenèse).
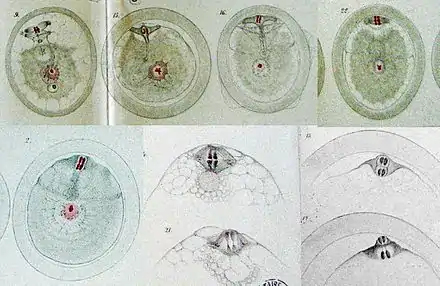
Bas : Planches XV et XVI, Formation du premier globule polaire.
Première phase de maturation et expulsion du premier globule polaire
- Dès que le spermatozoïde est complètement rentré, le noyau de l'ovocyte se transforme en une substance ponctuée et fibrillaire.
- À partir des résidus du noyau de l'ovocyte, se forme un fuseau entre deux pôles formés par des asters, selon une figure voisine de celle décrite dans la mitose par Walther Fleming. Au plan équatorial du fuseau, des globules chromatiques (=chromosomes) se répartissent en deux groupes de quatre.
- Cependant, la figure est plus complexe que celle de la mitose. À partir du fuseau, des fibrilles semblent se diriger vers le centre de la cellule. L'ensemble forme un Y que van Beneden appelle figure ypsiliforme. Les globules chromatiques sont positionnés à la croisée des branches du Y.
- Puis les pôles du fuseau se rapprochent de la membrane péricellulaire, les deux branches supérieures du Y s'alignent, formant un T. En vue latérale, la branche verticale du T disparaît. En vue surplombante à travers la membrane, une autre structure filamenteuse apparaît perpendiculairement au fuseau. La plaque équatoriale se divise en deux. Pendant ce temps, le spermatozoïde s'arrondit et gagne le centre de la cellule.
- « Chacun des deux disques chromatiques fournit au globule polaire la moitié de sa substance... Ce n'est pas l'un des pôle du fuseau qui est éliminé mais c'est dans le plan équatorial que se fait l'élimination. » Van Beneden parle d'élimination car il suppose que la formation et l'expulsion du globule polaire est une élimination d'une partie du matériel de l'ovocyte. « L'on ne peut donc pas comparer la genèse du premier globule polaire à une division cellulaire ordinaire (mitose). Il ne s'accomplit d'ailleurs ici rien qui rappelle les transformations bien connues des filaments chromatiques d'un noyau de cellule au moment de la division d'un noyau et le corps protoplasmique de l'œuf parait n'intervenir en rien dans la constitution du globule polaire. »[5]

- « Aussitôt libéré, le premier globule polaire s’aplatit à la surface interne de la première couche périvitelline et, aux stades suivants, on le retrouve toujours adjacent à la face interne de cette zone... La substance chromatique du globule forment fréquemment deux amas qui, eux-mêmes, sont composés chacun de deux éléments accolés. Celle qui reste dans l'ovocyte est en tous points semblable à celle du globule polaire éliminé. »[5]
- « Peu après la pénétration du spermatozoïde, on voit apparaître autour du vitellus une zone homogène qui donne lieu tout d'abord à un double contour (la couche périvitelline) ; la zone s'épaissit rapidement ; elle atteint son maximum d'épaisseur quelque temps avant l'élimination du premier globule polaire. »[5]
Deuxième phase de maturation et expulsion du deuxième globule polaire
- « Aussitôt après l'élimination du premier globule polaire, il apparaît dans le vitellus, au voisinage immédiat de la surface, une nouvelle figure très compliquée qui, dans son ensemble, représente bien certainement le second fuseau de direction ou le second amphiaster de rebut. »[5] Van Beneden traduit ici les termes allemands créés par Otto Butschli (fuseau de direction) et Walther Fleming (amphiaster de rebut).
- « Les fragments des deux éléments chromatiques primitifs, quel que soit d'ailleurs leur nombre, se regroupent en deux disques adjacents dans des plans perpendiculaires à l'axe du fuseau, au voisinage de son équateur... Le second globule polaire, pas plus que le premier, ne se forme aux dépens d'un des pôles du fuseau... L'expulsion se fait dans le plan équatorial du fuseau et non pas par une de ses extrémités. »[5]
- Pendant cette phase de maturation, il se forme une seconde zone périvitelline entre le vitellus et la première couche périvitelline. « Quand cette couche atteint son complet développement, elle présente, sur le frais (sans fixation ni coloration), un aspect bien particulier : il semble qu'elle soit formée de fibrilles s'entrecroisant entre elles et fortement enchevêtrées... Le premier globule polaire se trouve toujours interposé entre les deux couches périvitellines. Le second globule polaire n'est éliminé qu'après la formation de la deuxième couche. »[5]
- Ce n'est que lorsque l'ovocyte se trouve dans la partie tout à fait inférieure de l'utérus que le second globule polaire se trouve constitué.

Formation du pronucleus mâle et fécondation
- « Dès le moment où l'œuf s'est débarrassé de son second globule polaire, le zoosperme engendre le pronucleus et, à ce même moment, le pronucleus femelle se constitue... Le pronucleus mâle vient compléter cette cellule réduite que j'appelle le gonocyte femelle et en faire une nouvelle cellule complète. La fécondation parait constituée essentiellement dans cette reconstitution de la première cellule embryonnaire. »[5]
- « Les deux pronuclei, sans se confondre, interviennent l'un et l'autre dans la formation d'une même figure dicentrique. L'un et l'autre subissent simultanément, et alors qu'ils sont parfaitement distincts, les mêmes changements qui se produisent dans un noyau de cellule ordinaire en voie de division karyokinétique (mitotique). Chaque pronucleus fournit deux anses à l'étoile chromatique du disque équatorial. L'étoile se constitue de deux anses mâles et de deux anses femelles. Chacune de celle-ci se divise, dans le sens de sa longueur, en deux moitiés dont l'une se rend à l'un et l'autre à l'autre noyau fille... L'étoile chromatique de la plaque équatoriale, fournie en partie par le pronucleus mâle, en partie par le pronucleus femelle, est identique à celle qui se forme dans les blastomères lorsqu'ils se divisent. De part et d'autre, l'étoile se constitue de quatre anses chromatiques. »[5]

Première division de l'œuf
Van Beneden observe minutieusement les étapes de la mitose. La segmentation commence. Il constate « que chacun de ces quatre chromosomes se fend longitudinalement, se dédouble, et que chaque moitié se rapprochant des pôles opposés de la figure mitotique, entre dans la constitution du noyau d'une des deux cellules filles. D'où la conclusion, par généralisation, que dans toutes les cellules nées de la segmentation de l'œuf, le noyau contient quatre chromosomes dont deux sont femelles et tirent leur origine de l'œuf, les deux autres étant mâles apportés par le spermatozoïde. »[2]
En 1887, il démontre que les mécanismes mis en évidence au cours de la première division de l'œuf se reproduisent à chaque division cellulaire ultérieure.
Étude en deux temps
En fait, van Beneden a fait son étude en deux temps. Il a d'abord procédé comme il l'a décrit en tête de son article : en examinant méthodiquement chaque section du tractus génital pour décrire les différentes phases de maturation. Il est arrivé jusqu'à la formation du zygote qui prend place dans le vagin. Mais, chez Ascaris, l'embryon ne se développe qu'à l'extérieur du corps de la femelle. Van Beneden n'a donc pas pu observer les premières étapes de la formation de l'embryon, ce qui était décevant. Hésitant à démarrer la culture des œufs et étant très occupé, il suspend ses recherches, jette les femelles Ascaris vivantes dans l'alcool dilué pour les conserver et rédige un article qu'il envoie à l'imprimeur en [5].
Ayant par la suite un peu de temps, il décide disséquer les vers mis en réserve dans l'alcool, et de les colorer. Surprise ! L'alcool avait pénétré lentement vers l'intérieur des tissus et pendant ce temps, les ovocytes et les ovules fécondés avaient poursuivi leur développement. Van Beneden enregistre les premières divisions de l'embryon et constate qu'à certaines étapes, les filaments chromatiques sont parfaitement visibles, ce qu'il n'avait pas observé dans la première phase de son étude. Il rédige une nouvelle partie et dessine de nouvelles planches (XXIII bis, XIX bis et XIX ter) pour son article dont la publication est retardée, ce qu'il peut se permettre puisqu'il est co-éditeur de la revue. L'article, daté de 1883, parait finalement fin [7].
Conclusion
Van Beneden observe donc l'expulsion d'un premier, puis d'un second globule polaire. Il constate que, si les corpuscules chromatiques sont également répartis à chaque division entre ovocyte et globule polaire, la répartition du cytoplasme est extrêmement inégale, l'ovocyte conservant la quasi-totalité du cytoplasme. Ces divisions cellulaires ne sont donc pas identiques à celles décrites en 1882 par Walther Flemming (mitoses), divisions dans lesquelles les deux cellules filles sont également pourvues en chromatine et en cytoplasme.
Il observe que, lors de la fécondation, le spermatozoïde fournit à l'ovocyte le lot de chromosomes manquant car son noyau n'a, lui aussi, que la moitié du nombre normal de chromosomes. « Dès lors, la signification de la fécondation s'éclaire : elle consiste dans le remplacement, par l'élément mâle, des chromosomes que l'œuf a rejeté en mûrissant. Contrairement à ce que l'on croyait généralement, sur la foi des travaux d'Hertwig, la fécondation n'est donc pas la copulation de deux noyaux de sexe différent, car ni l'œuf, ni le spermatozoïde n'ont un noyau complet. Les observations de van Beneden établissent pour la première fois, et de façon définitive, le lien causal qui unit deux phénomènes biologiques qui se succèdent immuablement chez tous les animaux : la maturation et la fécondation. L'élimination chromatique qui se fait pendant la première rend intelligible et nécessaire le remplacement qui se fait pendant la seconde ; ou, en d'autres termes, la maturation est la condition indispensable de la fécondation et elle est en même temps sa raison d'être. »[2]
Les recherches sur la maturation de l'œuf et le développement de l'ovaire des mammifères (Homme, Lapin) seront poursuivies par Hans de Winiwarter, élève de Van Beneden, et publiées en 1901. Cette étude précisera les étapes de la méiose et introduira les termes : leptotène, zygotène, pachytène et diplotène[7].
Centrosome
Van Beneden ne s'est pas intéressé qu'aux noyaux et aux chromosomes. Il a aussi décrit les autres structures qui apparaissent au moment de la mitose : aster, fuseau mitotique, corpuscule central (=centrosome). « C'est incontestablement Édouard van Beneden qui a donné la première description minutieuse et exacte de la sphère attractive et de son corpuscule central dans l'œuf ou les blastomères d'Ascaris en division. C'est lui qui, le premier, leur a attribué un rôle essentiel dans la division cellulaire et en a admis l'existence dans toute cellule, même au repos. Sans doute, bien avant qu'il n'étudia l'Ascaris, on avait vu, aux pôles du fuseau nucléaire, un corpuscule plus ou moins bien individualisé, et lui-même, en 1876, l'avait nettement signalé chez les Dicyémides. Mais en 1887, il va bien plus loin et, dans une légende remarquable, il démontre avec figures et photographies à l'appui, qu'une sphère persiste dans la cellule à côté du noyau au repos dans l'intervalle entre les mitoses ; qu'on doit par conséquent la considérer comme un organe permanent au même titre que le noyau ; que, de même que celui-ci se multiplie par division, toute sphère attractive procède aussi par division d'une sphère antérieure ; qu'elle est enfin le facteur déterminant de la karyo- et de la cytodiérèse. Quand une cellule va se diviser, cela est annoncé par la division de la sphère elle-même qui précède ainsi celle du noyau ; et même le corpuscule central, sinon la sphère tout entière, s'allonge, s'étrangle, et se coupe en deux avant l'achèvement de la mitose. »[2]
Conflit de paternité
Van Beneden tenait à la découverte du centrosome, de sa division en deux précédant de peu l'apparition des chromosomes et de son rôle dans l'organisation du fuseau mitotique. Pourtant Theodor Boveri lui a contesté la priorité de la découverte. En fait, il s'agit d'un phénomène découvert simultanément par deux chercheurs travaillant sur le même objet et le même sujet. Il semble que van Beneden ait eu une légère avance et en ait fait mention oralement dans une conférence dès . Boveri communique ses observations à la Société de Morphologie et de Physiologie de Munich le . Édouard van Beneden dépose son article le à l'Académie des sciences de Bruxelles qui le publie le suivant. Le travail de Boveri ne parait qu'en 1888[2].
En conclusion, si van Beneden a sans doute la priorité de la découverte du centrosome, de sa division et de son rôle dans la mitose, Boveri a démontré de façon incontestable que c'est le centrosome du spermatozoïde qui initie la première division mitotique de l'œuf et que le centrosome d'origine maternelle est éliminé[2].
Embryologie des mammifères
On peut dire que pendant toute sa vie, l'œuf ou l'embryon des Mammifères a occupé une partie de l'activité d'Édouard van Beneden. En 1875, puis en 1880, en 1884, en 1886 et 1888 paraissent une série de travaux et, en 1899, deux notes préliminaires ; ses mémoires posthumes, publiés en 1911 et 1912, sont le développement de ces dernières. Au total, 14 des 77 publications de van Beneden, collectées par Albert Brachet, (certaines signées avec son élève, Charles Julin) exposent ses recherches sur l'ontogenèse du Lapin ou de la Chauve-souris, pris comme modèles des Mammifères. Il faut y ajouter les publications remarquables d'un autre élève, Hans de Winiwarter, sur l'ovogenèse chez le Lapin, l'Homme et le Chat. Ces recherches ont été accomplies dans le laboratoire, à l'initiative et sous la supervision de van Beneden entre 1897 et 1901.
Van Beneden poursuit ses recherches sur les œufs et l'embryon des mammifères. Il met en exergue le processus de formation en trois feuillets fondamentaux de l'embryon : endo-, ecto- et mésoderme. Il décrit la gastrulation chez le lapin et la souris. Avec son élève Charles Julin, il étudie la formation des annexes du fœtus et du placenta. Avec Albert Brachet, il développe l'application des méthodes expérimentales à la biologie du développement.
Cérianthes
.jpg.webp)
ou anémone de mer

Van Beneden travaille beaucoup sur la morphologie et le développement des Cériantes. Il publie trois notes, une sur des larves voisines de la larve de Semper (1890) et deux sur les Arachnactis (1891). Il leur consacre une importante monographie dans les publications de la campagne océanographique allemande de 1889 : la Plankton-Expedition[11]. Il nomme trois nouvelles espèces : Apiactis, Peponactis et Solasteractis. Il voit dans les Cérianthes les ancêtres de tous les Chordés, une conception qui n'est plus admise aujourd'hui. Sur la base d'observations abondantes et méticuleuses, il projette de faire une publication importante avec l'esquisse d'un arbre phylogénétique. Il fait état de ces recherches dans ses cours mais repousse la publication. En 1907, il avoue : « On a déjà assigné tant d'ancêtres aux vertébrés et ils sont tous tombés dans l'oubli ; malgré toute ma conviction, j'hésite à en proposer un de plus. »[2] Après sa mort, on a retrouvé de nombreux dessins et planches qui avaient manifestement été préparés pour être publiés. Son élève, Paul Cerfontaine a rassemblé ces documents et les a publiés[12].
Tuniciers

Van Beneden et Julin s'appliquent à décrire la morphologie et le développement des Tuniciers. À la suite d'Alexandre Kovalevski, ils décrivent la ressemblance du développement de l'œuf des Tuniciers avec celles des Vertébrés. L'appartenance des Tuniciers à l'embranchement des Chordés est confirmée par la comparaison des séquences d'ADN[13].
« Dans la morphologie des Tuniciers, van Beneden et Julin, après avoir poursuivi le cell-lineage jusqu'à la gastrula, appliquent la même méthode au développement des feuillets et des organes : système nerveux, chorde, mésoblaste. Or ils constatent que dans la gastrula déjà, les cellules initiales de ces éléments, non seulement sont reconnaissables, mais affectent une disposition très caractéristique. Elles sont groupées au pourtour du blastopore en des sortes de cercles concentriques. Les cellules nerveuses et chordo-mésoblastiques futures forment deux anneaux périblastoporaux inscrits l'un dans l'autre ; le premier externe et ectoblastique, le second interne et endoblastique, ou tout au moins incorporé dans l'endoblaste. »[2]
À sa mort, van Beneden a laissé une abondante documentation issue de ses recherches sur les Tuniciers collectés au cours de l'expédition antarctique belge du S.Y. Belgica en 1897-1898-1899[14]. Son élève Marc de Sélys Longchamp les a publiées[15],[16].
Travaux divers
Il rédigea le compte-rendu sommaire des recherches entreprises à la station de biologie d'Ostende pendant les mois d'été 1883. Il y constata que le fond marin belge avait glissé et n'était pas constitué uniquement de sable. Dès lors des fonds de gravier se trouvaient à proximité de Westhinder où s'était développée une faune particulière.
Par la suite il mena également des recherches sur l'état de la faune près de Thorntonbak.
Les Polychètes, Échinodermes, les Ascidies et les poissons qui évoluaient à 29 mètres de profondeur ont été rencontrés ailleurs dans la mer du Nord. Ce constat lui a permis de travailler sur la relation entre la répartition et l'environnement.
Hommages
- Membre depuis 1872 de l'Académie Royale des Sciences, des Lettres et des Beaux-Arts de Belgique qu'il préside en 1902
- Prix quinquennal des Sciences Naturelles en 1877, 1887 et 1891
- Prix Serres de l'Institut de France
- Docteur Honoris Causa des Universités de Bruxelles, Cambridge, Oxford, Leipzig, Iéna, Edimbourg
- Membre correspondant de l'Institut de France, de l'Académie des Sciences de Berlin, de l'Académie des Sciences de Vienne, de l'Académie des Sciences de Saint Petersbourg
- Membre des Académies de Munich, Lisbonne, Bologne, Philadelphie, Copenhague, de l'Institut de Genève
- Membre d'honneur de la Société de Biologie de Paris, de la Société Impériale des Naturalistes de Moscou, de l'Institut Senkenberg à Francfort
- L'Institut de Zoologie de l'Université de Liège porte son nom.
- Une statue le représentant est dressée à côté de l'entrée de l'Institut.
- Le quai où se situe l'Institut porte également son nom.
Notes et références
- Gabriel Hamoir, La révolution évolutionniste en Belgique. Du fixiste Pierre-Joseph van Beneden à son fils darwiniste Édouard, Liège, Éditions de l'ULg (Université de Liège), , 193 p. (ISBN 978-2-930322-33-9, lire en ligne).
- Albert Brachet, « Notice sur Édouard van Beneden, membre de l'Académie », Annuaire de l'Académie Royale de Belgique, , p. 167-242 (lire en ligne).
- « Publications d'Édouard Van Benden », sur orbi.ulg.ac.be (consulté le )
- Édouard van Beneden, « La maturation de l'oeuf, la fécondation et les premières phases du développement embryonnaire des Mammifères, d'après des recherches faites chez le Lapin », Bulletins de l'Académie Royale des Sciences de Belgique. Vol. 40, 2e série, , p. 646-736
- Édouard van Beneden, « Recherches sur la maturation de l'oeuf et la fécondation », Archives de Biologie, 1883 vol. 4, p. 265-640 (lire en ligne)
- (de) Oskar Hertwig, « Beitrage zur Kenntniss der Bildung, Befruchtung und Theilung des Thierischen Eies (Contribution à la connaissance de la maturation, de la fécondation et de la division de l'oeuf animal) », Morphologiches Jarhbuch, 1,, , p. 347-434
- (en) Gabriel Hamoir, « The discovery of meiosis by E. van Beneden, a breakthrough in the morphological phase of fertility », International Journal of Developmental Biology, , p. 9-15
- Édouard van Beneden et Charles Julin, « L'appareil sexuel femelle de l'Ascaride mégalocéphale », Archives de Biologie, 1883 volume 4, p. 95-142 et 1 planche
- Édouard van Beneden, Recherches sur la maturation de l'oeuf, la fécondation et la division cellulaire, Gand, Leipzig & Paris, Librairie Clemm & G. Masson, , 421 pages et 13 planches (lire en ligne)
- Édouard van Beneden et Charles Julin, « La spermatogenèse chez Ascaris mégalocéphale », Bulletins de l'Académie Royale des Sciences de Bruxelles Vol. 7 n°4, , p. 312-342 (lire en ligne)
- Édouard van Beneden, Les Anthozoaires de la "Plankton-Expedition" avec 16 planches, une carte et 59 figures dans le texte, Kiel, Leipzig, Lipsius & Tischer, , 222 p. (lire en ligne)
- Édouard van Beneden & Paul Cerfontaine, « Travaux posthumes d'Édouard van Beneden sur les Cérianthaires », Archives de Biologie - Volume hors-série, , p. 1-242 (lire en ligne)
- (en) Frédéric Delsuc, Henner Brinkmann, Daniel Chourrout et Hervé Philippe, « Tunicates and not cephalochordates are the closest living relatives of vertebrates », Nature, vol. 439, no 7079, , p. 965-968 (ISSN 0028-0836 et 1476-4687, DOI 10.1038/nature04336, résumé, lire en ligne).
- « Voyage de la Belgica, 15 mois dans l'Antarctique », sur archive.org, (consulté le )
- Édouard van Beneden & Marc de Selys-Longchamps, Tuniciers : Caducichordata (Ascidiacés et Thaliacés), Anvers, J.-E. Buschmann, , 122 p. (lire en ligne)
- Édouard van Beneden & Marc de Selys-Longchamps, Tuniciers (2ème partie) : embryogenèse, Anvers, J.-E. Buschmann, , 57 p. (lire en ligne)
Voir aussi
Article externe
- Biographie
- Gabriel Hamoir, La découverte de la méiose et du centrosome par Edouard van Beneden, Édition Académie Royale de Belgique, 2010, 128 p.
Liens externes
 Portail du Brabant flamand
Portail du Brabant flamand  Portail de l’histoire de la zoologie et de la botanique
Portail de l’histoire de la zoologie et de la botanique  Portail de Liège
Portail de Liège  Portail de la biologie cellulaire et moléculaire
Portail de la biologie cellulaire et moléculaire