Tetrapoda
Les tétrapodes (Tetrapoda[alpha 1]) forment une super-classe d'animaux vertébrés dont le squelette comporte habituellement deux paires de membres et dont la respiration est normalement pulmonaire. Les tétrapodes ont évolué à partir d'un clade connu sous le nom de tétrapodomorphes qui, à leur tour, descendent des poissons à nageoires lobées apparu vers environ 390 millions d'années, durant le Dévonien moyen[3]. Les premiers représentants du groupe-couronne des tétrapodes (d'un point de vue traditionnel basé sur l'apomorphie) sont apparus au tout début du Carbonifère, il y a environ 350 millions d'années[4]. Les ancêtres aquatiques spécifiques des tétrapodes et le processus par lequel ils ont colonisé la terre terrestre après avoir émergé de l'eau restent flous. Le passage d'un schéma corporel pour respirer et naviguer dans l'eau à un schéma corporel permettant à l'animal de se déplacer sur terre est l'un des changements évolutifs les plus profonds connus[5],[6]. Les tétrapodes ont de nombreuses caractéristiques anatomiques et physiologiques distinctes de leurs ancêtres aquatiques. Ceux-ci comprennent la structure de la mâchoire et des dents pour se nourrir sur terre, les ceintures et les extrémités des membres pour la locomotion terrestre, les poumons pour la respiration dans l'air, et les yeux et les oreilles pour voir et entendre dans l'air.
« Tétrapode » redirige ici. Pour les structures destinées à résister aux vagues, voir Tétrapode (structure).
Ne doit pas être confondu avec Quadrupède.

| Règne | Animalia |
|---|---|
| Sous-règne | Bilateria |
| Infra-règne | Deuterostomia |
| Embranchement | Chordata |
| Sous-embr. | Vertebrata |
| Infra-embr. | Gnathostomata |
Classes de rang inférieur
- Batrachomorpha / Amphibia
- Plusieurs clades éteints
- Lissamphibia
- Reptiliomorpha / Pan-Amniota
- Plusieurs clades éteints
- Amniota
- Synapsida (Inclut les mammifères et leurs parents éteints)
- Sauropsida (inclut les reptiles et les oiseaux
Les premiers tétrapodes antérieurs au groupe-couronne étaient essentiellement aquatiques. Les amphibiens modernes, qui ont évolué à partir de groupes antérieurs, sont généralement semi-aquatiques : la première étape de leur vie est celle des têtards ressemblant à des poissons, et les étapes ultérieures sont en partie terrestres et en partie aquatiques. Cependant, la plupart des espèces de tétrapodes sont aujourd'hui des amniotes, dont la plupart sont des tétrapodes terrestres dont la branche a évolué à partir de tétrapodes antérieurs au début du Carbonifère supérieur. L'innovation clé chez les amniotes par rapport aux amphibiens est l'amnios, ce qui permet aux œufs de conserver leur contenu aqueux sur terre, plutôt que de devoir rester dans l'eau. Certains amniotes ont ensuite développé une fertilisation interne, bien que de nombreuses espèces aquatiques en dehors de l'arbre des tétrapode aient évolué avant l'apparition des tétrapodes, par exemple Materpiscis. Certains tétrapodes, tels que les serpents et les céciliens, ont perdu tout ou une partie de leurs membres par une spéciation et une évolution plus poussée. Certains n'ont caché que des ossements vestigiaux comme vestige des membres de leurs lointains ancêtres. D'autres sont redevenus amphibies ou vivent autrement des vies partiellement ou entièrement aquatiques, les premiers au cours de la période du Carbonifère[7], d'autres aussi récemment que le Cénozoïque[8],[9].
Les amniotes donneront naissance ultérieurement aux sauropsides (qui comprend les lépidosaures, les dinosaures, leurs descendants oiseaux, les crocodiliens, les tortues et les parents disparus) et les synapsides (qui comprend les mammifères et leurs parents disparus). Les amniotes comprennent les tétrapodes qui ont évolué pour le vol, comme les oiseaux parmi les dinosaures et les chauves-souris parmi les mammifères.
Définitions
Les tétrapodes peuvent être définis en cladistique comme l'ancêtre commun le plus proche de tous les amphibiens vivants (les lissamphibiens) et de tous les amniotes vivants (reptiles, oiseaux et mammifères), ainsi que de tous les descendants de cet ancêtre. Il s'agit d'une définition basée sur les nœuds (le nœud étant l'ancêtre commun le plus proche). Le taxon est ainsi défini en tant que groupe-couronne. Le terme tétrapodomorphe est utilisé pour la définition basée sur la tige : tout animal qui est plus étroitement lié aux amphibiens, reptiles, oiseaux et mammifères vivants qu'aux dipneustes. Le groupe ainsi défini est connu sous le nom de groupe total des tétrapodes[10].
La clade Stegocephalia est un groupe plus large équivalent à certaines utilisations plus larges du mot tétrapode, utilisé par les scientifiques qui préfèrent réserver les tétrapodes en tant que groupe-couronne, basé sur l'ancêtre commun le plus proche des formes vivantes[11]. Ces scientifiques utilisent le terme « proto-tétrapode » pour désigner les vertébrés ressemblant à des tétrapodes qui ne sont pas membres du groupe-couronne, y compris les poissons tétrapodomorphes[12]
Les deux sous-clades du groupe-couronne des tétrapodes sont Batrachomorpha et Reptiliomorpha. Les batrachomorphes sont tous des animaux partageant une ascendance commune plus récente avec des amphibiens vivants qu'avec les amniotes (reptiles, oiseaux et mammifères). Les reptiliomorphes sont tous des animaux partageant une ascendance commune plus récente avec des amniotes qu'avec des amphibiens vivants[13].
Caractéristiques
Morphologie
Les tétrapodes constituent un clade au sein des sarcoptérygiens, qui regroupe aujourd'hui deux taxons : les amphibiens et les amniotes (reptiles, oiseaux, mammifères). Ils sont caractérisés par la présence de deux paires de membres chiridiens (deux membres antérieurs ou scapulaires et deux membres postérieurs ou pelviens) qui sont homologues des nageoires paires des poissons.
Les membres chiridiens sont divisés en trois parties :
- stylopode : humérus sur le membre antérieur, fémur sur le membre postérieur ;
- zeugopode : radius/cubitus sur le membre antérieur, tibia/péroné sur le membre postérieur ;
- autopode : carpes-métacarpes-phalanges (membre antérieur), tarses-métatarses-phalanges (membre postérieur).
Ce schéma de base a subi de très nombreuses variations en relation avec l'adaptation à divers modes de locomotion des tétrapodes (vol, nage, saut, course, fouissage...).
Autres caractères dérivés propres :
- conduit lacrymal entre œil et sac nasal ;
- tête séparée du reste du corps par un cou ;
- os hyomandibulaire de la mâchoire passant dans les os de l'audition ;
- première vertèbre cervicale devenant l'atlas.
Place des tétrapodes dans le monde animal
| EXPLOSION RADIATIVE DES ANIMAUX | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
EUKARYOTES |
se nourrissent d'organismes dont ils sont dissociés | ||||||||||||||||
|
├─o Éponges |
hydres, méduses symétrie bilatérale | ||||||||||||||||
| PROTOSTOMIENS | DEUTÉROSTOMIENS | ||||||||||||||||
| LOPHOTROCHOZOAIRES : | notochorde | ||||||||||||||||
| vers plats vers à anneaux | |||||||||||||||||
| crâne squelette mâchoires squel. osseux émail | |||||||||||||||||
| double coquille reptation crâne, bec corné | |||||||||||||||||
| ECDYSOZOAIRES : | mues cuticulaires |
|
pattes et doigts sac amniotique | ||||||||||||||
| carapace externe | |||||||||||||||||
| chélicères |
|||||||||||||||||
| lézards, serpents | |||||||||||||||||
| araignées, scorpions | |||||||||||||||||
| mandibules | |||||||||||||||||
| OISEAUX | adaptation au vol | ||||||||||||||||
| crabes, langoustes | |||||||||||||||||
| 6 pattes, larves |
gl. sudoripares, néocortex | ||||||||||||||||
La définition des tétrapodes n'est pas facile. Entre les premiers sarcoptérygiens, poissons à nageoires charnues d'une part, et les tétrapodes ayant conquis l'autonomie terrestre (amniotes) d'autre part, il existe de nombreux types intermédiaires, dits amphibiens. Au sein des vertébrés, les tétrapodes se caractérisent par leur histoire évolutive : celle de la conquête du milieu terrestre.
Biodiversité
Dans la nature actuelle, les tétrapodes « sont représentés par plus de 33 000 espèces comprenant les amphibiens « modernes », ou lissamphibiens (apodes, salamandres, tritons, grenouilles, crapauds), et les amniotes (oiseaux, mammifères ainsi que chéloniens, squamates et crocodiliens, ces trois derniers groupes représentant ce que l’on appelait naguère les reptiles). Au cours de leur longue histoire de près de 400 millions d’années, certains tétrapodes, comme les serpents, ont perdu leurs membres locomoteurs et les ont compensés par l’adaptation à la reptation. D’autres, comme les cétacés, ont perdu leurs membres postérieurs pour mieux se déplacer dans l’eau. Les tétrapodes comptent également un grand nombre de groupes éteints, pour la plupart d’âge paléozoïque (540 à 250 millions d'années), qui étaient des tétrapodes basaux ou des formes primitives apparentées soit aux lissamphibiens (c’est le cas, probablement, des temnospondyles et certainement des lepospondyles), soit aux amniotes (anthracosaures, seymouriamorphes)[14] ».
| Groupe de Tétrapodes | Illustration | Classe phylogénétique | Nombre estimé d'espèces décrites |
Espèces menacées selon la liste rouge de l'UICN |
Pourcentage d'espèces décrites |
Meilleure estimation du pourcentage d'espèces menacées |
|---|---|---|---|---|---|---|
| Anamniotes Vertébrés amphibies pondant leurs œufs dans l'eau |
 |
Amphibiens | 7 302 | 1 957 | 88 % | 41 % |
| Amniotes Vertébrés terrestres et aériens pondant leurs œufs sur terre |
 |
Sauropsides (reptiles et oiseaux) |
20 463 | 2 300 | 75 % | 13 % |
.png.webp) |
Synapsides (mammifères) |
5 513 | 1 199 | 100 % | 26 % | |
| Total | 33 278 | 5 456 | 80 % | ? | ||
Arbre phylogénétique simplifié des tétrapodes
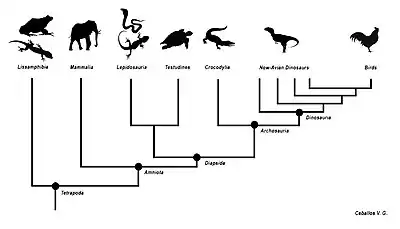
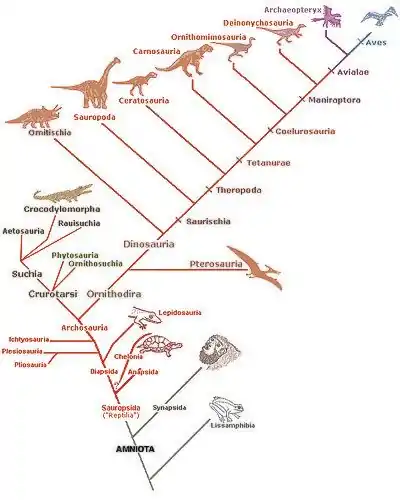
« Les archives fossiles, tout comme l'étude des vertébrés actuels, suggèrent que le tronc des tétrapodes s'est scindé très tôt en deux branches principales — les amphibiens et les amniotes (reptiles, oiseaux et mammifères)[16]. »
L'apparition de l'œuf amniotique va permettre une large radiation évolutive des tétrapodes, sans faire pour autant disparaître les tétrapodes non amniotiques (Lissamphibia). Tous les vertébrés terrestres actuels sont donc un groupe monophylétique descendant des premiers tétrapodes du Dévonien terminal, et étant donc considéré comme clade en cladistique[17]. La phylogénie ci-dessous représente une des trois hypothèses concurrentes, selon laquelle les lissamphibiens dérivent des temnospondyles[18]. Selon les deux autres hypothèses, les amphibiens actuels dérivent soit des lépospondyles[19], soit à la fois des temnospondyles et des lépospondyles (ils sont alors polyphylétiques)[20].
o TÉTRAPODES ├─o Temnospondyli (Arbre détaillé) │ └─o Lissamphibia (batraciens modernes - Arbre détaillé) │ ├─o Gymnophiones │ └─o │ ├─o Urodèles (tritons et salamandres) │ └─o Anoures (grenouilles et crapauds) └─o ├─o Lepospondyli (éteint - Arbre détaillé) └─o Reptiliomorpha └─o AMNIOTA (Arbre détaillé) ├─o Synapsida │ └─o Mammifères (Arbre détaillé) └─o Sauropsida ├─o Anapsida │ └─? Testudines (Tortues - Arbre détaillé) └─o Diapsida ├─o Ichthyosauria (éteint) └─o Sauria ├─o │ ├─o Plesiosauria (reptiles marins éteints) │ └─o Lepidosauria (Arbre détaillé) │ ├─o Rhynchocephalia (Sphenodon) │ └─o Squamata (serpents et lézards) └─o Archosauria ├─o Crocodilia (crocodiles - Arbre détaillé) └─o ├─o Pterosauria (reptiles volants éteints) └─o Dinosauria (dinosaures - Arbre détaillé) └─o Aves (oiseaux - Arbre détaillé)
Apparition de la respiration aérienne
La respiration pulmonaire, c'est-à-dire la capacité à respirer directement l'oxygène de l'air grâce à un poumon, (à l'origine plutôt un sac pulmonaire), est une caractéristique importante des tétrapodes, une de celle qui leur a permis de coloniser les milieux terrestres.
Cette adaptation est cependant plus ancienne que les tétrapodes, puisqu'elle remonte au-delà de 400 Ma, même si ceux-ci sont aujourd'hui presque les seuls vertébrés à avoir encore un ou des poumons.
Contraintes adaptatives
La respiration pulmonaire est au départ non pas une adaptation à la vie hors de l'eau, mais à une vie dans des milieux aquatiques pauvres en oxygène. « Pourquoi [...] certains poissons ont-ils évolué vers une respiration à l'air libre ? [...] La forme de leur crâne nous donne un premier indice : le crâne est aplati, ce qui suggère qu'ils vivaient dans des eaux peu profondes[21]. »
« Au Dévonien, certaines plantes se sont diversifiées et multipliées, et nombre d'entre elles ont acquis à cette époque des feuillages caducs [qui tombent de façon saisonnière]. Le bois et autres feuilles mortes produits par ces végétaux ont alors encombré, voire saturé, les milieux aquatiques peu profonds. [...] De surcroît, les eaux chaudes sont moins oxygénées que les eaux froides[21]. » Les eaux chaudes ont une capacité de dissolution de l'oxygène de l'air qui est plus faible, et la végétation pourrissante consomme (par oxydation) l'oxygène restant, un phénomène appelé eutrophisation.
Les eaux des marais chauds et encombrés de végétation pourrissante sont donc pauvres en oxygène. De nombreuses lignées de poissons vivant dans ces milieux ont donc développé des adaptations spécifiques pour capter directement l'oxygène de l'air. Il en va ainsi des anabantidés, qui ont développé un organe appelé labyrinthe. D'autres poissons « pipent » l'air en surface et le stockent dans leurs intestins, d'où l'oxygène passe dans le sang. Certains Sarcoptérygiens ont ainsi développé un organe spécifique, le poumon. Celui-ci est une exaptation dérivant de la vessie natatoire.
Les poumons primitifs des sarcoptérygiens, tels qu'ils existent encore chez les dipneustes ou les lissamphibiens modernes (amphibiens), diffèrent peu de ceux des reptiles ou de ceux des mammifères. Les côtes ne bougent en effet pas, et ne jouent aucun rôle lors de l'inspiration puis lors de l'expiration. C'est le mouvement de la bouche, plus précisément les mouvements du plancher buccal, qui envoient l'air dans les poumons[22], un fonctionnement appelé « pompe buccale ».
Date de l'apparition des poumons
Les sarcoptérygiens sont apparus vers la fin du Silurien (-444 à -416 Ma). Bien que probablement apparus en milieu marin, des lignées se spécialisent rapidement en eau douce. Les deux lignées principales que sont les actinistiens (dont les descendants sont les cœlacanthes) et les rhipidistiens (dont les descendants sont les tétrapodes et les dipneustes), divergent au début du Dévonien (-416 à -359 Ma)[23].
Aujourd'hui, les dipneustes sont des poissons, cousins des tétrapodes, qui ont des poumons (non-alvéolés) parfaitement fonctionnels au côté de branchies. La date de divergence d'avec la lignée qui mène aux tétrapodes étant supérieure à 400 Ma, l'apparition des poumons remonte donc au moins à cette date.
Par ailleurs, les cœlacanthes, redevenus purement marins, ont un organe vestigial qui est souvent considéré comme les restes d'un ancien poumon, aujourd'hui non fonctionnel. Si cette interprétation est la bonne, l'apparition des poumons, ou au moins de sacs pulmonaires primitifs, remonte encore quelques millions d'années plus tôt, avant la séparation entre les actinistiens et les rhipidistiens, c'est-à-dire tout au début du Dévonien.
Tétrapodomorphes
Après leur apparition à la fin du Silurien, les sarcoptérygiens ont connu un succès adaptatif important au Dévonien (-416 à -359 Ma). « Au cours de cet âge géologique [...], les poissons sarcoptérygiens dominaient toutes les mers, ne laissant guère qu'une portion congrue aux placodermes (poissons recouverts de plaques osseuses), aux acanthodiens (poissons à épines) et aux chondrichthyens (requins primitifs) [...][les] actinoptérygiens, les poissons tels que nous les connaissons, étaient alors très peu représentés[24] ».
Très présents également en eau douce (dont les sarcoptérygiens marins étaient au moins partiellement originaires), le groupe connaît différentes radiations évolutives. L'une des plus importantes, les rhipidistiens, va voir apparaître, après -400 Ma un groupe de poissons dit « tétrapodomorphes » qui annonce déjà les tétrapodes par certaines de leurs évolutions osseuses, en particulier au niveau des nageoires, dont certaines caractéristiques commencent à annoncer les pattes des premiers tétrapodes.
Ci-dessous un arbre généalogique simplifié des sarcoptérygiens (tous les taxons connus n'y apparaissent pas). On y voit la parenté étroite entre les dipneuste et les poissons tétrapodomorphes.
Sarcoptérygiens |--+--Onychodontiformes | `--Actinistia (Cœlacanthes) `--Rhipidistia (dipneustes et tétrapodes) |--Dipnomorpha (dipneustes) `--Tetrapodomorphes |--Rhizodontiformes `--Osteolepiformes |--Tristichopteridae | |--Eusthenopteron | `--+--Cabonnichthys | `--Mandageria `--Elpistostegalia |--Panderichthys `--TETRAPODES
Évolution des nageoires
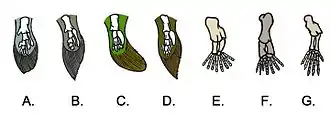

Chez les poissons les membres pairs, c'est-à-dire les nageoires pectorales et abdominales, consistent en lames aplaties et élargies, soutenues par une tige squelettique plurisegmentée mais non articulée, portant latéralement une série de rayons. Ces membres que les biologistes appellent pterygium ont des structures variées qui donnent le nom aux principaux groupes de poissons (-ptérygiens). Les membres chiridiens (appendices locomoteurs pairs et articulés, polydactyles puis pentadactyles[25], caractère ancestral plésiomorphique issu du membre chiridien[26]) des Vertébrés tétrapodes est homologue au pterygium des poissons, la principale différence étant que ses segments sont unis par de nombreuses articulations mobiles. La transformation des nageoires des ancêtres aquatiques des ostéolepiformes des Tétrapodes en membres chiridiens permet la mise en place de la quadrupédie, mode de locomotion terrestre initial largement privilégié[27].
Des nageoires lobées charnues, soutenues sur les os plutôt que les nageoires renforcées des raies, semblent avoir été un trait original des poissons osseux (Osteichthyes), groupe au sein duquel se sont individualisés les sarcoptérygiens. Le groupe le plus primitif avec des nageoires de raie, les bichirs, possède toujours des nageoires frontales charnues.
Les tétrapodes primitifs et leur membre chiridien se sont développés à partir d'un tétrapodomorphe, avec un cerveau à deux lobes dans un crâne aplati, une bouche large et un museau court, et dont les yeux dirigés vers le haut indique qu'il vivait sur le fond. Ses nageoires pectorales et pelviennes monobasales (un seul os s'articule avec la ceinture[28]) montraient déjà des signes d'adaptation avec des bases et des os charnus.
Un tétrapodomorphe encore plus proche des premiers tétrapodes était Panderichthys (-385 Ma), qui possédait même des choanes. Ce poisson utilisait ses nageoires comme des pagaies, car il vivait dans des eaux peu profondes envahies de plantes et de détritus. Il est possible qu'il ait utilisé ses nageoires pour s'accrocher aux plantes ou à autre chose pendant qu'il se tenait en embuscade attendant ses proies. Panderichthys est un Elpistostegalia, comme les tétrapodes, mais il en est plus un cousin qu'un ancêtre direct.
Les tétrapodomorphes vivent dans tous les milieux : eau douce, eau saumâtre (estuaires) et eau salée, et aussi bien dans le grand continent du sud à l'époque, le Gondwana, que le petit continent du nord, l'Euramérique.
Au dévonien supérieur (-383 à -359 Ma), les tétrapodes encore aquatiques semblent avoir surtout vécu dans des eaux saumâtres (estuaires, marais côtiers). L'analyse isotopiques de « fossiles des premiers tétrapodes et de leur faune associée, provenant du Groenland et de Chine [... a] démontré que ces tétrapodes vivaient dans des eaux saumâtres. [...] Les premiers tétrapodes étaient donc capables de tolérer une grande gamme de salinités, à l’instar de nombreux poissons actuels vivant dans ces milieux, comme les anguilles ou les saumons[29] ». Cette capacité à occuper des milieux très diversifiés auraient facilité leur rapide dispersion sur tous les continents, prélude à la terrestrialisation de certaines lignées.

Toujours vers -385 Ma, on connaît également le genre Eusthenopteron, qui montre des formes de dents et d'os du crâne typique des tétrapodes postérieurs, ainsi que des choanes.
Premiers tétrapodes

L'apparition de proto-pattes est le principal signe du passage des tétrapodomorphes comme Panderichthys (-385 Ma) aux premiers véritables tétrapodes, encore totalement aquatiques.
Contrairement à une idée reçue courante, les membres chiridiens sont d'abord apparus chez des organismes entièrement aquatiques. Les premières pattes fossiles apparaissent ainsi comme des sortes de pagaies, bien adaptées au déplacement (écarter, agripper) dans l'encombrement végétal des rives immergées et des marécages.
Secondairement, ces « pattes » aquatiques ont été réutilisées pour permettre le déplacement hors de l'eau, comme le montre l'étude des deux espèces de tétrapodes fossiles, Acanthostega et Ichthyostega.
Les caractéristiques universelles des tétrapodes : membres antérieurs tournés vers l'arrière en direction du coude et membres postérieurs tournés vers l'avant en direction du genou, peuvent être déjà repérées chez les premiers tétrapodes qui vivaient en eau peu profonde.
Neuf genres de tétrapodes dévoniens ont été décrits, plusieurs ne sont connus, en totalité ou en partie, que par ce qui reste de la mâchoire inférieure. Tous vivaient dans le supercontinent nord-américain-européen, qui comprenait l'Europe, l'Amérique du Nord et le Groenland. La seule exception est un genre unique du Gondwana, Metaxygnathus, qu'on a trouvé en Australie.
Le premier tétrapode dévonien asiatique identifié a été reconnu à partir d'une mâchoire fossile décrite en 2002. Le tétrapode chinois Sinostega pani a été découvert au milieu de plantes tropicales fossilisées et de sarcoptérygiens dans les sédiments de grès rouges du Ningxia Hui, région autonome de la Chine du nord-ouest. Cette découverte a considérablement étendu la distribution géographique de ces animaux et a soulevé de nouvelles questions sur leur distribution dans le monde et sur la grande diversité taxinomique à laquelle ils ont abouti en un temps relativement court.
À l'époque du dévonien supérieur, les plantes terrestres avaient stabilisé les habitats d'eau douce, permettant ainsi de se développer aux premiers écosystèmes de zone humide, avec des réseaux alimentaires de plus en plus complexes qui offraient de nouvelles possibilités. Les habitats d'eau douce n'étaient pas les seuls endroits où l'on pouvait trouver de l'eau remplie de matières organiques en liaison avec une végétation dense près du bord de l'eau. Les habitats marécageux comme les zones humides peu profondes, les lagunes côtières et les grands deltas salés des fleuves existaient aussi à cette époque, et bien des spécialistes suggèrent que c'est dans un tel environnement que les tétrapodes ont évolué. Des fossiles des plus anciens tétrapodes ont été découverts dans les sédiments marins et puisqu'on en a retrouvé tout autour du monde, ils doivent avoir fait leur extension en suivant les lignes côtières - ils ne sauraient donc n'avoir vécu que dans l'eau douce.
Pour trancher le débat sur la dominante eau douce, eau salée ou eau saumâtre des premiers tétrapodes aquatiques, l'équipe de Jean Goedert et Christophe Lécuyer a publié dans la revue Nature du un article[30] ou elle analyse les rapports entre isotopes de l’oxygène, du soufre et du carbone de fossiles, sachant que ces rapports varient en fonction de la salinité du milieu. Leur conclusion est que « les tétrapodes dévoniens étaient des animaux adaptés à des milieux de salinité variable où se trouvent des proies marines, en d’autres termes aux estuaires, aux lagunes et autres marais salés que l’on rencontre sur les côtes partout sur la planète[31] ». Le « rapport isotopique de l’oxygène 18 [...] s’avère bien plus bas que celui de l’eau de mer très oxygénée, ce qui suggère un environnement pauvre en oxygène[31] ». Si la mer n'était pas le milieu dominant des tétrapodes aquatiques, la tolérance aux milieux salés leur a cependant permis une extension rapide sur la planète.
Tétrapodes aquatiques

L'apparition des tétrapodes aquatiques date de « la fin du dévonien moyen, dans une fourchette étroite comprise entre 375 et 380 Ma[21] ». Tous les scientifiques ne sont pas d'accord avec cette ancienneté, non pas pour des incertitudes de datation, mais plutôt pour des raisons de définition de ce qui fonde la différence entre un tétrapodomorphe et un tétrapode. Certains voient ainsi plutôt l'apparition des tétrapodes vers -370 Ma, avec l'apparition des premiers humérus[32].
On suppose traditionnellement que les premiers tétrapodes ont évolué dans des habitats d'eau douce, peu profonds et marécageux.
Un des plus anciens fossiles connus proches des tétrapodes est celui de Tiktaalik roseae, daté de 380 à 375 Ma[33]
À côté d'attributs caractéristiques des poissons osseux (écailles, nageoires pectorale et pelvienne munis de rayons), les robustes nageoires pectorales de Tiktaalik étaient rigidifiées par un squelette osseux articulé. « Sa morphologie est similaire à celle des poissons mais l’articulation de ses nageoires pectorales suggère que le Tiktaalik pouvait supporter le poids de son corps. L’épaule et le coude sont flexibles, contrairement aux poissons, expliquent les chercheurs dans la revue Nature[33] ». Ces caractéristiques anatomiques, qui devaient lui permettre d'émerger ponctuellement de l'eau, en font un possible maillon évolutif entre les poissons (comme Panderichthys) et les tétrapodes plus affirmés (comme Acanthostega).
Quelques scientifiques le considèrent donc comme un tétrapode très primitif, quand d'autres le voient encore comme un tétrapodomorphe, simplement plus avancé que Panderichthys (-385 Ma) sur la voie de l'apparition des pattes. Un de ses découvreurs, Neil Shubin, l'a même qualifié de fishapod, contraction de fish (poisson) et tétrapode[34].
Datant de quelques millions d'années après Tiktaalik, « le plus ancien humérus fossile au monde aurait été découvert par des chercheurs [...]. Vieux de plus de 370 Ma, ce dernier offrirait la preuve aux scientifiques que les membres ont d’abord évolué dans l’eau [...]. Après l’analyse de ce fossile, Neil Shubin et ses collègues de l’Université de Chicago (États-Unis) ont constaté que les caractéristiques que les paléontologues attribuent d’habitude à la vie dans le monde terrestre sont plutôt d’un environnement aquatique. Cet humérus appartenant à une nageoire aurait permis, selon eux, à l’animal supporter le corps et, d’autre part l’aurait aidé à relever la tête[32] », peut-être pour la sortir de l'eau afin de respirer l'air.

Vers -363 Ma, Acanthostega est un tétrapode purement aquatique, dont les membres ont huit doigts, mais déjà assez évolués, avec ses quatre pattes nageuses, qui devaient lui permettre de se déplacer en agrippant la dense végétation aquatique des berges. Acanthostega est assez tardif, puisqu'il date de peu avant la grande disparition des tétrapodes du Dévonien terminal. Mais il est un bon modèle des tétrapodes aquatiques. Il est possible qu'il ait utilisé ses pattes de devant afin de prendre appui sur un support (branches immergées, fonds peu profonds), afin de hisser sa lourde tête hors de l'eau pour mieux respirer, mais le dessin de ses côtes et de ses pattes exclut une sortie de l'eau[24]. En effet, « les membres d'Acanthostega étaient trop faibles et mal orientés pour lui permettre de marcher sur la terre ferme[35] »

Vers la même époque qu'Acanthostega, Tulerpeton (-363 Ma) est un autre tétrapode aquatique reconstitué par un squelette fragmentaire retrouvé à Tula, en Russie. Les pattes, épaules antérieures et une partie du museau ont seuls été retrouvés. L'animal avait 6 doigts[36], et ses membres ressemblent plus à des pagaies aquatiques qu'à des pattes marcheuses. Néanmoins, les doigts sont relativement longs et fins, ce qui suggère un habitat plus terrestre que celui d’Acanthostega. Les os de l'épaule sont eux aussi plus robustes, ce qui aurait pu faciliter également des sorties de l'eau. Enfin, s'il est risqué de définir précisément le mode de vie de Tulerpeton, les ossements retrouvés semblent ceux d'un animal à dominante aquatique, quoique pouvant mieux sortir de l'eau qu’Acanthostega.
Premières sorties de l'eau

Les premières sorties de l'eau connues dateraient du Dévonien supérieur, vers -365 Ma, avec Ichthyostega, « le plus ancien des vertébrés connus présentant des adaptations pour une locomotion autre que la nage[21] ».
Ichthyostega n'est pas un tétrapode pleinement terrestre, mais il montre de nouvelles adaptations à la vie hors de l'eau. En effet, après l'apparition des poumons chez certains Sarcoptérygiens, de choanes chez les tétrapodomorphes et de pattes/nageoires chez les premiers tétrapodes aquatiques, Ichthyostega a trois adaptations majeures : des pattes antérieures capables de supporter son poids hors de l'eau, des cotes allongées évitant l'écrasement des poumons par le poids du corps hors de l'eau, et « une chaine d'os descendant jusqu'au milieu de la poitrine. Jennifer Clark, une des auteurs de l’étude, précise dans la revue Nature "que ces os au niveau de la poitrine constituent la première apparition, dans l'évolution, d'un sternum corporel. Une telle structure devait renforcer la cage thoracique d'Ichthyostega, lui permettant ainsi de soutenir une partie du poids de son corps au niveau de la poitrine pendant ses mouvements sur terre" [37]».
Ces 3 évolutions permettent ainsi à l'animal de respirer avec son corps hors de l'eau, alors que les précédents tétrapodes aquatiques comme Acanthostega ne pouvaient pas sortir de l'eau.
À ce titre, « Ichthyostega est le plus ancien tétrapode connu adapté à la fois à la nage et à une forme de locomotion terrestre. Il semble qu'il se déplaçait à terre comme un phoque, c'est-à-dire en projetant en avant ses pattes antérieures et sa tête avant de traîner la partie arrière de son corps. Manifestement, l'arrière du corps, avec sa large queue capable d'onduler et ses membres postérieurs en forme de pagaie, avait une vocation aquatique[21] ».
Il est probable qu'Ichthyostega ne venait que très partiellement à terre. D'une part, son corps est mal adapté à la vie terrestre, avec une mobilité à terre assez réduite et, d'autre part, sa taille pouvant atteindre 1,35 mètre devait rendre difficile de trouver de la nourriture sur terre. À cette époque-là, d'ailleurs, celle-ci était surtout peuplée d'arthropodes. Il est possible que les jeunes se soient nourris de ces derniers, mais ceux-ci ne pouvaient sans doute pas suffire à nourrir un adulte, par ailleurs très lent à terre. Le mode de vie de l'animal est inconnu, et on ne peut donc que supposer les raisons adaptatives de la venue à terre. Celle-ci était peut-être motivée par le besoin d'un refuge contre les prédateurs.

Si le squelette de Ichthyostega est assez bien connu, d'autres tétrapodes du Dévonien terminal (entre -365 et -360 Ma) sont connus par des fossiles plus lacunaire, comme Hynerpeton.
Conquête du milieu terrestre au Carbonifère

La raison pour laquelle les tétrapodes ont commencé à s'installer sur la terre ferme est toujours un sujet de discussion. Mais si elle est encore hésitante au Dévonien tardif (-360 Ma), l'adaptation au milieu terrestre devient très affirmée au cours du Carbonifère (-359 à 299 Ma).
Lacune de Romer
À partir de -360 Ma commence la période connue sous le nom de « lacune de Romer[38] ». Celle-ci dure une vingtaine de millions d'années, et est marquée par une disparition des fossiles de tétrapodes. « En général, le phénomène est attribué à une nouvelle crise globale, connue sous le nom de crise du Dévonien-Carbonifère[24] ».
La cause de l'extinction pourrait cependant être plus spécifique. La dérive des continents a en effet fortement rapproché les paléo-continents du Gondwana et de l'Euramérique. La grande majorité des tétrapodes connus vivaient sur ce dernier continent. « Des poissons tétrapodomorphes sont entrés en compétition avec les tétrapodes euraméricains dès que le Gondwana s'est rapproché assez de l'Euramérique[24] ». Pour les tétrapodes de l'époque, encore totalement (Acanthostega) ou largement (Ichtyostega) aquatiques, la compétition semble avoir été trop rude, puisque leurs lignées disparaissent. « Ainsi, il semble qu'aucun des groupes de tétrapodes connus au Dévonien n'ait dépassé la limite séparant le Dévonien et le Carbonifère. [...] Un irréductible petit groupe a toutefois subsisté dans quelques niches écologiques, où les gros poissons prédateurs ne s'aventuraient pas. S'agissait-il de la partie amont des réseaux fluviaux encombrés de plantes et de bois[24] ? ».
« Les rares tétrapodes peuplant la lacune de Romer ressemblent aux tétrapodes postérieurs à cette lacune, qui sont d'un type bien différent de ceux du Dévonien[24] ». Les animaux comme Ichthyostega ou Acanthostega ne sont ainsi pas les ancêtres des tétrapodes postérieurs. Une lignée encore inconnue a manifestement survécu à l'extinction des tétrapodes il y a 360 Ma, mais a mis quelque 20 millions d'années pour s'adapter pleinement à son nouveau milieu terrestre, et pour s'y répandre.
Depuis sa mise en évidence par Romer en 1956, cette lacune a été progressivement comblée avec les découvertes de tétrapodes précoces du Carbonifère tels que Pederpes[39],[40] et Crassigyrinus
Au Dévonien, certains poissons à nageoires lobées comme l'Eusthenopteron, bien que dotés déjà de nageoires dont le squelette interne évoque celui des tétrapodes, restent pélagiques ou strictement aquatiques. Des poissons tétrapodomorphes connaissent une diversification importante mais restent inféodés à des environnements côtiers (estuaires, lagunes) et d'eau douce (fleuves, rivières, mares), tout en présentant plusieurs adaptations qui leur permettent de se déplacer épisodiquement sur terre : Panderichthys rhombolepis, Tiktaalik, Acanthostega, Ichthyostega. |

Raisons de l'évolution
Le ou les groupes restreints qui ont survécu à l'extinction de la fin du Dévonien ont progressivement acquis des adaptations beaucoup plus fortes à la vie terrestre, en particulier quatre pattes pleinement marcheuses, ce que n'avait pas Ichtyostega.
Une première raison pourrait être la forte concurrence des poissons tétrapodomorphes aquatiques, qui en occupant les milieux originels des tétrapodes euraméricains auraient forcé la sélection des variants les plus adaptés au milieu terrestre, dénué de concurrents.
Il a été posé comme hypothèse que les jeunes auraient été parmi les premiers à coloniser ce milieu, les adultes étant plus aquatiques faute de nourriture suffisante à terre. Si les milieux terrestres semblent en effet avoir été pauvres en nourriture pour de gros animaux, les insectes qui les peuplaient pouvaient plus facilement convenir à des jeunes de petite taille.
De plus, les jeunes sont aussi les plus vulnérables, et donc les plus à même de bénéficier d'un refuge terrestre. Même sans sortie intégrale de l'eau, les eaux peu profondes près de la terre ferme offraient aussi aux jeunes un refuge. On peut d'ailleurs remarquer que les jeunes poissons actuels (et les amphibiens) passent souvent la première partie de leur vie dans la sécurité relative d'eaux peu profondes, comme les rives, les roseaux ou les mangroves, où les gros prédateurs ne peuvent accéder.
Finalement, la terre ferme, avec son absence totale de prédateurs, a sans doute offert des opportunités pour des animaux qui avaient développé des adaptations (poumons et pattes) spécifiques à des milieux aquatiques particuliers, mais réutilisables à terre, un phénomène connu en théorie de l'évolution sous le nom d'exaptation.
Quatre pattes marcheuses
L'un des premiers tétrapodes bien conservé et parfaitement adapté à la vie terrestre date ainsi de -345 Ma[41], juste un peu avant la fin de la « lacune de Romer », qu'il contribue à combler. Le Pederpes finneyae était une « créature, munie de dents, de quatre pattes et estimée longue de 90 centimètres. [...] Pederpes finneyae pouvait marcher sur la terre ferme, mais également évoluer dans l'eau[35] ». « Il ressemblait à un crocodile un peu disgracieux[41] ».
S'il n'est pas totalement terrestre, l'animal n'en est pas moins bien adapté à la vie sur terre, même s'il se reproduisait encore certainement dans l'eau. Ses quatre pattes marcheuses (contre seulement deux pattes marcheuses chez Ichtyostega) sont pleinement fonctionnelles. Elles ont déjà 5 doigts, une caractéristique propres aux tétrapodes modernes, alors que Acanthostega avait huit doigts, et que Ichthyostega en avait sept.
Caractères ancestraux et autres adaptations

Les tétrapodes ancestraux ne sont déjà plus tout à fait des poissons, mais ne sont pas encore complètement indépendants du milieu aquatique. À ce titre, ils conservent encore des branchies au côté de leurs poumons, et ce même à l'âge adulte. La disparition (ou la quasi-disparition) des branchies à l'âge adulte n'est pas bien documentée, mais date forcément du Carbonifère, durant lequel apparaissent les reptiles[42].
Au-delà des adaptations les plus visibles : poumons, pattes/nageoires puis pattes marcheuses, choanes, côtes protégeant les poumons de l'écrasement par le poids du corps, les tétrapodes ont aussi acquis des adaptations essentielles à la vie dans un milieu aérien (même si celui-ci n'est que partiel au début) :
- le cou devient articulé : l'attache des membres antérieurs recule, se désolidarisant de la tête. Les premières vertèbres se spécialisent pour permettre des mouvements du crâne indépendants du tronc ;
- l'oreille (qui était primitivement consacrée à l'équilibre) se spécialise pour permettre également l'audition dans l'air ;
- le développement du museau et le recul des orbites oculaires favorisent l'olfaction terrestre[43]. L'agrandissement des yeux, capables d'accommodation, est également une adaptation au milieu aérien[44]. Ils se couvrent d'une paupière protectrice, et un liquide lacrymal assure leur maintien en milieu humide ;
- les membres se renforcent, et permettent des mouvements performants en milieu aérien.
La conquête du milieu terrestre est donc bien amorcée dès la première moitié du carbonifère (-340 Ma), mais les tétrapodes primitifs conservent alors un mode de vie proche de celui des actuels batraciens, avec une forte dépendance aux milieux aquatiques, en particulier pour la reproduction (ponte des œufs). Malgré certaines ressemblances dans les modes de vie (partiellement aquatiques), de ponte (œufs aquatiques) ou de respiration (branchies, au moins chez les jeunes), ces tétrapodes primitifs ne sont cependant pas des amphibiens au sens moderne du terme. Le taxon lissamphibia est en effet d'apparition plus tardive, et porte des évolutions qui ne sont pas encore présentes chez les premiers tétrapodes. En ce sens, les batraciens actuels ne sont pas des tétrapodes primitifs, ni même n'en sont des approximations, même s'ils ont souvent conservé des caractères partiellement aquatiques.
Apparition de l'œuf amniotique

La fonctionnalité décisive qui permettra aux quadrupèdes de s'émanciper du milieu aquatique est l'invention de l'œuf amniotique, qui incorpore à l'œuf un micro-milieu aquatique nécessaire au développement de l'embryon.
Cet œuf à coquille, avec une poche liquide à l'intérieur, protège l'embryon du dessèchement, alors même que l'œuf est pondu dans un milieu aérien. Cette évolution n'est pas la seule à signer l'apparition des premiers amniotes. Elle s'accompagne aussi d'une peau écailleuse sèche et relativement étanche, qui ralentit beaucoup la perte d'humidité à travers la peau semi-perméable des premiers tétrapodes amphibiens.
L'œuf amniotique à coquille et la peau étanche des premiers reptiles vont très fortement augmenter l'indépendance aux milieux aquatiques des tétrapodes, et permettront l'explosion radiative des amniotes.
Ces deux évolutions laissant peu de traces paléontologiques, l'apparition des premiers reptiles est quelque peu difficile à dater, mais elle est généralement estimée à -320 Ma.

Westlothiana lizziae, découvert en 1989 dans un niveau de -338 Ma[45], soit juste à la fin de la lacune de Romer, est un animal de 20 cm de long, qui montre de nettes évolutions osseuses vers les reptiles[46], mais il est généralement considéré comme une forme intermédiaire plus proche des batraciens primitifs que des véritables amniotes, ou au moins comme une espèce indéterminée dans sa classification[47].
Vers -315 Ma, Hylonomus lyelli est le premier animal identifié pour lequel existe un consensus scientifique sur le fait qu'il était un reptile pondant des œufs amniotiques[48].
À partir de cette époque, les sauropsides vont se diversifier, donnant naissance à de nombreux groupes : archosauriens (parmi lesquels les actuels crocodiliens et oiseaux, mais aussi les anciens dinosaures ou ptérosauriens), tortues, lézards et les serpents.

Un autre groupes majeurs d'amniotes, les synapsides (souvent appelés reptiles mammaliens en raison de leurs apparences mais ce taxon ne regroupe en aucun cas des reptiles), seront les premiers représentant de ce clade à se diversifier. L'un des sous-groupes, les thérapsides, vont donner naissance aux mammifères, faisant de ces animaux, les seuls représentants actuels de cette lignée, tous les autres groupes étant désormais éteints.
Les tétrapodes sans œufs amniotiques n'ont cependant pas disparu, donnant naissance aux trois grands groupes modernes de Lissamphibia (batraciens) que sont les gymnophiones (ou apodes), les urodèles et les anoures.
Ammoniaque et urée
L'ancêtre commun de tous les gnathostomes actuels vivait dans l'eau douce et c'est plus tard qu'il est retourné vers la mer. Pour lutter contre la salinité beaucoup plus haute de l'eau marine, ses descendants ont acquis la capacité de transformer les déchets ammoniacaux en urée inoffensive, et de conserver cette dernière dans le corps afin de rendre le sang aussi salé que l'eau marine, sans empoisonner l'organisme.
Par la suite les actinoptérygiens sont revenus à l'eau douce et ont perdu cette possibilité. Comme leur sang contenait plus de sel que l'eau douce, ils pouvaient simplement se débarrasser de l'ammoniaque par leurs branchies. Et quand, une nouvelle fois, ils sont finalement revenus à la mer, il ne leur a pas été possible de récupérer le vieux système qui consistait à transformer l'ammoniaque en urée et il leur a fallu à la place élaborer des glandes qui excrétaient le sel.
Les dipneustes font de même: quand ils vivent dans l'eau, ils produisent de l'ammoniaque et pas d'urée ; mais quand l'eau s'assèche et qu'ils sont forcés de se creuser un abri dans la boue, ils recommencent à produire de l'urée. À la manière des poissons cartilagineux, le cœlacanthe peut conserver l'urée dans son sang, comme le peuvent les seuls amphibiens que l'on connaisse capables de vivre pendant de longues périodes dans l'eau salée (le crapaud Rhinella marina et la grenouille Fejervarya cancrivora). Il s'agit de traits qu'ils ont hérité de leurs ancêtres.
Si les premiers tétrapodes avaient vécu dans l'eau douce, ils auraient perdu la capacité de produire de l'urée et n'auraient plus utilisé que l'ammoniaque, ils auraient donc dû par la suite élaborer de nouveau cette capacité perdue à partir de zéro. Or, de tous les actinoptérygiens actuels, pas une seule espèce n'a été capable de le faire, il n'est donc pas probable que les tétrapodes l'auraient fait.
S'ils ne pouvaient produire que de l'ammoniaque les animaux terrestres seraient obligés de boire sans arrêt, ce qui rendrait impossible la vie sur la terre ferme (quelques exceptions existent, comme certains cloportes terrestres qui peuvent excréter leurs déchets azotés sous forme de gaz d'ammoniaque). Cela a dû probablement poser problème au début, au moment où les tétrapodes ont commencé à vivre une partie du temps hors de l'eau, mais finalement c'est le système de l'urée qui l'a complètement emporté. Pour cette raison il est fort douteux qu'ils soient venus de l'eau douce (à moins qu'ils n'aient d'abord émigré dans les habitats d'eau douce et soient ensuite passés sur la terre ferme au cours d'une deuxième émigration suivant de peu la précédente, alors qu'ils n'avaient pas encore oublié comment faire de l'urée), même si certains qui ne sont jamais allés sur la terre ferme (ou des espèces primitives éteintes qui sont revenues à la vie aquatique) pourraient évidemment s'être adaptés aux lacs et aux rivières d'eau douce.
Notes et références
Notes
- Du grec ancien τετρα / tetra « quatre » et πούς / poús « pied », pour donner littéralement « quatre pieds ».
Références
- (de) B. Hatschek et C. J. Cori, Elementarcus der Zootomie in fünfzen Vorlesungen [« Elementary Zootomy in Fifteen Lectures »], Jena, Gustav Fischer, (lire en ligne)
- (en) Phylonyms: A Companion to the PhyloCode, Boca Raton, CRC Press, , 759–764 p. (ISBN 978-1-138-33293-5), « Tetrapoda B. Hatschek and C. J. Cori 1896 [M. Laurin], converted clade name »
- (en) Katarzyna Narkiewicz et Marek Narkiewicz, « The age of the oldest tetrapod tracks from Zachełmie, Poland », Lethaia, vol. 48, no 1, , p. 10–12 (ISSN 0024-1164, DOI 10.1111/let.12083)
- (en) I. Irisarri, D. Baurain et H. Brinkmann, « Phylotranscriptomic consolidation of the jawed vertebrate timetree. », Nat Ecol E, vol. 1, , p. 1370–1378 (DOI 10.1038/s41559-017-0240-5)
- (en) Long JA, Gordon MS, « The greatest step in vertebrate history: a paleobiological review of the fish-tetrapod transition », Physiol. Biochem. Zool., vol. 77, no 5, sep–oct 2004, p. 700–19 (PMID 15547790, DOI 10.1086/425183, S2CID 1260442)
- (en) N. Shubin, Your Inner Fish: A Journey Into the 3.5-Billion-Year History of the Human Body, New York, Pantheon Books, (ISBN 978-0-375-42447-2)
- Laurin 2010, p. 163
- (en) Aurore Canoville et Michel Laurin, « Evolution of humeral microanatomy and lifestyle in amniotes, and some comments on paleobiological inferences », Biological Journal of the Linnean Society, vol. 100, no 2, , p. 384–406 (DOI 10.1111/j.1095-8312.2010.01431.x)
- (en) Michel Laurin, Aurore Canoville et Alexandra Quilhac, « Use of paleontological and molecular data in supertrees for comparative studies: the example of lissamphibian femoral microanatomy », Journal of Anatomy, vol. 215, no 2, , p. 110–123 (PMID 19508493, PMCID 2740958, DOI 10.1111/j.1469-7580.2009.01104.x)
- Clack 2012, p. 87–9
- (en) Michel Laurin, Marc Girondot et Armand de Ricqlès, « Early tetrapod evolution », Trends in Ecology & Evolution, vol. 15, no 3, , p. 118–123 (ISSN 0169-5347, PMID 10675932, DOI 10.1016/S0169-5347(99)01780-2, lire en ligne [archive du ], consulté le )
- Laurin 2010, p. 9
- Benton 2009, p. 99
- « Tétrapodes », sur universalis.fr (consulté le )
- (en) Union internationale pour la conservation de la nature, Liste rouge de l'UICN, 2014.3. Summary Statistics for Globally Threatened Species. Table 1: Numbers of threatened species by major groups of organisms (1996–2014).
- Stephen Jay Gould, Comme les huit doigts de la main. Réflexions sur l'histoire naturelle, Éditions du Seuil, , p. 87.
- W. Hennig, Phylogenetic Systematics, Illinois University Press, 1966, traduit par D. Dwight Davis & R. Zangerl.
- (en) M. Ruta, M.I. Coates et D.L.J. Quicke, « Early tetrapod relationships revisited », Biological Reviews of the Cambridge Philosophical Society, vol. 78, , p. 251–345 (DOI 10.1017/S1464793102006103, lire en ligne [PDF])
- (en) D. Marjanović et M. Laurin, « The origin(s) of modern amphibians: a commentary », Evolutionary Biology, vol. 36, no 3, , p. 336–338 (DOI 10.1007/s11692-009-9065-8, lire en ligne [PDF])
- (en) J.S. Anderson, R.R. Reisz, D. Scott, N.B. Fröbisch et S.S. Sumida, « A stem batrachian from the Early Permian of Texas and the origin of frogs and salamanders », Nature, vol. 453, , p. 515-518 (résumé).
- Jennifer Clack, « Le premier pied à terre », Pour la science, no 340, (résumé)
- «Un poisson à poumon, le dipneuste », documentaire audiovisuel de Alain Devez, production du CNRS, 1991.
- . Une mâchoire fossile de cœlacanthe a été trouvée dans une strate datée de -410 Ma près de Buchan, dans l'état de Victoria, en Australie. C'est le plus vieux fossile connu en 2007, et il a reçu le nom de Eoactinistia foreyi lors de la publication de la découverte en septembre 2006.
- Gaël Clément, « Les tétrapodes de paléo-Belgique », Pour la science, (résumé)
- Les amphibiens possèdent cinq doigts à leurs pattes arrière, quatre à leurs pattes avant. Les amniotes (oiseaux, reptiles, mammifères) présentent le modèle ancestral de cinq doigts (avec parfois des modifications superposées à cet état initial : quatre orteils pour les oiseaux, voire moins chez les mammifères à sabots que forment les ongulés). « L'embryologie montrerait qu'à l'origine cette disposition secondaire est bien acquise à partir d'un membre pentadactyle ». Cf Guillaume Lecointre, Hervé Le Guyader, Classification phylogénétique du vivant, Belin, , p. 9.
- Voir polydactylie chez les tétrapodomorphes (en)
- André Beaumont, Pierre Cassier et Daniel Richard, Biologie animale. Les Cordés : Anatomie comparée des vertébrés, Dunod, , p. 203-204.
- Les nageoires pectorales sont articulées à la ceinture scapulaire par l'humérus, les nageoires pelviennes à la ceinture pelvienne par le fémur.
- communiqué de presse du CNRS du 31 mai 2018 au sujet de l'article Euryhaline ecology of early tetrapods revealed by stable isotopes, par Jean Goedert et al. publié dans Nature du 30 mai 2018
- Euryhaline ecology of early tetrapods revealed by stable isotopes, Nature du 30 mai 2018.
- La sortie des eaux – saumâtres – des premiers tétrapodes, Pour la science, le 10/07/2018.
- « Découverte du plus vieil humérus », Sciences et Avenir.fr, (lire en ligne)
- Cécile Dumas, « Un poisson en marche vers la terre ferme », Sciences et Avenir, (lire en ligne)
- John Noble Wilford, « Scientists Call Fish Fossil the 'Missing Link' », The New York Times, (lire en ligne)
- « Premier pas sur terre », Caroline Idoux, Futura-Sciences, Paris, 4 juillet 2002, citant la revue Nature.
- O.A. Lebedev et M.I. Coates, « The postcranial skeleton of the Devonian tetrapod Tulerpeton curtum Lebedev », Zoological Journal of the Linnean Society, vol. 114, no 3, (DOI 10.1111/j.1096-3642.1995.tb00119.x)
- Joël Ignasse, « Comment les premiers tétrapodes sont sortis de l’eau », Sciences et Avenir.fr, (lire en ligne), citant S.E. Pierce, P.E. Ahlberg, J.R. Hutchinson, J.L. Molna, S. Sanchez, P. Tafforeau et J.A. Clack, « Vertebral architecture in the earliest stem tetrapods », Nature, (DOI 10.1038/nature11825, lire en ligne)
- Du nom du paléontologue américain Alfred Romer, qui le premier remarqua l'absence de fossiles de tétrapodes entre -360 et -340 Ma.
- Caroline Idoux, « Premier pas sur terre », sur Futura-sciences, (consulté le ).
- (en) J. A. Clark, « An early tetrapod from ‘Romer's Gap’ », Nature, vol. 418, , p. 72-76 (DOI 10.1038/nature00824).
- « Fossile du premier marcheur », Futura-Sciences, 7 juillet 2002, citant la revue Nature.
- Laurin, M, Systématique, paléontologie et biologie évolutive moderne : l’exemple de la sortie des eaux des vertébrés, Paris, Ellipses, , 176 p. (ISBN 978-2-7298-3892-8)
- (en) Kenneth V. Kardong, Vertebrates : Comparative Anatomy, Function, Evolution, McGraw-Hill, , p. 667.
- (en) M. A. MacIver et al, « Massive increase in visual range preceded the origin of terrestrial vertebrates », Proceedings of the National Academy of Sciences of the United states of America, vol. 114, no 12, (DOI 10.1073/pnas.1615563114).
- « Westlothiana », encyclopédie MSN Encarta.
- T.R. Smithson, R.L. Carroll, A.L. Panchen et S.M. Andrews, « Westlothiana lizziae from the Viséan of East Kirkton, West Lothian, Scotland, and the amniote stem », Earth and Environmental Science Transactions of the Royal Society of Edinburgh, vol. 84, , p. 383-412 (DOI 10.1017/S0263593300006192)
- Michael Taylor, « AMPHIBIANS that came to stay: How did odd-looking creatures that struggled out of warm equatorial waters to live on land hundreds of millions of years ago end up on a Scottish football ground? », New Scientist, (résumé)
- « Ancient reptile tracks unearthed », BBC News.
Voir aussi
Articles connexes
Bibliographie
![]() : document utilisé comme source pour la rédaction de cet article.
: document utilisé comme source pour la rédaction de cet article.
- John A. Long et Richard Cloutier, « Quand les poissons avaient des doigts », Pour la science, no 513, , p. 48-57 (présentation en ligne)
- Jean-Sébastien Steyer, La Terre avant les Dinosaures, Belin éditeur, (ISBN 978-2-701-142067, OCLC 2701142067).

- Michael Benton, Vertebrate Palaeontology, John Wiley & Sons, , 3e éd. (ISBN 978-1-4051-4449-0, lire en ligne), p. 1
- J.A. Clack, Gaining ground: the origin and evolution of tetrapods, Bloomington, Indiana, USA., Indiana University Press, , 2nd éd. (ISBN 9780253356758, lire en ligne)
- Michel Laurin, How Vertebrates Left the Water, University of California Press, (ISBN 978-0-520-26647-6, lire en ligne)
- George R. Jr. McGhee, When the Invasion of Land Failed: The Legacy of the Devonian Extinctions, Columbia University Press, (ISBN 978-0-231-16057-5, lire en ligne)
- Jennifer A. Clack, « The Fin to Limb Transition: New Data, Interpretations, and Hypotheses from Paleontology and Developmental Biology », Annual Review of Earth and Planetary Sciences, vol. 37, no 1, , p. 163–179 (DOI 10.1146/annurev.earth.36.031207.124146, Bibcode 2009AREPS..37..163C)
- Fins Into Limbs: Evolution, Development, and Transformation, Chicago, University of Chicago Press, (ISBN 978-0-226-31340-5, lire en ligne)
- Long JA, Young GC, Holland T, Senden TJ, Fitzgerald EM, « An exceptional Devonian fish from Australia sheds light on tetrapod origins », Nature, vol. 444, no 7116, , p. 199–202 (PMID 17051154, DOI 10.1038/nature05243, Bibcode 2006Natur.444..199L, S2CID 2412640)
- Michael Benton, Vertebrate Palaeontology, Blackwell Publishing, , 3rd éd.
Liens externes
- Ressources relatives au vivant :
- Notices dans des dictionnaires ou encyclopédies généralistes :
 Portail de la paléontologie
Portail de la paléontologie  Portail origine et évolution du vivant
Portail origine et évolution du vivant  Portail de la zoologie
Portail de la zoologie