History of catecholamine research
The catecholamines comprise the endogenous substances dopamine, noradrenaline (norepinephrine) and adrenaline (epinephrine) as well as numerous artificially synthesized compounds such as isoprenaline. Their investigation constitutes a prominent chapter in the history of physiology, biochemistry and pharmacology. Adrenaline was the first hormone extracted from its endocrine gland and obtained in pure form, before the word hormone was coined.[1] It was also the first hormone the structure and biosynthesis of which were clarified. Apart from acetylcholine, adrenaline and noradrenaline were the first neurotransmitters to be discovered and the first intercellular biochemical signals to be found in intracellular vesicles. The β-adrenoceptor was the first G protein-coupled receptor the gene of which was cloned. Goal-directed catecholamine research began with the preparation by George Oliver and Edward Albert Sharpey-Schafer of a pharmacologically active extract from the adrenal glands.
Adrenaline in the adrenal medulla
Forerunners
In a book on asthma of the 19th century, first published in 1860,[2] the British physician and physiologist Henry Hyde Salter (1823–1871) included a chapter on treatment ″by stimulants″. Strong coffee was very helpful, presumably because it dispelled sleep, which favoured asthma. Even more impressive, however, was the response to ″strong mental emotion″: ″The cure of asthma by violent emotion is more sudden and complete than by any other remedy whatever; indeed, I know few things more striking and curious in the whole history of therapeutics. ... The cure ... takes no time; it is instantaneous, the intensest paroxysm ceases on the instant.″ ″Cure″ due to release of adrenaline from the adrenals is the retrospective interpretation.
At the same time that Salter unwittingly made use of the adrenal medulla, the French physician Alfred Vulpian found that there was something unique about it:[3] material scraped from it coloured green when ferric chloride was added. This did neither occur with the adrenal cortex nor with any other tissue. The adrenal medulla hence contained "une matière spéciale, inconnue jusqu’ici et qui constitue le signe particulier de ces organes" ("a special material, unknown hereforth and which constitutes the particular sign of these organs"). Vulpian even came to the insight that the substance entered "le torrent circulatoire" ("the circulatory torrent"), for blood from the adrenal veins did give the ferric chloride reaction.

In the early 1890s, the German pharmacologist Carl Jacobj (1857–1944) in the laboratory of Oswald Schmiedeberg in Strasbourg studied the relationship between the adrenals and the intestine. Electrical stimulation of the vagus nerve or injection of muscarine elicited peristalsis. This peristalis was promptly abolished by electrical stimulation of the adrenals.[4] The experiment has been called "the first indirect demonstration of the role of the adrenal medulla as an endocrine organ <and> actually a more sophisticated demonstration of the adrenal medullary function than the classic study of Oliver and Schäfer".[5] While this may be true, Jacobj did not envisage a chemical signal secreted into the blood to influence distant organs, in other words a hormone, but nerves running from the adrenals to the gut, "Hemmungsbahnen für die Darmbewegung".
Oliver and Schäfer 1893/94
George Oliver was a physician practicing in the spa town of Harrogate in North Yorkshire. Edward Albert Schäfer was Professor of Physiology at University College London. In 1918, he prefixed the surname of his physiology teacher William Sharpey to his own to become Edward Albert Sharpey Schafer. The canonical story, told by Henry Hallett Dale, who worked at University College London from 1902 to 1904, runs as follows:[6]
Dr Oliver, I was told, … had a liking and a ′flair′ for the invention of simple appliances, with which observations and experiments could be made on the human subject. Dr Oliver had invented a small instrument with which he claimed to be able to measure, through the unbroken skin, the diameter of a living artery, such as the radial artery at the wrist. He appears to have used his family in his experiments, and a young son was the subject of a series, in which Dr Oliver measured the diameter of the radial artery, and observed the effect upon it of injecting extracts of various animal glands under the skin. … We may picture, then, Professor Schafer, in the old physiological laboratory at University College, … finishing an experiment of some kind, in which he was recording the arterial blood pressure of an anaesthetised dog. … To him enters Dr Oliver, with the story of the experiments on his boy, and, in particular, with the statement that injection under the skin of a glycerin extract from calf’s suprarenal gland was followed by a definite narrowing of the radial artery. Professor Schafer is said to have been entirely sceptical, and to have attributed the observation to self-delusion. … He can hardly be blamed, I think; knowing even what we now know about the action of this extract, which of us would be prepared to believe that injecting it under a boy’s skin would cause his radial artery to become measurably more slender? Dr Oliver, however, is persistent; he … suggests that, at least, it will do no harm to inject into the circulation, through a vein, a little of the suprarenal extract, which he produces from his pocket. So Professor Schafer makes the injection, expecting a triumphant demonstration of nothing, and finds himself standing ′like some watcher of the skies, when a new planet swims into his ken,′ watching the mercury rise in the manometer with amazing rapidity and to an astounding height.
However often this tale has been reiterated, it is not beyond doubt. Dale himself said that it was handed down in University College, and showed some surprise that the constriction of the radial artery was measurable. Of Oliver's descendants, none recalled experiments on his son.[7] Dale's report of subcutaneous injections contradicts the concerned parties. Oliver:[8] “During the winter of 1893–4, while prosecuting an inquiry as to … agents that vary the calibre of … arteries … I found that the administration by the mouth of a glycerine extract of the adrenals of the sheep and calf produced a marked constrictive action on the arteries.” Schäfer:[9] “In the autumn of 1893 there called upon me in my laboratory in University College a gentleman who was personally unknown to me. … I found that my visitor was Dr. George Oliver, <who> was desirous of discussing with me the results which he had been obtaining from the exhibition by the mouth of extracts from certain animal tissues, and the effects which these had in his hands produced upon the blood vessels of man.” Systemic effects of orally given adrenaline are highly unlikely. So details of the canonical text may be legend.

On March 10, 1894, Oliver and Schäfer presented their findings to the Physiological Society in London.[10] A 47-pages account followed a year later, in the style of the time without statistics, but with precise description of many individual experiments and 25 recordings on kymograph smoked drums, showing, besides the blood pressure increase, reflex bradycardia and contraction of the spleen.[11] ″It appears to be established as the result of these investigations that ... the suprarenal capsules are to be regarded although ductless, as strictly secreting glands. The material which they form and which is found, at least in its fully active condition, only in the medulla of the gland, produces striking physiological effects upon the muscular tissue generally, and especially upon that of the heart and arteries. Its action ... is produced mainly if not entirely by direct action.″
The reports created a sensation. Oliver was not slow to try adrenal extracts in patients, orally again and rather indiscriminately, from Addison's disease, hypotension (″loss of vasomotor tone″), Diabetes mellitus and Diabetes insipidus to Graves' disease (″exophthalmic goitre″).[8] It seems he adhered to contemporary ideas of organotherapy, believing that powerful substances existed in tissues and ought to be discovered for medicinal use.[12] In fact he immediately went on to extract the pituitary gland and, again with Schäfer, discovered vasopressin.[13] In 1903 adrenaline, meanwhile purified, was first used in asthma. The use was based, not on the bronchodilator effect, which was discovered later,[14][15] but on the vasoconstrictor effect, which was hoped to alleviate the “turgidity of the bronchial mucosa” – presumably vascular congestion and edema.[16] Also as of 1903, adrenaline was added to local anesthetic solutions. The surgeon Heinrich Braun in Leipzig showed that it prolonged the anesthesia at the injection site and simultaneously reduced ″systemic″ effects elsewhere in the body.[17]
Independent discoverers
A year after Oliver and Schäfer, Władysław Szymonowicz (1869–1939) and Napoleon Cybulski of the Jagiellonian University in Kraków reported essentially similar findings and conclusions. In one aspect, they went beyond the work in England: blood from the adrenal veins caused hypertension when injected intravenously in a recipient dog, whereas blood from other veins did not, showing that the adrenal pressor substance was in fact secreted into the blood and confirming Vulpian.[18] The Polish authors freely acknowledged the priority of Oliver and Schäfer, and the British authors acknowledged the independence of Szymonowicz and Cybulski. The main difference was in the location of the action: to the periphery by Oliver and Schäfer but, erroneously, to the central nervous system by Szymonowicz and Cybulski.
Another year later, the US-American ophthalmologist William Bates, perhaps motivated like Oliver, instilled adrenal extracts into the eye, found that ″the conjunctiva of the globe and lids whitened in a few minutes″, correctly explained the effect by vasoconstriction, and administered the extracts in various eye diseases.[19] Later catecholamine literature has not done him justice. Even an ophthalmological review article of 1905 mentions him only in passing.[20]
Chemistry
In 1897 John Jacob Abel in Baltimore partially purified adrenal extracts to what he called “epinephrin”,[21] and Otto von Fürth in Strasbourg to what he called “Suprarenin”.[22] The Japanese chemist Jōkichi Takamine, who had set up his own laboratory in New York, invented new isolation procedure and obtained it in pure crystal form in 1901[23] and arranged for Parke-Davis to market it as ”Adrenalin”, spelt without the terminal “e”. In 1903, natural adrenaline was found to be optically active and levorotary, in 1905 synthesis of the racemate was achieved by Friedrich Stolz at Hoechst AG in Höchst (Frankfurt am Main) and by Henry Drysdale Dakin at the University of Leeds, in 1906 the chemical structure was elucidated by Ernst Joseph Friedmann (1877–1956) in Strasbourg,[24] and in 1908 the dextrorotary enantiomer was shown to be almost inactive by Arthur Robertson Cushney (1866–1926) at the University of Michigan, leading him to brilliantly conclude that ″the ‘receptive substance’ affected by adrenalin″ is able to discriminate between the optical isomers and, hence, itself optically active.[25] Overall, 32 designations have been coined, of which “adrenaline”, preferred in the United Kingdom, and “epinephrine”, preferred in the United States, persist as generic names in the scientific literature.[26]
Adrenaline as a transmitter
A new chapter was opened when Max Lewandowsky in 1899 in Berlin observed that adrenal extracts acted on the smooth muscle of the eye and orbit of cats – as the iris dilator muscle and nictitating membrane – in the same way as sympathetic nerve stimulation.[27] The correspondence was extended by John Newport Langley and, under his supervision, Thomas Renton Elliott in Cambridge. In four papers in volume 31, 1904, of the Journal of Physiology Elliott described the similarities organ by organ. His visionary hypothesis stands in the abstract of a presentation to the Physiological Society of May 21, 1904, a little over ten years after Oliver and Schäfer's presentation:[28] ″Adrenalin does not excite sympathetic ganglia when applied to them directly, as does nicotine. Its effective action is localised at the periphery. ... I find that even after ... complete denervation, whether of three days’ or ten months’ duration, the plain muscle of the dilatator pupillae will respond to adrenalin, and that with greater rapidity and longer persistence than does the iris whose nervous relations are uninjured. Therefore, it cannot be than adrenalin excites any structure derived from, and dependent for its persistence on, the peripheral neurone. ... The point at which the stimulus of the chemical excitant is received, and transformed into what may cause the change of tension of the muscle fibre, is perhaps a mechanism developed out of the muscle cell in response to its union with the synapsing sympathetic fibre, the function of which is to receive and transform the nervous impulse. Adrenalin might then be the chemical stimulant liberated on each occasion when the impulse arrives at the periphery.″ The abstract is the ″birth certificate″ of chemical neurotransmission.[29] Elliott was never so explicit again. It seems he was discouraged by the lack of a favourable response from his seniors, Langley in particular, and a few years later he left physiological research.
- The ″birth certificate″ of chemical neurotransmission


The breakthrough for chemical neurotransmission came when, in 1921, Otto Loewi in Graz demonstrated the ″humorale Übertragbarkeit der Herznervenwirkung″ in amphibians.[30] Vagusstoff transmitted inhibition from the vagus nerves, and Acceleransstoff transmitted stimulation from the sympathetic nerves to the heart.[31] Loewi took some years to commit himself with respect to the nature of the Stoffe, but in 1926 he was sure that Vagusstoff was acetylcholine, and in 1936 he wrote:[32] ″I no longer hesitate to identify the Sympathicusstoff with adrenaline.″
He had luck in the latter statement. In most amphibian organs including the heart, the concentration of adrenaline far exceeds that of noradrenaline, and adrenaline is indeed the main transmitter. In mammals, however, difficulties arose. In a comprehensive structure-activity study of adrenaline-like compounds, Dale and the chemist George Barger in 1910 pointed out that Elliott's hypothesis assumed a stricter parallelism between the effects of sympathetic nerve impulses and adrenaline than actually existed.[33] For example, sympathetic impulses shared with adrenaline contractile effects in the trigone but not relaxant effects in the fundus of the cat's urinary bladder. In this respect, ″amino-ethanol-catechol″ – noradrenaline – mimicked sympathetic nerves more closely than adrenaline did. The Harvard Medical School physiologist Walter Bradford Cannon, who had popularized the idea of a sympatho-adrenal system preparing the body for fight and flight, and his colleague Arturo Rosenblueth developed an elaborate but ″queer″[34] theory of two sympathins, sympathin E (excitatory) and sympathin I (inhibitory). The Belgian pharmacologist Zénon Bacq as well as Canadian and US-American pharmacologists between 1934 and 1938 suggested that noradrenaline might be the – or at least one – postganglionic sympathetic transmitter.[34][35] However, nothing definite was brought to light till after the war. In the meantime, Dale created a terminology that since has imprinted the thinking of neuroscientists: that nerve cells should be named after their transmitter, i.e. cholinergic if the transmitter was ″a substance like acetylcholine", and adrenergic if it was ″some substance like adrenaline″.[36]
In 1936, the year of Loewi's final acceptance of adrenaline as the (amphibian) sympathetic transmitter, Dale and Loewi received the Nobel Prize in Physiology or Medicine ″for their discoveries relating to chemical transmission of nerve impulses″.
Formation and destruction
″Our modern knowledge of the biosynthetic pathway for the catecholamines begins in 1939, with the publication of a paper by Peter Holtz and his colleagues: they described the presence in the guinea-pig kidneys of an enzyme that they called dopa decarboxylase, because it catalysed the formation of dopamine and carbon dioxide from the amino acid L-dopa.″ The German-British biochemist Hermann Blaschko (1900–1993), who in 1933 had left Germany because he was Jewish, wrote this in 1987 in Oxford, looking back on “a half-century of research on catecholamine biosynthesis”.[37] The paper by Peter Holtz (1902–1970) and his coworkers originated from the Institute of Pharmacology in Rostock.[38] Already in that same year 1939 both Blaschko,[39] then in Cambridge, and Holtz[40] in Rostock predicted the entire sequence tyrosine → l-DOPA → oxytyramine = dopamine → noradrenaline → adrenaline. Edith Bülbring, who also had fled National Socialist racism in 1933, demonstrated methylation of noradrenaline to adrenaline in adrenal tissue in Oxford in 1949,[41] and Julius Axelrod detected phenylethanolamine N-methyltransferase in Bethesda, Maryland in 1962.[42] The two remaining enzymes, tyrosine hydroxylase and dopamine β-hydroxylase, were also characterized around 1960.
Even before contributing to the formation pathway, Blaschko had discovered a destruction mechanism. An enzyme tyramine oxidase described in 1928[43] also oxidized dopamine, noradrenaline and adrenaline.[44] It was later named monoamine oxidase. This seemed to clarify the fate of the catecholamines in the body, but in 1956 Blaschko suggested that, because the oxidation was slow, “other mechanisms of inactivation … will be found to play an important part. Here is a gap in our knowledge which remains to be filled.”[35] Within a year, Axelrod narrowed the gap by showing that dopamine, noradrenaline and adrenaline were O-methylated by catechol-O-methyl transferase.[45] To fill the gap completely, however, the role of membranes had to be appreciated (see below).
Noradrenaline
Thanks to Holtz and Blaschko it was clear that animals synthesized noradrenaline. What was lacking to attribute a transmitter role to it was proof of its presence in tissues at effective concentrations and not only as a short-lived intermediate. On April 16, 1945, Ulf von Euler of Karolinska Institutet in Stockholm, who had already discovered or co-discovered substance P and prostaglandins, submitted to Nature the first of a series of papers that gave this proof.[46] After many bioassays and chemical assays on organ extracts he concluded[47] that mammalian sympathetically innervated tissues as well as, in small amounts, the brain, but not the nerve-free placenta, contained noradrenaline and that noradrenaline was the sympathin of Cannon and Rosenblueth, the ″physiological transmitter of adrenergic nerve action in mammals″. Overflow of noradrenaline into the venous blood of the cat's spleen upon sympathetic nerve stimulation two years later bore out the conclusion.[48] In amphibian hearts, on the other hand, the transmitter role of adrenaline was confirmed.[47]
The war prevented Peter Holtz and his group in Rostock from being recognized side by side with von Euler as discoverers of the second catecholamine transmitter noradrenaline. Their approach was different. They sought for catecholamines in human urine and found a blood pressure-increasing material Urosympathin that they identified as a mixture of dopamine, noradrenaline and adrenaline.[49] “As to the origin of Urosympathin we would like to suggest the following. Dopamine in urine is the fraction that was not consumed for the synthesis of sympathin E and I. … Sympathin E and I, i.e. noradrenaline and adrenaline, are liberated in the region of the sympathetic nerve endings when these are excited.” The manuscript was received by Springer-Verlag in Leipzig on October 8, 1944. On October 15, the printing office in Braunschweig was destroyed by an airstrike. Publication was delayed to volume 204, 1947, of Naunyn-Schmiedebergs Archiv für Pharmakologie und Experimentelle Pathologie.[49] Peter Holtz later used to cite the paper as ″Holtz et al. 1944/47″ or ″Holtz, Credner and Kroneberg 1944/47″.
Remembering his and Barger's structure-activity analysis of 1910,[33] Dale wrote in 1953:[50] “Doubtless I ought to have seen that nor-adrenaline might be the main transmitter – that Elliott’s theory might be right in principle and faulty only in this detail. … It is easy, of course, to be wise in the light of facts recently discovered; lacking them I failed to jump to the truth, and I can hardly claim credit for having crawled so near and then stopped short of it.”
The next step led to the central nervous system. It was taken by Marthe Vogt, a refugee from Germany who at that time worked with John Henry Gaddum in the Institute of Pharmacology of the University of Edinburgh. ″The presence of noradrenaline and adrenaline in the brain has been demonstrated by von Euler (1946) and Holtz (1950). These substances were supposed, undoubtedly correctly, to occur in the cerebral vasomotor <= vasoconstrictor> nerves. The present work is concerned with the question whether these sympathomimetic amines, besides their role as transmitters at vasomotor endings, play a part in the function of the central nervous tissue itself. In this paper, these amines will be referred to as sympathin, since they were found invariably to occur together, with noradrenaline representing the major component, as is characteristic for the transmitter of the peripheral sympathetic system.″[51] Vogt created a detailed map of noradrenaline in the dog brain. Its uneven distribution, not reflecting the distribution of vasomotor nerves, and its persistence after removal of the superior cervical ganglia made it ″tempting to assign to the cerebral sympathin a transmitter role like that which we assign to the sympathin found in the sympathetic ganglia and their postganglionic fibres.″ Her assignment was confirmed, the finishing touch being the visualization of the noradrenaline as well as adrenaline and (see below) dopamine pathways in the central nervous system by Annica Dahlström and Kjell Fuxe with the formaldehyde fluorescence method developed by Nils-Åke Hillarp (1916–1965) and Bengt Falck (born 1927) in Sweden and by immunochemistry techniques.[52]
Dopamine
As noradrenaline is an intermediate on the path to adrenaline, dopamine is on the path to noradrenaline (and hence adrenaline.) In 1957 dopamine was identified in the human brain by researcher Katharine Montagu. In 1958/59 Arvid Carlsson and his group in the Pharmacology Department of the University of Lund, including the medical students Åke Bertler and Evald Rosengren, not only found dopamine in the brain, but also – like noradrenaline in Marthe Vogt's exemplary study – in uneven distribution, quite different from the distribution of noradrenaline. This argued for a function beyond an intermediate. The concentration was highest in the corpus striatum, which contained only traces of noradrenaline. Carlsson's group had previously found that reserpine, which was known to cause a Parkinsonism syndrome, depleted dopamine (as well as noradrenaline and serotonin) from the brain. They concluded that ″dopamine is concerned with the function of the corpus striatum and thus with the control of motor function″.[53][54] Thus for the first time the reserpine-induced Parkinsonism in laboratory animals and, by implication, Parkinson's disease in humans was related to depletion of striatal dopamine. A year later Oleh Hornykiewicz, who had been introduced to dopamine by Blaschko and was carrying out a colour reaction on extracts of human corpus striatum in the Pharmacological Institute of the University of Vienna, saw the brain dopamine deficiency in Parkinson's disease ″with <his> own naked eye: Instead of the pink colour given by the comparatively high concentrations of dopamine in the control samples, the reaction vials containing the extracts of the Parkinson's disease striatum showed hardly a tinge of pink discoloration″.[55][56]
In 1970, von Euler and Axelrod were two of three winners of the Nobel Prize in Physiology or Medicine, “for their discoveries concerning the humoral transmitters in the nerve terminals and the mechanism for their storage, release and inactivation”, and in 2000 Carlsson was one of three winners who got the prize “for their discoveries concerning signal transduction in the nervous system”.
Membrane passage
Membranes play a twofold role for catecholamines: catecholamines must pass through membranes and deliver their chemical message at membrane receptors.
Catecholamines are synthesized inside cells and sequestered in intracellular vesicles. This was first shown by Blaschko and Arnold Welch (1908–2003) in Oxford[57] and by Hillarp and his group in Lund[58] for the adrenal medulla and later for sympathetic nerves[59] and the brain.[60] In addition the vesicles contained adenosine triphosphate (ATP), with a molar noradrenaline:ATP ratio in sympathetic nerve vesicles of 5.2:1 as determined by Hans-Joachim Schümann (1919–1998) and Horst Grobecker (born 1934) in Peter Holtz′ group at the Goethe University Frankfurt.[61] Blaschko and Welch wondered how the catecholamines got out when nervous impulses reached the cells.[57] Exocytosis was not among the possibilities they considered. It required the analogy of the ″quantal″ release of acetylcholine at the neuromuscular junction shown by Bernard Katz, third winner of the 1970 Nobel Prize in Physiology or Medicine; the demonstration of the co-release with catecholamines of other vesicle constituents such as ATP and dopamine β-hydroxylase; and the unquestionable electron microscopic images of vesicles fusing with the cell membrane[62] – to establish exocytosis.
Acetylcholine, once released, is degraded in the extracellular space by acetylcholinesterase, which faces that space. In the case the catecholamines, however, the enzymes of degradation monoamine oxidase and catechol-O-methyl transferase, like the enzymes of synthesis, are intracellular. Not metabolism but uptake through cell membranes therefore is the primary means of their clearance from the extracellular space. The mechanisms were deciphered beginning in 1959. Axelrod's group in Bethesda wished to clarify the in vivo fate of catecholamines using radioactively labelled catecholamines of high specific activity, which had just become available. 3H-adrenaline and 3H-noradrenaline given intravenously to cats were partly O-methylated, but another part was taken up in the tissues and stored unchanged.[63][64] Erich Muscholl (born 1926) in Mainz, who had worked with Marthe Vogt in Edinburgh, wished to know how cocaine sensitized tissues to catecholamines – a fundamental mechanism of action of cocaine discovered by Otto Loewi and Alfred Fröhlich in 1910 in Vienna.[65] Intravenous noradrenaline was taken up into the heart and spleen of rats, and cocaine prevented the uptake, ″thus increasing the amount of noradrenaline available for combination with the adrenergic receptors″.[66] The uptake of 3H-noradrenaline was severely impaired after sympathectomy, indicating that it occurred mainly into sympathetic nerve terminals. In support of this, Axelrod and Georg Hertting (born 1925) showed that freshly incorporated 3H-noradrenaline was re-released from the cat spleen when the sympathetic nerves were stimulated.[67] A few years later, Leslie Iversen (born 1937) in Cambridge found that other cells also took up catecholamines. He called uptake into noradrenergic neurones, which were cocaine-sensitive, uptake1 and uptake into other cells, which were cocaine-resistant, uptake2. With the reserpine-sensitive uptake from the cytoplasm into the storage vesicles there were thus three catecholamine membrane passage mechanisms. Iversen's book of 1967 “The Uptake and Storage of Noradrenaline in Sympathetic Nerves” was successful,[68] showing the fascination of the field and its rich pharmacology.
With the advent of molecular genetics, the three transport mechanisms have been traced to the proteins and their genes since 1990. They now comprise the plasma membrane noradrenaline transporter (NAT or NET), the classical uptake1, and the analogous dopamine transporter (DAT); the plasma membrane extraneuronal monoamine transporter or organic cation transporter 3 (EMT or SLC22A3), Iversen's uptake2; and the vesicular monoamine transporter (VMAT) with two isoforms. Transporters and intracellular enzymes such as monoamine oxidase operating in series constitute what the pharmacologist Ullrich Trendelenburg at the University of Würzburg called metabolizing systems.[69]
Receptors
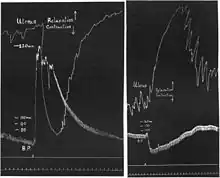

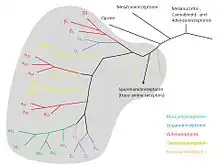
Research on the catecholamines was interwoven with research on their receptors. In 1904, Dale became head of the Wellcome Physiological Research Laboratory in London and started research on ergot extracts. The relevance of his communication in 1906[71] ″On some physiological actions of ergot″ lies less in the effects of the extracts given alone than in their interaction with adrenaline: they reversed the normal pressor effect of adrenaline to a depressor effect and the normal contraction effect on the early-pregnant cat's uterus to relaxation: adrenaline reversal. The pressor and uterine contraction effects of pituitary extracts, in contrast, remained unchanged, as did the effects of adrenaline on the heart and effects of parasympathetic nerve stimulation. Dale clearly saw the specificity of the ″paralytic″ (antagonist) effect of ergot for ″the so-called myoneural junctions connected with the true sympathetic or thoracic-lumbar division of the autonomic nervous system″ – the adrenoceptors. He also saw its specificity for the ″myoneural junctions″ mediating smooth muscle contraction as opposed to those mediating smooth muscle relaxation. But there he stopped. He did not conceive any close relationship between the smooth muscle-inhibitory and the cardiac sites of action of catecholamines.
Catecholamine receptors persisted in this wavering state for more than forty years. Additional blocking agents were found such as tolazoline in Switzerland[72] and phenoxybenzamine in the United States,[73] but like the ergot alkaloids they blocked only the smooth muscle excitatory receptors. Additional agonists also were synthesized. Outstanding among them became isoprenaline, N-isopropyl-noradrenaline, of Boehringer Ingelheim, studied pharmacologically along with adrenaline and other N-substituted noradrenaline derivatives by Richard Rössler (1897–1945) and Heribert Konzett (1912–2004) in Vienna. The Viennese pharmacologists used their own Konzett-Rössler test to examine bronchodilation.[74] Intravenous injection of pilocarpine to induce bronchospasm was followed by intravenous injection of the agonists. “Arrangement of all amines according to their bronchodilator effect yields a series from the most potent, isopropyl-adrenaline, via the approximately equipotent bodies adrenaline, propyl-adrenaline and butyl-adrenaline, to the weakly active isobutyl-adrenaline.”[75] Isoprenaline also exerted marked positive chronotropic and inotropic effects. Boehringer introduced it for use in asthma in 1940. After the war it became available to Germany's former enemies and over the years was traded under about 50 names. In addition to this therapeutic success it was one of the agonists with which Raymond P. Ahlquist solved the ″myoneural junction″ riddle. “By virtue of this property the reputation of the substance spread all over the world and it became a tool for many investigations on different aspects of pharmacology and therapeutics.”[76] The story had a dark side: overdosage caused numerous deaths due to cardiac side effects, an estaimated three thousands in the United Kingdom alone.[77]
Ahlquist was head of the Department of Pharmacology of the University of Georgia School of Medicine, now Georgia Regents University. In 1948 he saw what had escaped Dale in 1906. “The adrenotropic receptors have been considered to be of two classes, those whose action results in excitation and those whose action results in inhibition of the effector cells. Experiments described in this paper indicate that although there are two kinds of adrenotropic receptors they cannot be classified simply as excitatory or inhibitory since each kind of receptor may have either action depending on where it is found.”[78] Ahlquist chose six agonists, including adrenaline, noradrenaline, α-methylnoradrenaline and isoprenaline, and examined their effects on several organs. He found that the six substances possessed two – and only two – rank orders of potency in these organs. For example, the rank order of potency was ″adrenaline > noradrenaline > α-methylnoradrenaline > isoprenaline″ in promoting contraction of blood vessels, but ″isoprenaline > adrenaline > α-methylnoradrenaline > noradrenaline″ in stimulating the heart. The receptor with the first rank order (for example for blood vessel contraction) he called alpha adrenotropic receptor (now α-adrenoceptor or α-adrenergic receptor), while the receptor with the second rank order (for instance for stimulation of the heart, but also for bronchodilation) he called beta adrenotropic receptor (now β-adrenoceptor or β-adrenergic receptor). ″This concept of two fundamental types of receptors is directly opposed to the concept of two mediator substances (sympathin E and sympathin I) as propounded by Cannon and Rosenblueth and now widely quoted as ‘law’ of physiology. ... There is only one adrenergic neuro-hormone, or sympathin, and that sympathin is identical with epinephrine.”[78]
The haze surrounding the receptors was thus blown away. Yet, perhaps because Ahlquist dismissed Cannon and Rosenblueth rather harshly, his manuscript was rejected by the Journal of Pharmacology and Experimental Therapeutics and only in a second submission accepted by the American Journal of Physiology.
In retrospect, although Ahlquist was right in his ″one transmitter – two receptors″ postulate, he erred in the identification of the transmitter with adrenaline. There is an additional qualification. For many responses to sympathetic nerve stimulation, the ATP co-stored with noradrenaline (see above) is a cotransmitter. It acts through purinoceptors.[79] Lastly, Ahlquist failed to adduce the selectivity of all antagonists known at his time for the α-adrenoceptor as an additional argument.
The α,β-terminology initially was slow to spread. This changed with two publications in 1958. In the first, from Lilly Research Laboratories, dichloroisoprenaline selectively blocked some smooth muscle inhibitory effects of adrenaline and isoprenaline;[80] in the second, it blocked cardiac excitatory effects of adrenaline and isoprenaline as well.[81] In the first, which does not mention Ahlquist, dichloroisoprenaline blocked ″certain adrenergic inhibitory receptor sites″; but in the second the results ″support the postulate of Ahlquist (1948) that the adrenotropic inhibitory receptors and the cardiac chronotropic and inotropic adrenergic receptors are functionally identical, i.e., that both are beta type receptors. … It is suggested that this terminology be extended to the realm of adrenergic blocking drugs, e.g., that blocking drugs be designated according to the receptor for which they have the greatest affinity, as either alpha or beta adrenergic blocking drugs.”
Dichloroisoprenaline was the first beta blocker; it retains some intrinsic activity. Pronethalol followed in 1962 and propranolol in 1964,[82] both invented by James Black and his colleagues at Imperial Chemical Industries Pharmaceuticals in England. In 1967, β-adrenoceptors were subdivided into β1 and β2,[83] and a third β type began to be suspected in the late 1970s, above all in adipocytes.[84]
After premonitions for example in the work of the Portuguese pharmacologist Serafim Guimarães, α-adrenoceptor subclassification came in 1971 with the discovery of the self-regulation of noradrenaline release through α-adrenoceptors on noradrenergic synaptic terminals, presynaptic α-autoreceptors. Their existence was initially combated but is now established, for example by the demonstration of their messenger RNA in noradrenergic neurones.[85][86][87] They differed from α-receptors on effector cells and in 1974 became the prototype α2-receptors, the long-known smooth muscle contraction-mediating receptors becoming α1.[88]
Even before dopamine was identified as the third catecholamine transmitter, Blaschko suspected it might possess receptors of its own, since Peter Holtz and his group in 1942 had found that small doses of dopamine lowered the blood pressure of rabbits and guinea pigs, whereas adrenaline always increased the blood pressure.[89] Holtz erred in his interpretation, but Blaschko had ″no doubt that his observations are of the greatest historical importance, as the first indication of an action of dopamine that characteristically and specifically differs from those of the two other catecholamines″.[37] A re-investigation of the blood pressure-lowering effect in dogs in 1964 proposed ″specific dopamine receptors for dilation″,[90] and at the same time evidence for dopamine receptors distinct from α- and β-adrenoceptors accrued from other experimental approaches.
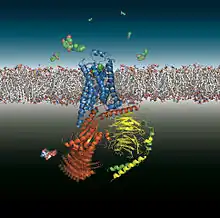
In 1986, the first gene coding for a catecholamine receptor, the β2-adrenoceptor from hamster lung, was cloned by a group of sixteen scientists, among them Robert Lefkowitz and Brian Kobilka of Duke University in Durham, North Carolina.[91] Genes for all mammalian catecholamine receptors have now been cloned, for the nine adrenoceptors α1A, α1B, α1D, α2A, α2B, α2C, β1, β2 and β3 and the five dopamine receptors D1, D2, D3, D4 und D5. Their fine structure, without agonist or agonist-activated, is being studied at high resolution.[92]
Earl Wilbur Sutherland won the 1971 Nobel Prize in Physiology or Medicine ″for his discoveries concerning the mechanisms of the action of hormones″, in particular the discovery of cyclic adenosine monophosphate as second messenger in the action of catecholamines at β-adrenoceptors and of glucagon at glucagon receptors, which led on to the discovery of heterotrimeric G proteins. In 1988 James Black was one of three winners of the Nobel Prize in Physiology or Medicine ″for their discoveries of important principles for drug treatment″, Black's ″important principles″ being the blockade of β-adrenoceptors and of histamine H2 receptors. In 2012, Robert Lefkowitz and Brian Kobilka shared the Nobel Prize in Chemistry ″for studies of G-protein-coupled receptors″.
References
- ↑ John Henderson (2005). "Ernest Starling and 'Hormones': an historical commentary". Journal of Endocrinology. 184 (1): 5–10. doi:10.1677/joe.1.06000. PMID 15642778.
- ↑ Henry Hyde Salter: On Asthma: its pathology and therapy. Philadelphia, Blanchard 1864.
- ↑ A. Vulpian (1856). "Note sur quelques réactions propres à la substance des capsules surrénales". Comptes Rendus de l'Académie des Sciences. 43: 663–665.
- ↑ C. Jacobj (1892). "Beiträge zur physiologischen und pharmakologischen Kenntniss der Darmbewegungen mit besonderer Berücksichtigung der Beziehung der Nebenniere zu denselben". Archiv für experimentelle Pathologie und Pharmakologie. 29 (3–4): 171–211. doi:10.1007/BF01966116. S2CID 21462860.
- ↑ Stephen W. Carmichael; Rochester (1989). "The history of the adrenal medulla". Reviews in the Neurosciences. 2 (2): 83–99. doi:10.1515/REVNEURO.1989.2.2.83. PMID 21561250. S2CID 43591356.
- ↑ H. Dale (1938). "Natural chemical stimulators". Edinburgh Medical Journal. 45 (7): 461–480. PMC 5307706. PMID 29646414.
- ↑ H. Barcroft; J. F. Talbot (1968). "Oliver and Schäfer's discovery of the cardiovascular action of suprarenal extract". Postgraduate Medical Journal. 44 (507): 6–8. doi:10.1136/pgmj.44.507.6. PMC 2466464. PMID 4867248.
- 1 2 George Oliver (1895). "On the therapeutic employment of the suprarenal glands". British Medical Journal. 1895, part 2 (1811): 653–655. doi:10.1136/bmj.2.1811.635. S2CID 220141848.
- ↑ E. A. Schäfer (1908). "On the present condition of our knowledge of the function of the suprarenal capsules". British Medical Journal. 1908, part 1 (2474): 1277–1281. doi:10.1136/bmj.1.2474.1277. PMC 2436722. PMID 20763861.
- ↑ G. Oliver; E. A. Schäfer (1894). "On the physiological action of extract of the suprarenal capsules". The Journal of Physiology. 16 (3–4): I–IV. doi:10.1113/jphysiol.1894.sp000503. PMC 1514529. PMID 16992168.
- ↑ G. Oliver; E. A. Schäfer (1894). "The physiological effects of extracts of the suprarenal capsules". The Journal of Physiology. 18 (3): 230–276. doi:10.1113/jphysiol.1895.sp000564. PMC 1514629. PMID 16992252.
- ↑ Merriley Borell (1976). "Organotherapy, British physiology, and discovery of the internal secretions". Journal of the History of Biology. 9 (2): 235–286. doi:10.1007/bf00209884. PMID 11610067. S2CID 12016920.
- ↑ G. Oliver; E. A. Schäfer (1895). "On the physiological action of extracts of pituitary body and certain other glandular organs". The Journal of Physiology. 18 (3): 277–279. doi:10.1113/jphysiol.1895.sp000565. PMC 1514634. PMID 16992253.
- ↑ R. H. Kahn (1907). "Zur Physiologie der Trachea". Archiv für Anatomie und Physiologie, Archiv für Physiologie: 398–426.
- ↑ Hans Januschke; Leo Pollak (1911). "Zur Pharmakologie der Bronchialmuskulatur". Archiv für experimentelle Pathologie und Pharmakologie. 66 (3): 205–220. doi:10.1007/BF01841068. S2CID 12164315.
- ↑ Jesse G. M. Bullowa; David M. Kaplan (1903). "On the hypodermatic use of adrenalin chloride in the treatment of asthmatic attacks". New York Medical Journal and Medical Record: A Weekly Review of Medicine. 83: 787–790.
- ↑ H. Braun (1903). "Ueber den Einfluss der Vitalität der Gewebe auf die örtlichen und allgemeinen Giftwirkungen localanästhesirender Mittel und über die Bedeutung des Adrenalins für die Localanästhesie". Archiv für Klinische Chirurgie. 69: 541–591.
- ↑ Ladislaus Szymonowicz (1895). "Die Function der Nebenniere". Archiv für die gesamte Physiologie des Menschen und der Tiere. 64 (3–4): 97–164. doi:10.1007/BF01661663. S2CID 29884198.
- ↑ W. H. Bates (1896). "The use of extract of suprarenal capsule in the eye". New York MedicalJournal: 647–650.
- ↑ Erich Spengler (1905). "Kritisches Sammelreferat über die Verwendung einiger neuerer Arzneimittel in der Augenheilkunde". Ophthalmologica. 13: 33–47. doi:10.1159/000290295.
- ↑ John J. Abel (1899–1900). "Ueber den blutdrucksteigernden Bestandtheil der Nebenniere, das Epinephrin". Zeitschrift für physiologische Chemie. 28 (3–4): 318–361. doi:10.1515/bchm2.1899.28.3-4.318.
- ↑ Otto v. Fürth (1900). "Zur Kenntniss der brenzcatechinähnlichen Substanz der Nebennieren". Zeitschrift für physiologische Chemie. 29 (2): 105–123. doi:10.1515/bchm2.1900.29.2.105.
- ↑ Jokichi Takamine (1901). "Adrenalin the active principle of the suprarenal glands and its mode of preparation". The American Journal of Pharmacy. 73: 523–535.
- ↑ E. Friedmann (1906). "Die Konstitution des Adrenalins". Beiträge zur chemischen Physiologie und Pathologie. 8: 95–120.
- ↑ Arthur R. Cushney (1908). "The action of optical isomers". The Journal of Physiology. 37 (2): 130–138. doi:10.1113/jphysiol.1908.sp001261. PMC 1533541. PMID 16992920.
- ↑ E. M. Tansey (1995). "What's in a name? Henry Dale and adrenaline, 1906". Medical History. 39 (4): 459–476. doi:10.1017/s0025727300060373. PMC 1037030. PMID 8558993.
- ↑ M. Lewandowsky (1899). "Ueber die Wirkung des Nebennierenextractes auf die glatten Muskeln, im Besonderen des Auges". Archiv für Anatomie und Physiologie, Archiv für Physiologie: 360–366.
- ↑ T. R. Elliott (1904). "On the action of adrenalin". The Journal of Physiology. 31 (Suppl): XX–XXI. doi:10.1113/jphysiol.1904.sp001055. PMC 1465436.
- ↑ L. Stjärne, P. Hedqvist, H. Lagercrantz, Å. Wennmalm (Eds): Chemical Neurotransmission 75 Years. London, Academic Press, 1981, p. XIII.
- ↑ O. Loewi (1921). "Über humorale Übertragbarkeit der Herznervenwirkung. I. Mitteilung". Pflügers Archiv für die gesamte Physiologie des Menschen und der Tiere. 189: 239–242. doi:10.1007/BF01738910. S2CID 52828335.
- ↑ O. Loewi (1922). "Über humorale Übertragbarkeit der Herznervenwirkung. II. Mitteilung". Pflügers Archiv für die gesamte Physiologie des Menschen und der Tiere. 193: 201–213. doi:10.1007/BF02331588. S2CID 34861770.
- ↑ O. Loewi (1936). "Quantitative und qualitative Untersuchungen über den Sympathicusstoff". Pflügers Archiv für die gesamte Physiologie des Menschen und der Tiere. 237: 504–514. doi:10.1007/BF01753035. S2CID 41787500.
- 1 2 G. Barger; H. H. Dale (1910). "Chemical structure and sympathomimetic action of amines". The Journal of Physiology. 41 (1–2): 19–59. doi:10.1113/jphysiol.1910.sp001392. PMC 1513032. PMID 16993040.
- 1 2 Z. M. Bacq ZM: Chemical transmission of nerve impulses. In: M. J. Parnham, J. Bruinvels (Eds.): Discoveries in Pharmacology. Amsterdam, Elsevier, 1983, vol. 1, pp 49–103. ISBN 0-444-80493-5.
- 1 2 H. Blaschko: Catecholamines 1922–1971. In: H. Blaschko und E. Muscholl (Ed.): Catcholamines. Handbuch der experimentellen Pharmakologie volume 33. Berlin, Springer-Verlag, 1972, pp. 1–15. ISBN 0-387-05517-7.
- ↑ H. H. Dale (1934). "Nomenclature of fibres in the autonomic system and their effectsle". The Journal of Physiology. 80 (Suppl): 10P–11. doi:10.1113/jphysiol.1934.sp003110. PMC 1394004.
- 1 2 Herman Blaschko (1987). "A half-century of research on catecholamine biosynthesis". Journal of Applied Cardiology: 171–183.
- ↑ Peter Holtz; Rudolf Heise; Käthe Lüdtke (1939). "Fermentativer Abbau von l-Dioxyphenylalanin (Dopa) durch Niere". Naunyn-Schmiedebergs Archiv für Pharmakologie und Experimentelle Pathologie. 191: 87–118. doi:10.1007/BF01994628. S2CID 46334627.
- ↑ Hermann Blaschko (1939). "The specific action of l-dopa decarboxylase". The Journal of Physiology. 96 (Suppl): 50 P–51 P. doi:10.1113/jphysiol.1939.sp003786. PMC 1393737.
- ↑ P. Holtz (1939). "Dopadecarboxylase". Die Naturwissenschaften. 27 (43): 724–725. Bibcode:1939NW.....27..724H. doi:10.1007/bf01494245.
- ↑ Edith Bülbring (1949). "The methylation of noradrenaline by minced suprarenal tissue". British Journal of Pharmacology. 4 (3): 234–244. doi:10.1111/j.1476-5381.1949.tb00542.x. PMC 1509915. PMID 18141084.
- ↑ Julius Axelrod (1962). "Purification and properties of phenylethanolamine-N-methyl transferase". The Journal of Biological Chemistry. 237 (5): 1657–1660. doi:10.1016/S0021-9258(19)83758-4. PMID 13863458.
- ↑ Mary Lilias Christian Hare (1928). "Tyramine oxidase. I. A new enzyme system in liver". Biochemical Journal. 22 (4): 968–979. doi:10.1042/bj0220968. PMC 1252213. PMID 16744124.
- ↑ Hermann Blaschko; Derek Richter & Hans Schlossmann (1937). "The oxidation of adrenaline and other amines". Biochemical Journal. 31 (12): 2187–2196. doi:10.1042/bj0312187. PMC 1267198. PMID 16746563.
- ↑ Julius Axelrod (1962). "O-Methylation of epinephrine and other catechols". Science. 126 (3270): 400–401. doi:10.1126/science.126.3270.400. PMID 13467217.
- ↑ U. S. v. Euler (1945). "A sympathomimetic pressor substance in animal organ extracts". Nature. 156 (3949): 18–19. Bibcode:1945Natur.156...18V. doi:10.1038/156018b0. S2CID 4100718.
- 1 2 U. S. v. Euler (1946). "A specific sympathomimetic ergone in adrenergic nerve fibres (sympathin) and its relations to adrenaline and nor-adrenaline". Acta Physiologica Scandinavica. 12: 73–97. doi:10.1111/j.1748-1716.1946.tb00368.x.
- ↑ W. S. Peart (1938). "The nature of splenic sympathin". The Journal of Physiology. 108 (4): 491–501. doi:10.1113/jphysiol.1949.sp004352. PMC 1392468. PMID 16991880.
- 1 2 Peter Holtz; Kartl Credner & Günther Kroneberg (1947). "Über das sympathicomimetische pressorische Prinzip des Harns ("Urosympathin")". Naunyn-Schmiedebergs Archiv für Pharmakologie und Experimentelle Pathologie. 204: 228–243. doi:10.1007/BF00738347. S2CID 43098228.
- ↑ Henry Hallett Dale: Adventures in Pharmacology. With Excursions into Autopharmacology. Pergamon Press, London 1953, p. 98.
- ↑ Marthe Vogt (1954). "The concentration of sympathin in different parts of the central nervous system under normal conditions and after the administration of drugs". The Journal of Physiology. 123 (3): 451–481. doi:10.1113/jphysiol.1954.sp005064. PMC 1366219. PMID 13152692.
- ↑ A. Dahlström; K. Fuxe (1964). "Evidence for the existence of monoamine neurons in the central nervous system. I. Demonstration of monoamines in the cell bodies of brain stem neurons". Acta Physiologica Scandinavica. 62, suppl. 247: 1–55.
- ↑ Å. Bertler; E. Rosengren (1959). "Occurrence and distribution of dopamine in brain and other tissues". Experientia. 15 (1): 10–11. doi:10.1007/bf02157069. PMID 13619664. S2CID 1970402.
- ↑ Arvid Carlsson (1959). "The occurrence, distribution and physiological role of catcholamines in the nervous system". Pharmacological Reviews. 11 (2, Part 2): 490–493. PMID 13667431.
- ↑ H. Ehringer; O. Hornykiewicz (1960). "erteilung von Noradrenalin und Dopamin (3-Hydroxytyramin) im Gehirn des Menschen und ihr Verhalten bei Erkrankungen des extrapyramidalen Systems". Klinische Wochenschrift. 38 (24): 1236–1239. doi:10.1007/BF01485901. PMID 13726012. S2CID 32896604.
- ↑ Oleh Hornykiewicz: From dopamine to Parkinson’s disease: a personal research record. In: Fred Samson, George Adelman (Eds.): The Neurosciences: Paths of Discovery II. Basel, Birkhäuser, 1992, pp. 125–147.
- 1 2 H. Blaschko; A. D. Welch (1953). "Localization of adrenaline in cytoplasmic particles of the bovine adrenal medulla". Naunyn-Schmiedebergs Archiv für Pharmakologie und Experimentelle Pathologie. 219 (1–2): 17–22. doi:10.1007/BF00246245. PMID 13099333. S2CID 26532902.
- ↑ Nils-Åke Hillarp; Sten Lagerstedt; Bodil Nilson (1954). "The isolation of a granular fraction from the suprarenal medulla, containing the sympathomimetic catechol amines". Acta Physiologica Scandinavica. 29 (2–3): 251–263. doi:10.1111/j.1748-1716.1953.tb01022.x. PMID 13114000.
- ↑ U. S. von Euler; N. Å. Hillarp (1956). "Evidence for the presence of noradrenaline in submicroscopic structures of adrenergic axons". Nature. 177 (4497): 44–45. Bibcode:1956Natur.177...44E. doi:10.1038/177044b0. PMID 13288591. S2CID 4214745.
- ↑ E. de Robertis; Amanda Pellegrino de Iraldi; Georgina Rodríguez de Lores Arnaiz; Luis M. Zieher (1965). "Synaptic vesicles from the rat hypothalamus. Isolation and norepinephrine content". Life Sciences. 4 (2): 193–201. doi:10.1016/0024-3205(65)90119-0. PMID 14288585.
- ↑ H. J. Schümann; H. Grobecker (1958). "Über den Noradrenalin- und ATP-Gehalt sympathischer Nerven". Naunyn-Schmiedebergs Archiv für Pharmakologie und Experimentelle Pathologie. 233 (3): 296–300. doi:10.1007/BF00245643. S2CID 29615845.
- ↑ Å. Thureson-Klein (1983). "Exocytosis from large and small dense cored vesicles in noradrenergic nerve terminals". Neuroscience. 10 (2): 245–252. doi:10.1016/0306-4522(83)90132-X. PMID 6633860. S2CID 22592654.
- ↑ J. Axelrod; H. Weil-Malherbe; R. Tomchick (1959). "The physiological disposition of H3-epinephrine and its metabolite metanephrine". Journal of Pharmacology and Experimental Therapeutics. 127: 251–256. PMID 13795315.
- ↑ L. G. Whitby; J. Axelrod; H. Weil-Malherbe (1961). "The fate of H3-norepinephrine in animals". Journal of Pharmacology and Experimental Therapeutics. 132: 193–201. PMID 13784801.
- ↑ A. Fröhlich; O. Loewi (1910). "Über eine Steigerung der Adrenalinempfindlichkeit durch Cocaïn". Archiv für experimentelle Pathologie und Pharmakologie. 62 (2–3): 159–169. doi:10.1007/BF01840652. S2CID 43503888.
- ↑ E. Muscholl (1961). "Effect of cocaine and related drugs on the uptake of noradrenaline by heart and spleen". British Journal of Pharmacology. 16 (3): 352–359. doi:10.1111/j.1476-5381.1961.tb01095.x. PMC 1482029. PMID 13727081.
- ↑ G. Hertting; J. Axelrod (1961). "Fate of tritiated noradrenaline at the sympathetic nerve-endings". Nature. 192 (4798): 172–173. Bibcode:1961Natur.192..172H. doi:10.1038/192172a0. PMID 13906919. S2CID 4188539.
- ↑ Leslie L. Iversen: The Uptake and Storage of Noradrenaline in Sympathetic Nerves. University Press, Cambridge 1967.
- ↑ U. Trendelenburg (1986). "The metabolizing systems involved in the inactivation of catecholamines". Naunyn-Schmiedeberg's Archives of Pharmacology. 332 (3): 201–297. doi:10.1007/BF00504854. PMID 3713866. S2CID 44614695.
- ↑ Robert Frederiksson; Malin C. Lagerström; Lars-Gustav Lundin; Helgi B. Schiöth (2003). "The G-protein-coupled receptors in the human genome form five main families. Phylogenetic analysis, paralogon groups, and fingerprints". Molecular Pharmacology. 63 (6): 1256–1272. doi:10.1124/mol.63.6.1256. PMID 12761335.
- ↑ H. H. Dale (1906). "On some physiological actions of ergot". The Journal of Physiology. 34 (3): 163–206. doi:10.1113/jphysiol.1906.sp001148. PMC 1465771. PMID 16992821.
- ↑ Max Hartmann; Hans Isler (1939). "Chemische Konstitution und pharmakologische Wirksamkeit von in 2-Stellung substituierten Imidazolinen". Naunyn-Schmiedebergs Archiv für Pharmakologie und Experimentelle Pathologie. 192 (2–5): 141–154. doi:10.1007/BF01924807. S2CID 40545320.
- ↑ Mark Nickerson (1949). "The pharmacology of adrenergic blockade". Pharmacological Reviews. 1: 27–101.
- ↑ Heribert Konzett; Richard Rössler (1940). "Versuchsanordnung zu Untersuchungen an der Bronchialmuskulatur". Naunyn-Schmiedebergs Archiv für Pharmakologie und Experimentelle Pathologie. 195: 71–74. doi:10.1007/BF01861842. S2CID 27059235.
- ↑ Heribert Konzett (1941). "Neue broncholytisch hochwirksame Körper der Adrenalinreihe". Naunyn-Schmiedebergs Archiv für Pharmakologie und Experimentelle Pathologie. 197: 27–40. doi:10.1007/BF01936304. S2CID 5181392.
- ↑ H. Konzett (1981). "On the discovery of isoprenaline". Trends in Pharmacological Sciences. 2: 47–49. doi:10.1016/0165-6147(81)90259-5.
- ↑ Walter Sneader: Drug Discovery: The Evolution of Modern Medicines. John Wiley & Sons, Chichester, 1985, S. 103. ISBN 0 471 90471 6.
- 1 2 Raymond P. Ahlquist (1948). "A study of the adrenotropic receptors". American Journal of Physiology. 153 (3): 586–600. doi:10.1152/ajplegacy.1948.153.3.586. PMID 18882199. S2CID 1518772.
- ↑ Ivar von Kügelgen; Klaus Starke (1991). "Noradrenalin-ATP co-transmission in the sympathetic nervous system". Trends in Pharmacological Sciences. 12 (9): 319–324. doi:10.1016/0165-6147(91)90587-I. PMID 1658999.
- ↑ C. E. Powell; I. H. Slater (1958). "Blocking of inhibitory adrenergic receptors by a dichloro analog of isoproterenol". Journal of Pharmacology and Experimental Therapeutics. 122 (4): 480–488. PMID 13539775.
- ↑ Neil C. Moran; Marjorie E. Perkins (1958). "Adrenergic blockade of the mammalian heart by a dichloro analogue of isoproterenol". Journal of Pharmacology and Experimental Therapeutics. 124: 222–237.
- ↑ J. W. Black; A. F. Crowther; R. G. Shanks; A. C. Dornhorst (1964). "A new adrenergic beta-receptor antagonist". The Lancet. 283 (7342): 1080–1081. doi:10.1016/S0140-6736(64)91275-9. PMID 14132613.
- ↑ A. M. Lands; A. Arnold; J. P. McAuliff; F. P. Luduena; T. G. Brown (1967). "Differentiation of receptor systems activated by sympathomimetic amines". Nature. 214 (5088): 597–598. Bibcode:1967Natur.214..597L. doi:10.1038/214597a0. PMID 6036174. S2CID 4165965.
- ↑ Johan Zaagsma; Stefan R. Nahorski (1990). "Is the adipocyte β-adrenoceptor a prototype of the recently cloned β3-adrenoceptor?". Trends in Pharmacological Sciences. 11 (1): 3–7. doi:10.1016/0165-6147(90)90032-4. PMID 2155496.
- ↑ Anthony P. Nicholas; Vincent Pieribone; Tomas Hökfelt (1993). "Distributions of mRNAs for alpha-2 adrenergic receptor subtypes in rat brain: an in situ hybridization study". Journal of Comparative Neurology. 328 (4): 575–594. doi:10.1002/cne.903280409. PMID 8381444. S2CID 25109961.
- ↑ Klaus Starke (2001). "Presynaptic autoreceptors in the third decade: focus on α2-adrenoceptors". Journal of Neurochemistry. 78 (4): 685–693. doi:10.1046/j.1471-4159.2001.00484.x. PMID 11520889. S2CID 82607249.
- ↑ Ralf Gilsbach, Lutz Hein: Presynaptic metabotropic receptors for acetylcholine and adrenaline/noradrenaline. In: Thomas C. Südhoff, Klaus Starke (Eds.): Pharmacology of Neurotransmitter Release. Handbook of Experimental Pharmacology 184. Springer, Berlin 2008, pp. 261–288. ISBN 978-3-540-74804-5.
- ↑ Salomón Z. Langer (1974). "Presynaptic regulation of catecholamine release". Biochemical Pharmacology. 23 (13): 1793–1800. doi:10.1016/0006-2952(74)90187-7. PMID 4617579.
- ↑ Peter Holtz; Karl Credner; Wolfgang Koepp (1942). "Die enzymatische Entstehung von Oxytyramin im Organismus und die physiologische Bedeutung der Dopadecarboxylase". Naunyn-Schmiedebergs Archiv für Pharmakologie und Experimentelle Pathologie. 200 (2–5): 356–388. doi:10.1007/BF01860725. S2CID 34006359.
- ↑ John Nelson Eble (1964). "A proposed mechanism for the depressor effect of dopamine in the anesthetized dog". Journal of Pharmacology and Experimental Therapeutics. 145: 64–70. PMID 14209513.
- ↑ Richard A. F. Dixon; Brian K. Kobilka; David J. Strader; et al. (1986). "Cloning of the gene and cDNA for mammalian β-adrenergic receptor and homology with rhodopsin". Nature. 321 (6065): 75–79. Bibcode:1986Natur.321...75D. doi:10.1038/321075a0. PMID 3010132. S2CID 4324074.
- ↑ Daniel M. Rosenbaum; Cheng Zhang; Joseph A. Lyons; et al. (2011). "Structure and function of an irreversible agonist-β2 adrenoceptor complex". Nature. 469 (7329): 236–240. Bibcode:2011Natur.469..236R. doi:10.1038/nature09665. PMC 3074335. PMID 21228876.
Further reading
- Paul Trendelenburg: Adrenalin und adrenalinverwandte Substanzen. In: Arthur Heffter (Ed.): Handbuch der experimentellen Pharmakologie volume 2 part 2. Berlin, Julius Springer 1924, p. 1130–1293.
- H. Blaschko: Catecholamines 1922–1971. In: H. Blaschko und E. Muscholl (Ed.): Catcholamines. Handbuch der experimentellen Pharmakologie volume 33. Berlin, Springer-Verlag, 1972, pp. 1–15. ISBN 0-387-05517-7.
- Herman Blaschko (1987). "A half-century of research on catecholamine biosynthesis". Journal of Applied Cardiology: 171–183.
- Zénon M. Bacq: Chemical transmission of nerve impulses. In: M. J. Parnham, J. Bruinvels (Eds.): Discoveries in Pharmacology. Volume 1: Psycho- and Neuropharmacology, Amsterdam, Elsevier, 1983, pp. 49–103. ISBN 0-444-80493-5.
- M. R. Bennett (1999). "One hundred years of adrenaline: the discovery of autoreceptors". Clinical Autonomic Research. 9 (3): 145–149. doi:10.1007/BF02281628. PMID 10454061. S2CID 20999106.
- Josef Donnerer, Fred Lembeck: Adrenaline, noradrenaline and dopamine: the catecholamines. In: The Chemical Languages of the Nervous System. Basel, Karger, 2006, p. 150–160.