Delta endotoxin
| delta endotoxin, N-terminal domain | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 | |||||||||
| Identifiers | |||||||||
| Symbol | Endotoxin_N | ||||||||
| Pfam | PF03945 | ||||||||
| InterPro | IPR005639 | ||||||||
| SCOP2 | 1dlc / SCOPe / SUPFAM | ||||||||
| TCDB | 1.C.2 | ||||||||
| |||||||||
| delta endotoxin, middle domain | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Identifiers | |||||||||
| Symbol | Endotoxin_M | ||||||||
| Pfam | PF00555 | ||||||||
| Pfam clan | CL0568 | ||||||||
| InterPro | IPR015790 | ||||||||
| SCOP2 | 1dlc / SCOPe / SUPFAM | ||||||||
| TCDB | 1.C.2 | ||||||||
| |||||||||
| delta endotoxin, middle domain, Cry2A and Cry18 | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 insecticidal crystal protein cry2aa | |||||||||
| Identifiers | |||||||||
| Symbol | Endotoxin_mid | ||||||||
| Pfam | PF09131 | ||||||||
| InterPro | IPR015214 | ||||||||
| SCOP2 | 1i5p / SCOPe / SUPFAM | ||||||||
| |||||||||
| delta endotoxin, C-terminal | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Identifiers | |||||||||
| Symbol | Endotoxin_C | ||||||||
| Pfam | PF03944 | ||||||||
| Pfam clan | CL0202 | ||||||||
| InterPro | IPR005638 | ||||||||
| SCOP2 | 1dlc / SCOPe / SUPFAM | ||||||||
| TCDB | 1.C.2 | ||||||||
| CDD | cd04085 | ||||||||
| |||||||||
| Cytolytic Delta-endotoxin Cyt1/2 | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Identifiers | |||||||||
| Symbol | CytB | ||||||||
| Pfam | PF01338 | ||||||||
| InterPro | IPR001615 | ||||||||
| SCOP2 | 1cby / SCOPe / SUPFAM | ||||||||
| TCDB | 1.C.71 | ||||||||
| |||||||||
Delta endotoxins (δ-endotoxins) are pore-forming toxins produced by Bacillus thuringiensis species of bacteria. They are useful for their insecticidal action and are the primary toxin produced by Bt maize/corn. During spore formation the bacteria produce crystals of such proteins (hence the name Cry toxins) that are also known as parasporal bodies, next to the endospores; as a result some members are known as a parasporin. The Cyt (cytolytic) toxin group is a group of delta-endotoxins different from the Cry group.
Mechanism of action
When an insect ingests these proteins, they are activated by proteolytic cleavage. The N-terminus is cleaved in all of the proteins and a C-terminal extension is cleaved in some members. Once activated, the endotoxin binds to the gut epithelium and causes cell lysis by the formation of cation-selective channels, which leads to death.[2][1]
For many years there was no clarity as to the relationship between aminopeptidase N and Bt toxins. Although AP-N does bind Cry proteins in vitro[3] (reviewed by Soberón et al. 2009[4] and Pigott & Ellar 2007[5]),[6] no cases of resistance – or even reduced in vitro binding – due to AP-N structure alteration were known through 2002, and there was some doubt that the resistance mechanism was so straight forward. Indeed, Luo et al. 1997, Mohammed et al. 1996, and Zhu et al. 2000 positively found this to not occur in Lepidoptera examples.[3] Subsequently, however Herrero et al. 2005 showed correlation between nonexpression and Bt resistance,[6] and actual resistance was found in Helicoverpa armigera by Zhang et al. 2009,[6][7] in Ostrinia nubilalis by Khajuria et al. 2011, and in Trichoplusia ni by Baxter et al. 2011 and Tiewsiri & Wang 2011 (also all Lepidoptera).[6] There continues to be confirmation that AP-Ns do not by themselves affect resistance in some cases, possibly due to sequential binding by the toxin being required to produce its effect. In this sequence each binding step is theoretically not indispensable, but if it occurs does contribute to the final pore formation result.[7]
Structure
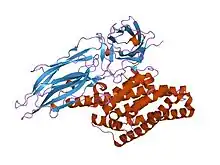
The activated region of the delta toxin is composed of three distinct structural domains: an N-terminal helical bundle domain (InterPro: IPR005639) involved in membrane insertion and pore formation; a beta-sheet central domain involved in receptor binding; and a C-terminal beta-sandwich domain (InterPro: IPR005638) that interacts with the N-terminal domain to form a channel.[1][2]
Types
B. thuringiensis encodes many proteins of the delta endotoxin family (InterPro: IPR038979), with some strains encoding multiple types simultaneously.[8] A gene mostly found on plasmids,[9] delta-entotoxins sometimes show up in genomes of other species, albeit at a lower proportion than those found in B. thuringiensis.[10] The gene names looks like Cry3Bb, which in this case indicates a Cry toxin of superfamily 3 family B subfamily b.[11]
Cry proteins that are interesting to cancer research are listed under a parasporin (PS) nomenclature in addition to the Cry nomenclature. They do not kill insects, but instead kill leukemia cells.[12][13][14] The Cyt toxins tend to form their own group distinct from Cry toxins.[15] Not all Cry — crystal-form — toxins directly share a common root.[16] Examples of non-three-domain toxins that nevertheless have a Cry name include Cry34/35Ab1 and related beta-sandwich binary (Bin-like) toxins, Cry6Aa, and many beta-sandwich parasporins.[17]
Specific delta-endotoxins that have been inserted with genetic engineering include Cry3Bb1 found in MON 863 and Cry1Ab found in MON 810, both of which are maize/corn cultivars. Cry3Bb1 is particularly useful because it kills Coleopteran insects such as the corn rootworm, an activity not seen in other Cry proteins.[1] Other common toxins include Cry2Ab and Cry1F in cotton and maize/corn.[18] In addition, Cry1Ac is effective as a vaccine adjuvant in humans.[19]
Some insects populations have started to develop resistance towards delta endotoxin, with five resistant species found as of 2013. Plants with two kinds of delta endotoxins tend to make resistance happen slower, as the insects have to evolve to overcome both toxins at once. Planting non-Bt plants with the resistant plants will reduce the selection pressure for developing the toxin. Finally, two-toxin plants should not be planted with one-toxin plants, as one-toxin plants act as a stepping stone for adaption in this case.[18]
References
- 1 2 3 4 Galitsky N, Cody V, Wojtczak A, Ghosh D, Luft JR, Pangborn W, English L (August 2001). "Structure of the insecticidal bacterial delta-endotoxin Cry3Bb1 of Bacillus thuringiensis". Acta Crystallographica. Section D, Biological Crystallography. 57 (Pt 8): 1101–1109. doi:10.1107/S0907444901008186. PMID 11468393.
- 1 2 Grochulski P, Masson L, Borisova S, Pusztai-Carey M, Schwartz JL, Brousseau R, Cygler M (December 1995). "Bacillus thuringiensis CryIA(a) insecticidal toxin: crystal structure and channel formation". Journal of Molecular Biology. 254 (3): 447–464. doi:10.1006/jmbi.1995.0630. PMID 7490762.
- 1 2 Ferré J, Van Rie J (2002). "Biochemistry and genetics of insect resistance to Bacillus thuringiensis". Annual Review of Entomology. Annual Reviews. 47 (1): 501–533. doi:10.1146/annurev.ento.47.091201.145234. PMID 11729083.
- ↑ Soberón M, Gill SS, Bravo A (April 2009). "Signaling versus punching hole: How do Bacillus thuringiensis toxins kill insect midgut cells?". Cellular and Molecular Life Sciences. Springer. 66 (8): 1337–1349. doi:10.1007/s00018-008-8330-9. PMID 19132293. S2CID 5928827.
- ↑ Pigott CR, Ellar DJ (June 2007). "Role of receptors in Bacillus thuringiensis crystal toxin activity". Microbiology and Molecular Biology Reviews. American Society for Microbiology. 71 (2): 255–281. doi:10.1128/mmbr.00034-06. PMC 1899880. PMID 17554045. S2CID 13982571.
- 1 2 3 4 Pardo-López L, Soberón M, Bravo A (January 2013). "Bacillus thuringiensis insecticidal three-domain Cry toxins: mode of action, insect resistance and consequences for crop protection". FEMS Microbiology Reviews. Federation of European Microbiological Societies (OUP). 37 (1): 3–22. doi:10.1111/j.1574-6976.2012.00341.x. PMID 22540421.
- 1 2 Vachon V, Laprade R, Schwartz JL (September 2012). "Current models of the mode of action of Bacillus thuringiensis insecticidal crystal proteins: a critical review". Journal of Invertebrate Pathology. Academic Press (Elsevier). 111 (1): 1–12. doi:10.1016/j.jip.2012.05.001. PMID 22617276.
- ↑ "Pesticidal crystal protein (IPR038979)". InterPro. Retrieved 12 April 2019.
- ↑ Dean DH (1984). "Biochemical genetics of the bacterial insect-control agent Bacillus thuringiensis: basic principles and prospects for genetic engineering". Biotechnology & Genetic Engineering Reviews. 2: 341–363. doi:10.1080/02648725.1984.10647804. PMID 6443645.
- ↑ "Species: Pesticidal crystal protein (IPR038979)". InterPro.
- ↑ "Bacillus thuringiensis Toxin Nomenclature". Bt toxin specificity database. Retrieved 12 April 2019.
- ↑ Mizuki E, Park YS, Saitoh H, Yamashita S, Akao T, Higuchi K, Ohba M (July 2000). "Parasporin, a human leukemic cell-recognizing parasporal protein of Bacillus thuringiensis". Clinical and Diagnostic Laboratory Immunology. 7 (4): 625–634. doi:10.1128/CDLI.7.4.625-634.2000. PMC 95925. PMID 10882663.
- ↑ Ohba M, Mizuki E, Uemori A (January 2009). "Parasporin, a new anticancer protein group from Bacillus thuringiensis". Anticancer Research. 29 (1): 427–433. PMID 19331182.
- ↑ "List of Parasporins". Committee of Parasporin Classification and Nomenclature. Accessed Jan 4, 2013
- ↑ Crickmore N. "Other Cry Seqences" (PDF). Retrieved 12 April 2019.
- ↑ Crickmore N, Zeigler DR, Feitelson J, Schnepf E, Van Rie J, Lereclus D, et al. (September 1998). "Revision of the nomenclature for the Bacillus thuringiensis pesticidal crystal proteins". Microbiology and Molecular Biology Reviews. 62 (3): 807–813. doi:10.1128/MMBR.62.3.807-813.1998. PMC 98935. PMID 9729610.
- ↑ Kelker MS, Berry C, Evans SL, Pai R, McCaskill DG, Wang NX, et al. (2014-11-12). "Structural and biophysical characterization of Bacillus thuringiensis insecticidal proteins Cry34Ab1 and Cry35Ab1". PLOS ONE. 9 (11): e112555. Bibcode:2014PLoSO...9k2555K. doi:10.1371/journal.pone.0112555. PMC 4229197. PMID 25390338.
- 1 2 Tabashnik BE, Brévault T, Carrière Y (June 2013). "Insect resistance to Bt crops: lessons from the first billion acres". Nature Biotechnology. 31 (6): 510–521. doi:10.1038/nbt.2597. PMID 23752438. S2CID 205278530.
- ↑ Rodriguez-Monroy MA, Moreno-Fierros L (March 2010). "Striking activation of NALT and nasal passages lymphocytes induced by intranasal immunization with Cry1Ac protoxin". Scandinavian Journal of Immunology. 71 (3): 159–168. doi:10.1111/j.1365-3083.2009.02358.x. PMID 20415781.
Further reading
- Bravo A, Gill SS, Soberón M (March 2007). "Mode of action of Bacillus thuringiensis Cry and Cyt toxins and their potential for insect control". Toxicon. 49 (4): 423–435. doi:10.1016/j.toxicon.2006.11.022. PMC 1857359. PMID 17198720.
- Pigott CR, Ellar DJ (June 2007). "Role of receptors in Bacillus thuringiensis crystal toxin activity". Microbiology and Molecular Biology Reviews. 71 (2): 255–281. doi:10.1128/MMBR.00034-06. PMC 1899880. PMID 17554045.
- Palma L, Muñoz D, Berry C, Murillo J, Caballero P (December 2014). "Bacillus thuringiensis toxins: an overview of their biocidal activity". Toxins. 6 (12): 3296–3325. doi:10.3390/toxins6123296. PMC 4280536. PMID 25514092.
External links
- Cry3Bb1 at the United States Environmental Protection Agency
| |||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Bacterial toxins |
| ||||||||||||||||||||||||||
| Mycotoxins |
| ||||||||||||||||||||||||||
| Plant toxins |
| ||||||||||||||||||||||||||
| Invertebrate toxins |
| ||||||||||||||||||||||||||
| Vertebrate toxins |
| ||||||||||||||||||||||||||
| |||||||||||||||||||||||||||
| |||||||||||||||||||||||||||