BK channel
| KCNMA1 | |||||||
|---|---|---|---|---|---|---|---|
 The domain structure of BK channels | |||||||
| Identifiers | |||||||
| Symbol | KCNMA1 | ||||||
| Alt. symbols | SLO | ||||||
| NCBI gene | 3778 | ||||||
| HGNC | 6284 | ||||||
| OMIM | 600150 | ||||||
| RefSeq | NM_002247 | ||||||
| UniProt | Q12791 | ||||||
| Other data | |||||||
| Locus | Chr. 10 q22 | ||||||
| |||||||
| KCNMB1 | |||||||
|---|---|---|---|---|---|---|---|
| Identifiers | |||||||
| Symbol | KCNMB1 | ||||||
| NCBI gene | 3779 | ||||||
| HGNC | 6285 | ||||||
| OMIM | 603951 | ||||||
| RefSeq | NM_004137 | ||||||
| UniProt | Q16558 | ||||||
| Other data | |||||||
| Locus | Chr. 5 q34 | ||||||
| |||||||
| KCNMB2 | |||||||
|---|---|---|---|---|---|---|---|
| Identifiers | |||||||
| Symbol | KCNMB2 | ||||||
| NCBI gene | 10242 | ||||||
| HGNC | 6286 | ||||||
| OMIM | 605214 | ||||||
| RefSeq | NM_181361 | ||||||
| UniProt | Q9Y691 | ||||||
| Other data | |||||||
| Locus | Chr. 3 q26.32 | ||||||
| |||||||
| KCNMB3 | |||||||
|---|---|---|---|---|---|---|---|
| Identifiers | |||||||
| Symbol | KCNMB3 | ||||||
| Alt. symbols | KCNMB2, KCNMBL | ||||||
| NCBI gene | 27094 | ||||||
| HGNC | 6287 | ||||||
| OMIM | 605222 | ||||||
| RefSeq | NM_171828 | ||||||
| UniProt | Q9NPA1 | ||||||
| Other data | |||||||
| Locus | Chr. 3 q26.3-q27 | ||||||
| |||||||
| KCNMB3L | |
|---|---|
| Identifiers | |
| Symbol | KCNMB3L |
| Alt. symbols | KCNMB2L, KCNMBLP |
| NCBI gene | 27093 |
| HGNC | 6288 |
| RefSeq | NG_002679 |
| Other data | |
| Locus | Chr. 22 q11.1 |
| KCNMB4 | |||||||
|---|---|---|---|---|---|---|---|
| Identifiers | |||||||
| Symbol | KCNMB4 | ||||||
| NCBI gene | 27345 | ||||||
| HGNC | 6289 | ||||||
| OMIM | 605223 | ||||||
| RefSeq | NM_014505 | ||||||
| UniProt | Q86W47 | ||||||
| Other data | |||||||
| Locus | Chr. 12 q15 | ||||||
| |||||||
| Calcium-activated BK potassium channel alpha subunit | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Identifiers | |||||||||
| Symbol | BK_channel_a | ||||||||
| Pfam | PF03493 | ||||||||
| InterPro | IPR003929 | ||||||||
| |||||||||
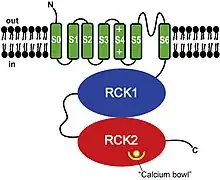
BK channels (big potassium), are large conductance calcium-activated potassium channels,[1] also known as Maxi-K, slo1, or Kca1.1. BK channels are voltage-gated potassium channels that conduct large amounts of potassium ions (K+) across the cell membrane, hence their name, big potassium. These channels can be activated (opened) by either electrical means, or by increasing Ca2+ concentrations in the cell.[2][3] BK channels help regulate physiological processes, such as circadian behavioral rhythms and neuronal excitability.[4] BK channels are also involved in many processes in the body, as it is a ubiquitous channel. They have a tetrameric structure that is composed of a transmembrane domain, voltage sensing domain, potassium channel domain, and a cytoplasmic C-terminal domain, with many X-ray structures for reference. Their function is to repolarize the membrane potential by allowing for potassium to flow outward, in response to a depolarization or increase in calcium levels.
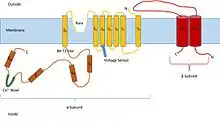
Structure
Structurally, BK channels are homologous to voltage- and ligand-gated potassium channels, having a voltage sensor and pore as the membrane-spanning domain and a cytosolic domain for the binding of intracellular calcium and magnesium.[5] Each monomer of the channel-forming alpha subunit is the product of the KCNMA1 gene (also known as Slo1). The Slo1 subunit has three main structural domains, each with a distinct function: the voltage sensing domain (VSD) senses membrane potential across the membrane, the cytosolic domain (senses calcium concentration, Ca²⁺ ions), and the pore-gate domain (PGD) which opens and closes to regulate potassium permeation. The activation gate resides in the PGD, which is located at either the cytosolic side of S6 or the selectivity filter (selectivity is the preference of a channel to conduct a specific ion).[5] The Voltage sensing domain and pore-gated domain are collectively referred as the membrane-spanning domains and are formed by transmembrane segments S1-S4 and S5-S6, respectively. Within the S4 helix contains a series of positively charged residues which serve as the primary voltage sensor.[6]
BK channels are quite similar to voltage gated K⁺ channels, however, in BK channels only one positively charged residue (Arg213) is involved in voltage sensing across the membrane.[5] Also unique to BK channels is an additional S0 segment, this segment is required for β subunit modulation.[7][8] and voltage sensitivity.[9]
The Cytosolic domain is composed of two RCK (regulator of potassium conductance) domains, RCK1 and RCK2. These domains contain two high affinity Ca²⁺ binding sites: one in the RCK1 domain and the other in a region termed the Ca²⁺ bowl that consists of a series of Aspartic acid (Asp) residues that are located in the RCK2 domain. The Mg²⁺ binding site is located between the VSD and the cytosolic domain, which is formed by: Asp residues within the S0-S1 loop, Asparagine residues in the cytosolic end of S2, and Glutamine residues in RCK1.[5] In forming the Mg²⁺ binding site, two residues come from the RCK1 of one Slo1 subunit and the other two residues come from the VSD of the neighboring subunit. In order for these residues to coordinate the Mg²⁺ ion, the VSD and cytosolic domain from neighboring subunits must be in close proximity.[5] Modulatory beta subunits (encoded by KCNMB1, KCNMB2, KCNMB3, or KCNMB4) can associate with the tetrameric channel. There are four types of β subunits (β1-4), each of which have different expression patterns that modify the gating properties of the BK channel. The β1 subunit is primarily responsible for smooth muscle cell expression, both β2 and β3 subunits are neuronally expressed, while β4 is expressed within the brain.[5] The VSD associates with the PGD via three major interactions:
- Physical connection between the VSD and PGD through the S4-S5 linker.
- Interactions between the S4-S5 linker and the cytosolic side of S6.
- Interactions between S4 and S5 of a neighboring subunit.
Regulation
BK channels are associated and modulated by a wide variety of intra- and extracellular factors, such as auxiliary subunits (β, γ), Slobs (slo binding protein), phosphorylation, membrane voltage, chemical ligands (Ca²⁺, Mg²⁺), PKC, The BK α-subunits assemble 1:1 with four different auxiliary types of β-subunits (β1, β2, β3 or β4).[10]
Trafficking to and expression of BK channels in the plasma membrane has been found to be regulated by distinct splicing motifs located within the intracellular C-terminal RCK domains. In particular a splice variant that excluded these motifs prevented cell surface expression of BK channels and suggests that such a mechanism impacts physiology and pathophysiology.[10]
BK channels in the vascular system are modulated by agents naturally produced in the body, such as angiotensin II (Ang II), high glucose or arachidonic acid (AA) which is modulated in diabetes by oxidative stress (ROS).[10]
A weaker voltage sensitivity allows BK channels to function in a wide range of membrane potentials. This ensures that the channel can properly perform its physiological function.[11]
Inhibition of BK channel activity by phosphorylation of S695 by protein kinase C (PKC) is dependent on the phosphorylation of S1151 in C terminus of channel alpha-subunit. Only one of these phosphorylations in the tetrameric structure needs to occur for inhibition to be successful. Protein phosphatase 1 counteracts phosphorylation of S695. PKC decreases channel opening probability by shortening the channel open time and prolonging the closed state of the channel. PKC does not affect the single-channel conductance, voltage dependence, or the calcium sensitivity of BK channels.[11]
Activation mechanism
BK channels are synergistically activated through the binding of calcium and magnesium ions, but can also be activated via voltage dependence.[10] Ca²⁺ - dependent activation occurs when intracellular Ca²⁺ binds to two high affinity binding sites: one located in the C-terminus of the RCK2 domain (Ca²⁺ bowl), and the other located in the RCK1 domain.[5] The binding site within the RCK1 domain has somewhat of a lower affinity for calcium than the Ca²⁺ bowl, but is responsible for a larger portion of the Ca²⁺ sensitivity.[12] Voltage and calcium activate BK channels using two parallel mechanisms, with the voltage sensors and the Ca²⁺ bindings sites coupling to the activation gate independently, except for a weak interaction between the two mechanisms. The Ca²⁺ bowl accelerates activation kinetics at low Ca²⁺ concentrations while RCK1 site influences both activation and deactivation kinetics.[11] One mechanism model was originally proposed by Monod, Wyman, and Changeux, known as the MWC model. The MWC model for BK channels explains that a conformational change of the activation gate in channel opening is accompanied by a conformational change to the Ca²⁺ binding site, which increases the affinity of Ca²⁺ binding.[12]
Magnesium-dependent activation of BK channels activates via a low-affinity metal binding site that is independent from Ca²⁺-dependent activation. The Mg²⁺ sensor activates BK channels by shifting the activation voltage to a more negative range. Mg²⁺ activates the channel only when the voltage sensor domain stays in the activated state. The cytosolic tail domain (CTD) is a chemical sensor that has multiple binding sites for different ligands. The CTD activates the BK channel when bound with intracellular Mg²⁺ to allow for interaction with the voltage sensor domain (VSD).[11] Magnesium is predominantly coordinated by six oxygen atoms from the side chains of oxygen-containing residues, main chain carbonyl groups in proteins, or water molecules.[12] D99 at the C-terminus of the S0-S1 loop and N172 in the S2-S3 loop contain side chain oxygens in the voltage sensor domain that are essential for Mg²⁺ binding. Much like the Ca²⁺-dependent activation model, Mg²⁺-dependent activation can also be described by an allosteric MCW gating model. While calcium activates the channel largely independent of the voltage sensor, magnesium activates the channel by channel by an electrostatic interaction with the voltage sensor.[12] This is also known as the Nudging model, in which Magnesium activates the channel by pushing the voltage sensor via electrostatic interactions and involves the interactions among side chains in different structural domains.[5] Energy provided by voltage, Ca²⁺, and Mg²⁺ binding will propagate to the activation gate of BK channels to initiate ion conduction through the pore.[5]
Effects on the neuron, organ, body as a whole
Cellular level
BK channels help regulate both the firing of neurons and neurotransmitter release.[13] This modulation of synaptic transmission and electrical discharge at the cellular level is due to BK channel expression in conjunction with other potassium-calcium channels.[10] The opening of these channels causes a drive towards the potassium equilibrium potential and thus play a role in speeding up the repolarization of action potentials.[10] This would effectively allow for more rapid stimulation.[10] There is also a role played in shaping the general repolarization of cells, and thus after hyperpolarization (AHP) of action potentials.[14] The role that BK channels have in the fast phase of AHP has been studied extensively in the hippocampus.[14] It can also play a role in inhibiting the release of neurotransmitters.[15] There are many BK channels in Purkinje cells in the cerebellum, thus highlighting their role in motor coordination and function.[14] Furthermore, BK channels play a role in modulating the activity of dendrites as well as astrocytes and microglia.[15] They not only play a role in the CNS (central nervous system) but also in smooth muscle contractions, the secretion of endocrine cells, and the proliferation of cells.[13] Various γ subunits during early brain development are involved in neuronal excitability and in non-excitable cells they often are responsible as a driving force of calcium.[10] Therefore, these subunits can be targets for therapeutic treatments as BK channel activators.[10] There is further evidence that inhibiting BK channels would prevent the efflux of potassium and thus reduce the usage of ATP, in effect allowing for neuronal survival in low oxygen environments.[10] BK channels can also function as a neuronal protectant in terms such as limiting calcium entry into the cells through methionine oxidation.[10]
Organ level
BK channels also play a role in hearing.[14] This was found when the BK ɑ-subunit was knocked out in mice and progressive loss of cochlear hair cells, and thus hearing loss, was observed.[14] BK channels are not only involved in hearing, but also circadian rhythms. Slo binding proteins (Slobs) can modulate BK channels as a function of circadian rhythms in neurons.[10] BK channels are expressed in the suprachiasmatic nucleus (SCN), which is characterized to influence the pathophysiology of sleep.[14] BK channel openers can also have a protective effect on the cardiovascular system.[10] At a low concentration of calcium BK channels have a greater impact on vascular tone.[10] Furthermore, the signaling system of BK channels in the cardiovascular system have an influence on the functioning of coronary blood flow.[10] One of the functions of the β subunit in the brain includes inhibition of the BK channels, allowing for the slowing of channel properties as well as the ability to aid in prevention of seizures in the temporal lobe.[10]
Bodily function level
Mutations of BK channels, resulting in a lower amount of expression in mRNA, is more common in people who are mentally challenged (via hypofunction [15]), schizophrenic or autistic.[10] Moreover, increased repolarization caused by BK channel mutations may lead to dependency of alcohol initiation of dyskinesias, epilepsy or paroxysmal movement disorders.[10] Not only are BK channels important in many cellular processes in the adult it also is crucial for proper nutrition supply to a developing fetus.[10] Thus, estrogen can cause an increase in the density of BK channels in the uterus.[10] However, increased expression of BK channels have been found in tumor cells, and this could influence future cancer therapy, discussed more in the pharmacology section.[10] BK channels are ubiquitous throughout the body and thus have a large and vast impact on the body as a whole and at a more cellular level, as discussed.
Pharmacology
Potential issues
Several issues arise when there is a deficit in BK channels. Consequences of the malfunctioning BK channel can affect the functioning of a person in many ways, some more life threatening than others. BK channels can be activated by exogenous pollutants and endogenous gasotransmitters carbon monoxide,[16][17] nitric oxide, and hydrogen sulphide.[18] Mutations in the proteins involved with BK channels or genes encoding BK channels are involved in many diseases. A malfunction of BK channels can proliferate in many disorders such as: epilepsy, cancer, diabetes, asthma, and hypertension.[13] Specifically, β1 defect can increase blood pressure and hydrosaline retention in the kidney.[13] Both loss of function and gain of function mutations have been found to be involved in disorders such as epilepsy and chronic pain.[15] Furthermore, increases in BK channel activation, through gain-of-function mutants and amplification, has links to epilepsy and cancer.[13] Moreover, BK channels play a role in tumors as well as cancers. In certain cancers gBK, a variant ion channel called glioma BK channel, can be found.[14] It is known that BK channels do in some way influence the division of cells during replication, which when unregulated can lead to cancers and tumors.[14] Moreover, an aspect studied includes the migration of cancer cells and the role in which BK channels can facilitate this migration, though much is still unknown.[14] Another reason why BK channel understanding is important involves its role in organ transplant surgery. This is due to the activation of BK channels influencing repolarization of the resting membrane potential.[10] Thus, understanding is crucial for safety in effective transplantation.
Current developments
BK channels can be used as pharmacological targets for the treatment of several medical disorders including stroke[19] and overactive bladder.[20] There have been attempts to develop synthetic molecules targeting BK channels,[21] however their efforts have proven largely ineffective thus far. For instance, BMS-204352, a molecule developed by Bristol-Myers Squibb, failed to improve clinical outcome in stroke patients compared to placebo.[22] However, there have been some success from the agonist to BKCa channels, BMS-204352, in treating deficits observed in Fmr1 knockout mice, a model of Fragile X syndrome.[23][24] BK channels also function as a blocker in ischemia and are a focus in investigating its use as a therapy for stroke.[10]
Future directions
There are many applications for therapeutic strategies involving BK channels. There has been research displaying that a blockage of BK channels results in an increase in neurotransmitter release, effectively indicating future therapeutic possibilities in cognition enhancement, improved memory, and relieving depression.[13] A behavioral response to alcohol is also modulated by BK channels,[10] therefore further understanding of this relationship can aid treatment in patients who are alcoholics. Oxidative stress on BK channels can lead to the negative impairments of lowering blood pressure through cardiovascular relaxation have on both aging and disease.[10] Thus, the signaling system can be involved in treating hypertension and atherosclerosis[10] through targeting of the ɑ subunit to prevent these detrimental effects. Furthermore, the known role that BK channels can play in cancer and tumors is limited. Thus, there is not a lot of current knowledge regarding specific aspects of BK channels that can influence tumors and cancers.[14] Further study is crucial, as this could lead to immense development in treatments for those suffering from cancer and tumors. It is known that epilepsies are due to over-excitability of neurons, which BK channels have a large impact on controlling hyperexcitability.[4] Therefore, understanding could influence the treatment of epilepsy. Overall, BK channels are a target for future pharmacological agents that can be used for benevolent treatments of disease.
See also
- Calcium-activated potassium channel subunit alpha-1
- Calcium-activated potassium channel
- Voltage-gated potassium channel
References
- ↑ Zang K, Zhang Y, Hu J, Wang Y (2018). "The Large Conductance Calcium- and Voltage-activated Potassium Channel (BK) and Epilepsy". CNS & Neurological Disorders Drug Targets. 17 (4): 248–254. doi:10.2174/1871527317666180404104055. PMID 29623857.
- ↑ Miller, C. (2000). Genome Biology, 1(4), reviews0004.1. https://dx.doi.org/10.1186/gb-2000-1-4-reviews0004
- ↑ Yuan, P., Leonetti, M., Pico, A., Hsiung, Y., & MacKinnon, R. (2010). Structure of the Human BK Channel Ca2+-Activation Apparatus at 3.0 A Resolution. Science, 329(5988), 182-186. https://dx.doi.org/10.1126/science.1190414
- 1 2 N'Gouemo P (November 2011). "Targeting BK (big potassium) channels in epilepsy". Expert Opinion on Therapeutic Targets. 15 (11): 1283–95. doi:10.1517/14728222.2011.620607. PMC 3219529. PMID 21923633.
- 1 2 3 4 5 6 7 8 9 Lee US, Cui J (September 2010). "BK channel activation: structural and functional insights". Trends in Neurosciences. 33 (9): 415–23. doi:10.1016/j.tins.2010.06.004. PMC 2929326. PMID 20663573.
- ↑ Atkinson NS, Robertson GA, Ganetzky B (August 1991). "A component of calcium-activated potassium channels encoded by the Drosophila slo locus". Science. 253 (5019): 551–5. Bibcode:1991Sci...253..551A. doi:10.1126/science.1857984. PMID 1857984. S2CID 11317087.
- ↑ Morrow JP, Zakharov SI, Liu G, Yang L, Sok AJ, Marx SO (March 2006). "Defining the BK channel domains required for beta1-subunit modulation". Proceedings of the National Academy of Sciences of the United States of America. 103 (13): 5096–101. Bibcode:2006PNAS..103.5096M. doi:10.1073/pnas.0600907103. PMC 1458800. PMID 16549765.
- ↑ Wallner M, Meera P, Toro L (December 1996). "Determinant for beta-subunit regulation in high-conductance voltage-activated and Ca(2+)-sensitive K+ channels: an additional transmembrane region at the N terminus". Proceedings of the National Academy of Sciences of the United States of America. 93 (25): 14922–7. Bibcode:1996PNAS...9314922W. doi:10.1073/pnas.93.25.14922. PMC 26238. PMID 8962157.
- ↑ Koval OM, Fan Y, Rothberg BS (March 2007). "A role for the S0 transmembrane segment in voltage-dependent gating of BK channels". The Journal of General Physiology. 129 (3): 209–20. doi:10.1085/jgp.200609662. PMC 2151615. PMID 17296928.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 Hermann A, Sitdikova GF, Weiger TM (August 2015). "Oxidative Stress and Maxi Calcium-Activated Potassium (BK) Channels". Biomolecules. 5 (3): 1870–911. doi:10.3390/biom5031870. PMC 4598779. PMID 26287261.
- 1 2 3 4 Yang H, Zhang G, Cui J (2015). "BK channels: multiple sensors, one activation gate". Frontiers in Physiology. 6: 29. doi:10.3389/fphys.2015.00029. PMC 4319557. PMID 25705194.
- 1 2 3 4 Cui J, Yang H, Lee US (March 2009). "Molecular mechanisms of BK channel activation". Cellular and Molecular Life Sciences. 66 (5): 852–75. doi:10.1007/s00018-008-8609-x. PMC 2694844. PMID 19099186.
- 1 2 3 4 5 6 Yu M, Liu SL, Sun PB, Pan H, Tian CL, Zhang LH (January 2016). "Peptide toxins and small-molecule blockers of BK channels". Acta Pharmacologica Sinica. 37 (1): 56–66. doi:10.1038/aps.2015.139. PMC 4722972. PMID 26725735.
- 1 2 3 4 5 6 7 8 9 10 Bentzen BH, Olesen SP, Rønn LC, Grunnet M (2014). "BK channel activators and their therapeutic perspectives". Frontiers in Physiology. 5: 389. doi:10.3389/fphys.2014.00389. PMC 4191079. PMID 25346695.
- 1 2 3 4 Contet C, Goulding SP, Kuljis DA, Barth AL (2016). "BK Channels in the Central Nervous System". Big on Bk - Current Insights into the Function of Large Conductance Voltage- and Ca2+ -Activated K+ Channels at the Molecular, Cellular and Systemic Levels. International Review of Neurobiology. Vol. 128. pp. 281–342. doi:10.1016/bs.irn.2016.04.001. ISBN 9780128036198. PMC 4902275. PMID 27238267.
- ↑ Dubuis E, Potier M, Wang R, Vandier C (Feb 2005). "Continuous inhalation of carbon monoxide attenuates hypoxic pulmonary hypertension development presumably through activation of BKCa channels". Cardiovascular Research. 65 (3): 751–61. doi:10.1016/j.cardiores.2004.11.007. PMID 15664403.
- ↑ Hou S, Xu R, Heinemann SH, Hoshi T (Mar 2008). "The RCK1 high-affinity Ca2+ sensor confers carbon monoxide sensitivity to Slo1 BK channels". Proceedings of the National Academy of Sciences of the United States of America. 105 (10): 4039–43. Bibcode:2008PNAS..105.4039H. doi:10.1073/pnas.0800304105. PMC 2268785. PMID 18316727.
- ↑ Sitdikova GF, Weiger TM, Hermann A (Feb 2010). "Hydrogen sulfide increases calcium-activated potassium (BK) channel activity of rat pituitary tumor cells". Pflügers Archiv. 459 (3): 389–97. doi:10.1007/s00424-009-0737-0. PMID 19802723. S2CID 23073556.
- ↑ Gribkoff VK, Starrett JE, Dworetzky SI (Apr 2001). "Maxi-K potassium channels: form, function, and modulation of a class of endogenous regulators of intracellular calcium". The Neuroscientist. 7 (2): 166–77. doi:10.1177/107385840100700211. PMID 11496927. S2CID 8791803.
- ↑ Layne JJ, Nausch B, Olesen SP, Nelson MT (Feb 2010). "BK channel activation by NS11021 decreases excitability and contractility of urinary bladder smooth muscle". American Journal of Physiology. Regulatory, Integrative and Comparative Physiology. 298 (2): R378–84. doi:10.1152/ajpregu.00458.2009. PMC 2828174. PMID 19923353.
- ↑ Gribkoff VK, Winquist RJ (May 2005). "Voltage-gated cation channel modulators for the treatment of stroke". Expert Opinion on Investigational Drugs. 14 (5): 579–92. doi:10.1517/13543784.14.5.579. PMID 15926865. S2CID 10236998.
- ↑ Jensen BS (2002). "BMS-204352: a potassium channel opener developed for the treatment of stroke". CNS Drug Reviews. 8 (4): 353–60. doi:10.1111/j.1527-3458.2002.tb00233.x. PMC 6741660. PMID 12481191.
- ↑ Laumonnier F, Roger S, Guérin P, Molinari F, M'rad R, Cahard D, Belhadj A, Halayem M, Persico AM, Elia M, Romano V, Holbert S, Andres C, Chaabouni H, Colleaux L, Constant J, Le Guennec JY, Briault S (2006). "Association of a functional deficit of the BKCa channel, a synaptic regulator of neuronal excitability, with autism and mental retardation". The American Journal of Psychiatry. 163 (9): 1622–1629. doi:10.1176/ajp.2006.163.9.1622. PMID 16946189. S2CID 25225269.
- ↑ Hébert B; Pietropaolo S; Même S; Laudier B; Laugeray A; Doisne N; Quartier A; Lefeuvre S; Got L; Cahard D; Laumonnier F; Crusio WE; Pichon J; Menuet A; Perche O; Briault S (2014). "Rescue of fragile X syndrome phenotypes in Fmr1 KO mice by a BKCa channel opener molecule". Orphanet Journal of Rare Diseases. 9: 124. doi:10.1186/s13023-014-0124-6. PMC 4237919. PMID 25079250.
Further reading
- Ge L, Hoa NT, Wilson Z, Arismendi-Morillo G, Kong XT, Tajhya RB, Beeton C, Jadus MR (October 2014). "Big Potassium (BK) ion channels in biology, disease and possible targets for cancer immunotherapy". International Immunopharmacology. 22 (2): 427–43. doi:10.1016/j.intimp.2014.06.040. PMC 5472047. PMID 25027630.
- Kyle BD, Braun AP (2014). "The regulation of BK channel activity by pre- and post-translational modifications". Frontiers in Physiology. 5: 316. doi:10.3389/fphys.2014.00316. PMC 4141542. PMID 25202279.
- Nardi A, Olesen SP (2008). "BK channel modulators: a comprehensive overview". Current Medicinal Chemistry. 15 (11): 1126–46. doi:10.2174/092986708784221412. PMID 18473808.
- Zhang J, Yan J (2014). "Regulation of BK channels by auxiliary γ subunits". Frontiers in Physiology. 5: 401. doi:10.3389/fphys.2014.00401. PMC 4197896. PMID 25360119.
External links
- BK+Channels at the US National Library of Medicine Medical Subject Headings (MeSH)
- "Calcium-Activated Potassium Channels". IUPHAR Database of Receptors and Ion Channels. International Union of Basic and Clinical Pharmacology.