Tiarajudens
Tiarajudens est un genre éteint de thérapsides anomodontes ayant vécu au Permien moyen (Guadalupien), dans ce qui est aujourd’hui le Brésil. Son unique espèce, Tiarajudens eccentricus, fut nommée en 2011 par Juan Carlos Cisneros, Fernando Abdala, Bruce S. Rubidge, Paula Camboim Dentzien-Dias, et Ana de Oliveira Bueno. Elle n’est connue que par son holotype UFRGS PV393P, un squelette partiel comprenant un crâne très écrasé, des os des membres, et des gastralia[1]. Tiarajudens est caractérisé par une hétérodontie extrême. Sa denture se compose ainsi d’incisives en forme de feuilles, d’une paire de canines hyper allongées et en forme de lame, ainsi que d’une batterie de dents molariformes sur le palais qui formait une surface de broyage. Les incisives et surtout les dents palatales indiquent un régime alimentaire végétarien basé sur des plantes à haute teneur en fibres. Il a été suggéré que les canines hyper développées de l’animal auraient pu servir à écarter des prédateurs ou à éloigner un congénère. Les analyses cladistiques ont montré que Tiarajudens est étroitement apparenté au genre Anomocephalus d’Afrique du Sud avec lequel il partage la présence d’une batterie dentaire capable d’occlusion. Ces deux genres forment un clade de grands anomodontes basaux aux proportions robustes, les Anomocephaloidea, qui aurait rayonné dans le sud de la Pangée il y a plus de 260 millions d’années[1],[2].

| Règne | Animalia |
|---|---|
| Embranchement | Chordata |
| Sous-embr. | Vertebrata |
| Classe | Synapsida |
| Ordre | Therapsida |
| Sous-ordre | † Anomodontia |
| Clade | † Anomocephaloidea |
Espèces de rang inférieur
- Tiarajudens eccentricus Cisneros (d) et al., 2011
Étymologie
Le nom de genre vient de Tiarajú, la localité type, et du Latin dens, « dent ». L’épithète spécifique eccentricus signifie « hors du centre » utilisé ici dans son sens moderne, signifiant quelque chose qui s'écarte d'une norme ou d'un modèle conventionnel ou établi. Une allusion aux dimensions hors norme des canines de cet animal, comparées à celles des autres anomodontes[1].
Description
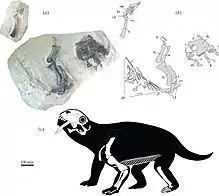
Crâne
Le crâne du Tiarajudens est relativement grand pour un anomodonte basal avec une longueur de plus de 20 cm. Il montre des caractères typiques d’anomodontes comme un museau court et une arcade zygomatique élevée. La hauteur maximale du crâne (10,7 cm) est située près de la marge antérieure de l'orbite. Il est moins haut devant et en arrière de l'orbite, ce qui donne au crâne un profil en dôme. Les orbites, relativement grandes, sont légèrement plus longues que les fosses temporales, un caractère typique des anomodontes basaux chez qui ces deux ouvertures ont le plus souvent des dimensions assez proches (sauf chez Biseridens). Chez les anomodontes évolués, les dicynodontes, les fosses temporales deviennent beaucoup plus grandes que l’orbite. La mâchoire inférieure n’est représentée que par trois os incomplets provenant de la partie postérieure de la mandibule : un morceau de la région dorsale de l’articulaire, la marge dorsale du surangulaire, et un morceau de la lame réfléchie de l’angulaire. Bien que fragmentaire, le surangulaire semble s’élever antéro-dorsalement pour former une éminence coronoïde comme chez Anomocephalus[1],[2].
Dentition
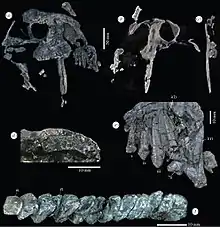
Les principales caractéristiques du Tiarajudens concernent sa dentition fortement hétérodonte. Les prémaxillaires portent au moins quatre incisives en forme de feuilles et dont les couronnes à l’apex pointu portent des dentelures grossières. La première incisive est la plus grande de la série. Les maxillaires portent au moins une incisive (une autre était peut-être présente à l’emplacement d’un morceau d’os manquant sur le fossile), suivie d’une minuscule précanine d’à peine 2 mm de largeur, et immédiatement derrière une énorme canine[2].
La paire de canines hyper développée est la caractéristique la plus frappante de l’animal. Avec leur 12 cm de longueur, les canines du Tiarajudens sont aussi grandes que celles du gorgonopsien Inostrancevia. Mais portées par un crâne de 22,5 cm de longueur, les canines du Tiarajudens sont proportionnellement beaucoup plus grandes : leur taille est égale à plus de 120 % de la hauteur maximale du museau (située juste devant l’orbite) et à plus de 60 % de la longueur totale du crâne. Ces canines, comprimées latéralement, ont une section réniforme à leur base et comporte un émail. Ce qui les distingue clairement de la paire de défenses des anomodontes dicynodontes, lesquelles sont plus courtes et plus épaisses, ont une section circulaire, et sont dépourvues d’émail. Les canines du Tiarajudens sont dépourvues de dentelures et possèdent une racine profonde de 8,5 cm qui atteint presque le toit crânien juste devant l'orbite[1],[2].
Au-dessous de l'orbite, deux os du palais, l'ectoptérygoïde et le ptérygoïde, portent une rangée de treize dents molariformes, élargies transversalement. Les couronnes sont disposées obliquement par rapport à la ligne médiane du crâne, et sont inclinées de 20° par rapport à la marge latérale du jugal. Les dents sont étroitement espacées et disposées en échelons. Les deux dents les plus antérieures sont situées médialement et sont masquées en vue latérale par la très grande canine. Les dents non usées ont des couronnes en forme de lame, avec des crêtes fortes qui se rejoignent apicalement. Une facette naturelle, en croissant, est nettement distincte le long de la marge linguale des nouvelles dents. Les dents usées présentent une surface occlusale élargie, ellipsoïde, avec deux plates-formes d'usure irrégulières : une facette labiale courte et élevée et une facette linguale plus longue et plus basse que la précédente. Toutes les dents palatales sont thécodontes car implantées dans des alvéoles distinctes. Les dents molariformes étant situées sur certains os du palais, et la position très postérieure des grandes canines sur le museau (juste devant les orbites), font que presque toute la dentition marginale est composée de dents incisiformes[1],[2].
Plusieurs dents de substitution sont également visibles, suggérant un taux de remplacement particulièrement élevé, lié à une consommation de plantes abrasives[1],[2].
Squelette post-crânien
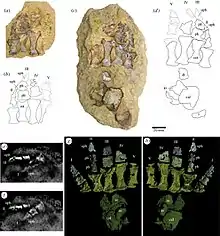
Le squelette post-crânien est représenté par deux fragments de côtes, le membre antérieur gauche presque complet, des gastralia, le tibia gauche, et un pied gauche presque complet. Le membre antérieur est principalement caractérisé par un humérus moyennement robuste, aux extrémités proximales et distales très élargies, la première étant nettement plus large que la seconde. La crête deltopectorale, très développée, forme une plaque presque triangulaire s'étendant sur la moitié de la longueur de l'humérus. Les deux extrémités de l’os sont séparées par une courte diaphyse présentant une face ventrale aplatie et une face dorsale arrondie. De la main, seuls deux doigts ont été retrouvés ainsi que deux os mal conservés et à l’identification incertaine (incluant peut-être un métacarpe). Le plus petit des deux doigts est le mieux conservé. Il comprend deux phalanges non terminales relativement robustes et de formes quadrangulaires, ainsi qu’une phalange terminale représentée par un moulage naturel dans la roche. Cette phalange, dite unguéale car c’est elle qui soutenait une griffe cornée, est plus large et beaucoup plus longue que les phalanges précédentes, et est de forme ogivale avec une extrémité pointue[2].
Au moins quinze gastralia gauches et trois gastralia droites ont été conservées. Plusieurs autres sont préservées par des moulages naturels. Elles se présentent comme des os longs et très minces, mesurant généralement moins d’un millimètre d'épaisseur. Les éléments gauches et droits semblent avoir eu un contact médian, et devaient être courbées du vivant de l’animal[2].
Les éléments du pied ayant été conservés sont l’astragale, le calcanéum, quatre tarses distaux, et cinq doigts partiels, lesquels sont représentés soit par leurs os fossilisés, soit par leurs moulages naturels imprimés dans la roche. Le doigt I, complètement enchâssé dans la gangue rocheuse, n’a été repéré qu’après que l’échantillon fit l’objet d’une tomodensitométrie. Tous les doigts sont robustes. Le doigt I, le plus petit de la série, possède un métatarse réduit et quadrangulaire, et des phalanges qui lui sont comparables en taille. Les métatarses des autres doigts sont plus grands et plus allongées. Le doigt II est caractérisé par des phalanges beaucoup plus petites que le métatarse. Ces phalanges sont également beaucoup plus étroites que celles des doigts III et IV. Les phalanges du doigt V ne sont pas conservées. Une synchondrose était probablement présente entre les os tarsiens et métatarsiens, limitant considérablement la mobilité entre ces éléments. Par contre, la présence d’articulations arthrodiales entre les métatarses et les phalanges proximales, ainsi qu'entre les phalanges, permettait probablement une certaine mobilité dorsoplantaire du pied[2].
Paléobiologie
Régime alimentaire
La dentition du Tiarajudens est clairement celle d’un animal végétarien spécialisé. La morphologie des incisives, lancéolées, et avec des dentelures grossières, est apparue plusieurs fois parmi les tétrapodes herbivores et se retrouve chez d’autres anomodontes basaux (tels Ulemica et Suminia), les paréiasaures, divers groupes de dinosaures (parmi lesquels des sauropodomorphes, des ornithischiens basaux, et les thérizinosaures), certains archosaures pseudosuchiens basaux comme les aétosaures, certains crocodyliformes notosuchiens, et chez les iguanes actuels. Ces incisives coupaient la végétation qui devait ensuite être broyée par la batterie de dents palatales. Bien que le dentaire ne soit pas connu chez Tiarajudens, les larges surfaces d'usure observées sur les dents palatales montrent qu’une occlusion dentaire était certainement possible avec les dents de la mandibule, comme cela a été confirmé chez son cousin Anomocephalus. Chez ce dernier, l'articulation cranio-mandibulaire permettait un certain mouvement avant et arrière de la mâchoire inférieure (appelé propalinie) qui participait à écraser la matière végétale. Selon Cisneros et al., étant donné les fortes ressemblances entre Anomocephalus et Tiarajudens, il est probable que ce dernier bénéficiait du même mécanisme. Le mouvement mandibulaire était toufefois plus limité que le propalinisme avancé reconnu chez Suminia et les dicynodontes, mais comparable à la propalinisation naissante proposée pour Ulemica. La possibilité d’une occlusion dentaire, associée à la fois à une forte usure et un taux élevé de remplacement des dents, suggère que Tiarajudens devait se nourrir de plantes particulièrement fibreuses[1],[2].
Fonction des canines en dents de sabre

Tiarajudens est remarquable par ses canines en dents de sabre, uniques parmi les anomodontes. Avant sa découverte, les plus vieux herbivores à dents de sabre étaient des mammifères datant du Paléocène, il y a 60 millions d’années, comme le pantodonte Titanoides et le uintathère Prodinoceras. Tiarajudens démontre que cette spécialisation était déjà apparue durant le Paléozoïque, 200 millions d’années auparavant. Les canines en dents de sabre du Tiarajudens semblent fragiles au premier abord, mais elles étaient renforcées structurellement par leur section réniforme. La racine extrêmement longue de la canine lui assurait également une stabilité pendant son utilisation. Cisneros et al. suggèrent que les canines du Tiarajudens étaient principalement utilisées pour repousser un rival, que ce soit lors d’une compétition visuelle ou lors d’un affrontement direct, comme le font plusieurs espèces actuelles de cerfs à dents de sabre, tels le cerf d’eau, les porte-musc, et les Muntjacs. Dans le cas d’un combat physique, il est suggéré que Tiarajudens utilisait ses canines pour lacérer la surface corporelle de son adversaire, plutôt que pour infliger des blessures profondes. Cette stratégie aurait permis au Tiarajudens de réduire le risque de briser ses canines, comme ce serait le cas si la canine perforait profondément le corps d'un adversaire se déplaçant brusquement pour s'échapper. Il est probable également que ces canines aient été utilisées pour dissuader des prédateurs[1],[2].

Chez les cerfs à dents de sabre, qu’ils soient actuels ou fossiles, les grandes canines sont uniquement l’apanage des mâles. Il est possible que l’holotype et pour l’instant unique spécimen connu du Tiarajudens représente un individu mâle. Un dimorphisme sexuel chez les anomodontes a été signalé chez les dicynodontes Aulacephalodon et Diictodon, grâce à l’étude de nombreux spécimens. Tiarajudens et Anomocephalus, représenté chacun par un seul spécimen, se ressemblent énormément et ne se différencient que par la présence d’une paire de canines chez le premier, et leur absence chez le second. Cisneros et al. considèrent que bien que cette différence pourrait être l’expression d’un dimorphisme sexuel, il est encore prématuré de voir Tiarajudens et Anomocephalus comme le mâle et la femelle d’un même taxon. Ils préfèrent pour l’instant considérer ces deux genres, trouvés dans différents bassins sédimentaires, comme deux taxons distincts, et que seule la découverte de nouveaux spécimens permettra peut-être de résoudre cette question[2].
Un comportement agonistique basé sur l’affichage visuel et/ou le combat physique avec des dents de sabre constituait une alternative au combat tête contre tête pratiqué par les dinocéphales herbivores contemporains. Ce schéma se retrouve aujourd’hui par exemple entre les cerfs à dents de sabre et les ruminants équipés de bois ou de cornes. Ainsi, certaines spécialisations comportementales considérées comme caractéristiques des mammifères du Cénozoïque, étaient déjà présentes chez les herbivores terrestres du Permien moyen. C’est à cette époque que s’établit pour la première fois un réseau trophique moderne dans lequel une grande et diverse population d'herbivores supporte un nombre relativement faible de carnivores supérieurs[2].
Répartition géographique et stratigraphique
Les fossiles du Tiarajudens furent découvert en 2009 dans le Membre Morro Pelado de la Formation Rio do Rasto, près de la ville de São Gabriel, dans l’État Brésilien du Rio Grande do Sul (District de Tiarajú). Les os de l’animal étaient préservés dans une lentille de grès intercalée dans des siltstones massifs. Ces roches correspondent à des sédiments déposés dans une ancienne plaine inondable, coupée par de petits canaux fluviaux[2].
D’autres sites du Membre Morro Pelado ont livré divers tétrapodes. De la ferme Boqueirão, proviennent le dinocéphale carnivore Pampaphoneus[3], le petit dicynodonte Rastodon[4], et l’amphibien Konzhukovia sangabrielensis[5]. Le site de la ferme Fagundes a fourni le paréiasaure Provelosaurus[6], des dents de dinocéphales carnivores et herbivores (Anteosauridae, Titanosuchidae, et Tapinocephalidae)[7], ainsi qu’un amphibien indéterminé[8]. Des environs de Serra do Cadeado, dans l’État du Paraná, proviennent l’amphibien Australerpeton[8], une espèce indéterminée du dicynodonte Endothiodon[9], ainsi qu’un dinocéphale Tapinocephalidae indéterminé montrant des similitudes avec les genres Moschops et Moschognathus[8].
Le Membre Morro Pelado est considéré comme étant en corrélation avec la Zone d'assemblage à Tapinocephalus du Karoo Sud-Africain en raison de la co-occurrence des dinocéphales et des paréiasaures[1]. Des datations radiométriques ont donné un âge Capitanien inférieur pour cette biozone Sud-Africaine[10],[11]. Des incertitudes demeures sur la contemporanéité des sites Brésiliens. D’après Boos et al., la localisation précise de plusieurs découvertes anciennes est incertaine et ces taxa pourraient provenir de niveaux stratigraphiques différents au sein de la Formation Rio do Rasto, dont l’âge s’étend du Wordien au Wuchiapingien (milieu du Permien moyen au début du Permien supérieur). C’est le cas de la faune de Serra do Cadeado, où les restes de tapinocephalidé et du dicynodonte Endothiodon pourraient provenir de deux niveaux distincts[8]. Toutefois, le genre Sud-Africain Anomocephalus, qui est très similaire au Tiarajudens, provient de la Zone à Tapinocephalus et est donc daté du Capitanien[12],[2]. Un âge comparable est très probable pour le taxon Brésilien.
Phylogénie
L’analyse cladistique présentée par Cisneros et al. réunit Tiarajudens et Anomocephalus dans un nouveau clade nommé Anomocephaloidea, lequel occupe l’une des positions les plus basale parmi les anomodontes[1].
Ci-dessous, le cladogramme montrant la position phylogénétique du Tiarajudens et des Anomocephaloidea [1]:
| Therapsida |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Notes et références
- (en) J.C. Cisneros, F. Abdala, B.S. Rubidge, P.C.D. Dias et A.O. Bueno, « Dental occlusion in a 260-million-year-old therapsid with saber canines from the Permian of Brazil », Science, vol. 331, , p. 1603-1605 (DOI 10.1126/science.1200305)
- (en) J.C. Cisneros, F. Abdala, T. Jashashvili, A. Bueno et P. Dentzien-Dias, « Tiarajudens eccentricus and Anomocephalus africanus, two bizarre anomodonts (Synapsida, Therapsida) with dental occlusion from the Permian of Gondwana », Royal Society Open Science, vol. 2 (7): 150090, (DOI 10.1098/rsos.150090)
- (en) J.C. Cisneros, F. Abdala, S. Atayman-Güven, B.S. Rubidge, A.M.C. Şengör et C.L. Schultz, « Carnivorous dinocephalian from the Middle Permian of Brazil and tetrapod dispersal in Pangaea », Proceedings of the National Academy of Sciences of the United States of America, vol. 109, no 5, , p. 1584-1588 (DOI 10.1073/pnas.1115975109, lire en ligne)
- (en) A.D.S. Boos, C.F. Kammerer, C.L. Schultz, M.B. Soares et A.L.R. Ilha, « A New Dicynodont (Therapsida: Anomodontia) from the Permian of Southern Brazil and Its Implications for Bidentalian Origins », Plos ONE, 11(5), (DOI 10.1371/journal.pone.0155000)
- (en) C.P. Pacheco, E. Eltink, R.T. Müller et S.A. Dias-da-Silva, « A new Permian temnospondyl with Russian affinities from South America, the new family Konzhukoviidae, and the phylogenetic status of Archegosauroidea », Journal of Systematic Palaeontology, (DOI 10.1080/14772019.2016.1164763)
- (en) J.C. Cisneros, F. Abdala et M.C. Malabarba, « Pareiasaurids from the Rio do Rasto Formation, southern Brazil: Biostratigraphic implications for Permian faunas of the Paraná Basin », Revista Brasileira de Paleontologia, vol. 8(1), , p. 13-24
- (en) M.C. Langer, « The first record of dinocephalians in South America: Late Permian (Rio do Rasto Formation) of the Paraná Basin, Brazil », Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, vol. 215, , p. 69-95
- (en) A.D.S. Boos, C.F. Kammerer, C.L. Schultz et V.D. Paes Neto, « A tapinocephalid dinocephalian (Synapsida, Therapsida) from the Rio do Rasto Formation (Paraná Basin, Brazil): Taxonomic, ontogenetic and biostratigraphic considerations », Journal of American Earth Sciences, vol. 63, , p. 375-384
- (en) A.D.S. Boos, C.L. Schultz, C.S. Vega et J.J. Aumond, « On the presence of the Late Permian dicynodont Endothiodon in Brazil », Palaeontology, vol. 56, , p. 837-848 (DOI 10.1111/pala.12020)
- (en) B.S. Rubidge, D. Erwin, J. Ramenazi, S.A. Browning et W.J. De Klerk, « High-precision temporal calibration of Late Permian vertebrate biostratigraphy: U-Pb zircon constrains from the Karoo Supergroup, South Africa », Geology, no 41(3), , p. 363-366 (DOI 10.1130/G33622.1)
- (en) M.O. Day, J. Ramenazi, S.A. Bowring, P.M. Sadler, D.H. Erwin, F. Abdala et B.S. Rubidge, « When and how did the terrestrial mid-Permian mass extinction occur? Evidence from the tetrapod record of the Karoo Basin, South Africa », Proceedings of the Royal Society B, no 282:20150834, (DOI 10.1098/rspb.2015.0834)
- (en) S. Modesto, B. Rubidge et J. Welman, « The most basal anomodont therapsid and the primacy of Gondwana in the evolution of the anomodonts », Proceedings of the Royal Society B 266, , p. 331-337 (DOI 10.1098/rspb.1999.0642)
 Portail de la paléontologie
Portail de la paléontologie  Portail de l’herpétologie
Portail de l’herpétologie  Portail du Brésil
Portail du Brésil