Hyalospheniidae
| Hyalospheniidae Temporal range: | |
|---|---|
 | |
| Hyalosphenia papilio | |
| Scientific classification | |
| Domain: | Eukaryota |
| Phylum: | Amoebozoa |
| Class: | Tubulinea |
| Order: | Arcellinida |
| Suborder: | Glutinoconcha |
| Infraorder: | Hyalospheniformes Lahr et al. 2019[1] |
| Family: | Hyalospheniidae Schulze, 1877[2] emend. Kosakyan & Lara, 2012[3] |
| Type genus | |
| Hyalosphenia Stein 1857 | |
| Genera | |
|
14 (see text) | |
| Diversity | |
| 78 species | |
| Synonyms[4] | |
Hyalospheniidae is a family of arcellinid testate amoebae and the sole family of the infraorder Hyalospheniformes. Commonly referred to as "hyalospheniids", these lobose amoebae are characterized by their ability to generate a shell composed of either organic matter or siliceous particles that may be recycled from euglyphid amoebae. They inhabit soil or freshwater habitats, and are abundant on Sphagnum mosses.
Hyalospheniid amoebae originated after the middle Devonian, around 370 million years ago. They are considered important bioindicators, and are frequently used for environmental monitoring. Their fossils are studied to investigate the paleoecology of prehistoric wetland habitats. The classification of hyalospheniids has changed several times since the 19th century based on morphological criteria. Initially classified as two separate families, Hyalospheniidae and Nebelidae, they were later proven to be synonymous through phylogenetic analyses.
Morphology
Hyalospheniidae are testate amoebae—unicellular amoeboid protists that generate mineral agglutinated shells. They are characterized by ovoid, pyriform, vase or flask-shaped shells, which are laterally compressed.[6][7] Shell construction and composition varies substantially within the family. It can be either entirely secreted by their own cells and composed of an organic matrix (e.g. Hyalosphenia), or have additional non-organic siliceous scales. These mineral scales can also be self-secreted (e.g. Quadrulella), or can be recycled from the shell plates of small euglyphid amoebae or other similar material such as diatom frustules (e.g. Apodera, Padaungiella, Nebela).[6] The trait of recycling shell plates from euglyphids is known as "kleptosquamy", and appears to be an ancestral trait within the family.[8]
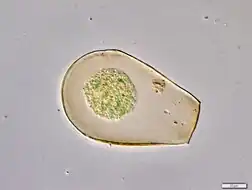
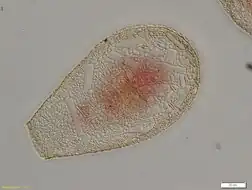
Ecology
Hyalospheniid amoebae are considered important bioindicators in environmental monitoring studies. Their sensitivity to environmental changes, such as atmospheric pollution,[9] make them reliable indicators of hydrological changes. Together with the preservation of their shells over thousands of years, their environmental sensitivity gives them a prominent role in the reconstruction of the paleoclimate in peatlands, bogs and fens.[6]
This family includes several of the most common, well-studied lobose testate amoebae. Its members are especially diverse and abundant in oligotrophic wetland ecosystems, such as peatlands dominated by Sphagnum mosses. Some of them can also be found in different mosses, freshwater habitats, and soil. Although multiple hyalospheniid species have a cosmopolitan distribution, many species are restricted to the Southern hemisphere and the tropical range within the Northern hemisphere.[7]
Evolution
Hyalospheniidae is a family of Arcellinida, an order of lobose testate amoebae within the eukaryotic supergroup Amoebozoa. In contrast to filose testate amoebae, found within the supergroup Rhizaria (e.g. euglyphid amoebae), they present thicker pseudopods with blunt ends. It is the only family of the infraorder Hyalospheniformes, which belongs to the suborder Glutinoconcha. Glutinoconcha, which contains the majority of arcellinid species, evolved from a common ancestor with mineral agglutinated shells, in contrast to the organic shells of Organoconcha. In particular, Hyalospheniformes and Volnustoma, a different infraorder of Glutinoconcha, both evolved from ancestors with xenosomic agglutinated shells (i.e. composed of particles incorporated from an external source).[1][10]
| Arcellinida |
| ||||||||||||||||||||||||||||||||||||||||||
Through molecular clock approximations, the age of Hyalospheniidae was estimated in 2015 to be around 370 million years old, between the Devonian and the early Carboniferous. This molecular reconstruction suggests that hyalospheniids diversified after the middle Devonian, when the diversification of land plants formed extensive forests with an abundant production of organic matter and soils. Kleptosquamy, the ability of hyalosphenid amoebae to "steal" test scales from their prey, euglyphid amoebae, is hypothesized to be an ancestral trait within the family. This working hypothesis is based on the presence of kleptosquamy on most hyalospheniids.[8] In addition to the molecular clock estimates, it has been suggested that 750–million-year-old vase-shaped microfossils could belong to this family.[11][6][12]
Systematics
History of taxonomy
There has been several attempts at classifying Hyalospheniidae among testate amoebae, as well as its internal classification. American paleontologist Joseph Leidy, in 1874, was possibly the first to notice common characteristics between the cells. He described the vase-shaped tests as composed of small siliceous particles ("discoid plates and minute rods") caught within an organic matrix, interpreted to be originated by the amoeba ("intrinsic"). He grouped those species within the genus Nebela, restricting them from the previously known genus of testate amoebae Difflugia. Instead, he described Difflugia species as having a "test composed of extraneous bodies, such as particles of quartzose sand, and diatom cases".[13][6]
In 1877, German zoologist Franz Eilhard Schulze described the families Hyalosphenidae, Arcellidae, Quadrulidae and Difflugidae. Amoebae with an organic homogenous test such as Hyalosphenia were placed in Hyalospheniidiae, while Nebela was placed in Difflugidae, and Quadrulella in Quadrulidae.[2][6]
In 1882, Taranek described the family Nebelidae to include amoebae with siliceous plates: Nebela, Lesquereusia, Quadrulella, Corythion (which was later excluded), Amphizonella, Cochliopodium, Hyalosphenia, Leptochlamys and Zonomyxa.[5] This family was redefined later in 1942 by Jung and organized into thirteen genera newly described by him: Alocodera, Apodera, Argynnia, Deflandria, Nebela, Leidyella, Penardiella, Physochila, Porosia, Pterygia, Quadrulella, Schaudinnia and Umbonaria.[14] However, Jung did not designate type species in his classification, which invalidated all genera with more than one species.[lower-alpha 1] Consequently, only monotypic genera such as Alocodera, Physochila and Porosia were recognized, and the remaining genera were absorbed by Nebela. Later, by assigning types, micropaleontologists Loeblich and Tappan validated Apodera and Certesella in 1961,[16] and Vucetich validated Argynnia in 1974.[17]
In 2002, German protozoologist Ralf Meisterfeld wrote the last review of the family based exclusively on morphological characters. He reclassified Nebela and similar genera into two families: Hyalospheniidae, composed of genera with rigid, chitinoid, organic tests (Hyalosphenia and Leptochlamys); and Nebelidae, composed of genera with tests constructed from plates of small euglyphids or diatom fragments (Apodera, Argynnia, Certesella, Nebela, Physochila, Porosia, Schoenbornia). Following a 1979 classification,[18] he excluded the genus Quadrulella into the family Lesquereusiidae along with other arcellinid genera with self-secreted siliceous rods in their tests.[19][6]
The first phylogenetic analyses of Arcellinida based on molecular data demonstrated that Nebela and Nebelidae were not monophyletic, i.e. did not form a clade or group of taxa evolved from a common ancestor without including other genera from Hyalospheniidae.[20] Species of Apodera, Porosia, Nebela and Hyalosphenia were intermingled with each other in a clade known as "core Nebelas".[7] Because of this result, and presence of distinguishing Hyalospheniidae traits within some Nebelidae, the two families were synonymised under the first name.[3] In addition, many species of the polyphyletic Nebela were separated into new monophyletic genera, namely Padaungiella in 2012,[3] Cornutheca, Gibbocarina, Longinebela, Mrabella and Planocarina in 2016,[21] and finally Alabasta in 2018.[22]
Molecular phylogeny
The following cladogram illustrates the evolutionary relationships between all hyalospheniid genera found through a 2018 phylogenetic analysis,[22] with the exception of Porosia, a genus excluded from the analysis that appears to be closely related to Certesella[23] and is therefore placed next to it in the cladogram. Another genus excluded by the same analysis is Apodera, which is the sister group to a clade containing Alocodera and Padaungiella, according to a 2021 phylogeny.[24]
| Hyalospheniidae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
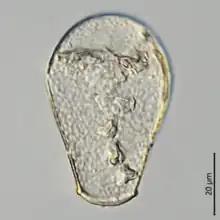
Genera
The current taxonomy of the family recognizes 14 genera,[1][25] with a total of 78 species:
- Certesella Loeblich & Tappan, 1961 — 5 species.[29][30]
- Cornutheca Kosakyan, Lahr, Mulot, Meisterfeld, Mitchell & Lara, 2016 — 3 species.[21]
- Hyalosphenia (Stein, 1857) Schulze, 1877 — 17 species.[6]
- Mrabella Kosakyan, Lahr, Mulot, Meisterfeld, Mitchell & Lara, 2016 — 2 species.[21]
- Padaungiella Lara et Todorov, 2012 — 5 species.[3]
- Planocarina Kosakyan, Lahr, Mulot, Meisterfeld, Mitchell & Lara, 2016 — 4 species.[21]
- Quadrulella Cockerell, 1909 em. Kosakyan, Lahr, Mulot, Meisterfeld, Mitchell & Lara, 2016 — 11 species.[21]
Notes
References
- 1 2 3 Lahr D, Kosakyan A, Lara E, Mitchell E, Morais L, Porfirio-Sousa AL, Ribeiro GM, Tice AK, Pánek T, Kang S, Brown MW (2019). "Phylogenomics and Morphological Reconstruction of Arcellinida Testate Amoebae Highlight Diversity of Microbial Eukaryotes in the Neoproterozoic". Current Biology. 29 (6): 991–1001. doi:10.1016/j.cub.2019.01.078. PMID 30827918. S2CID 72333352. Archived from the original on 2022-06-29. Retrieved 2022-07-13.
- 1 2 Schultze FE (1877). "Rhizopodenstudien VI". Archiv für Mikroskopische Anatomie. 13: 9–30. doi:10.1007/BF02933929. Archived from the original on 2022-10-25. Retrieved 2023-09-24.
- 1 2 3 4 Kosakyan A, Heger TJ, Leander BS, Todorov M, Mitchell EAD, Lara E (2012). "COI Barcoding of Nebelid Testate Amoebae (Amoebozoa: Arcellinida): Extensive Cryptic Diversity and Redefinition of the Hyalospheniidae Schultze". Protist. 163 (3): 415–434. doi:10.1016/j.protis.2011.10.003. ISSN 1434-4610. PMID 22130576. Archived from the original on 2023-10-22. Retrieved 2022-11-23.
- ↑ Kosakyan, Anush; Gomaa, Fatma; Lara, Enrique; Lahr, Daniel J.G. (2016). "Current and future perspectives on the systematics, taxonomy and nomenclature of testate amoebae". European Journal of Protistology. 55B (Pt B): 105–117. doi:10.1016/j.ejop.2016.02.001. ISSN 0932-4739. PMID 27004416. Archived from the original on 2023-10-22. Retrieved 2022-07-13.
- 1 2 Taranek KJ (1882). "Monographie der Nebeliden Böhmens. Ein Beitrag zur Kenntnis der Süsswasser Monothalamien" [Monograph of the Nebelids of Bohemia. A contribution to the knowledge of freshwater monothalamids]. Abhandlungen der Königlichen Böhmischen Gesellschaft der Wissenschaften (in German). VI: 1–56.
- 1 2 3 4 5 6 7 8 9 10 Kosakyan, Anush (16 April 2014). Phylogeny, systematics and ecology of free living protists. Case study: family Hyalospheniidae (Thesis). University of Neuchâtel. Archived from the original on 29 September 2023. Retrieved 24 September 2023.
- 1 2 3 Lara E, Heger TJ, Ekelund F, Lamentowicz M, Mitchell EAD (2008). "Ribosomal RNA genes challenge the monophyly of the Hyalospheniidae (Amoebozoa: Arcellinida)". Protist. 159 (2): 165–176. doi:10.1016/j.protis.2007.09.003. PMID 18023614. Archived from the original on 2021-06-23. Retrieved 2023-10-22.
- 1 2 Lahr, Daniel JG; Bosak, Tanja; Lara, Enrique; Mitchell, Edward AD (September 2015). "The Phanerozoic diversification of silica-cycling testate amoebae and its possible links to changes in terrestrial ecosystems". PeerJ. 3: e1234. doi:10.7717/peerj.1234. PMC 4699787. PMID 26734499.
- ↑ Payne R, Mitchell E, Nguyen-Viet H, Gilbert D (2012). "Can pollution bias peatland paleoclimate reconstruction?". Quaternary Research. 78 (2): 170–173. Bibcode:2012QuRes..78..170P. doi:10.1016/j.yqres.2012.05.004. S2CID 27837542.
- ↑ González-Miguéns, Rubén; Todorov, Milcho; Blandenier, Quentin; Duckert, Clément; Porfirio-Sousa, Alfredo L.; Ribeiro, Giulia M.; Ramos, Diana; Lahr, Daniel J.G.; Buckley, David; Lara, Enrique (2022). "Deconstructing Difflugia: The tangled evolution of lobose testate amoebae shells (Amoebozoa: Arcellinida) illustrates the importance of convergent evolution in protist phylogeny". Molecular Phylogenetics and Evolution. 175: 107557. doi:10.1016/j.ympev.2022.107557. ISSN 1055-7903. PMID 35777650.
- ↑ Porter SM, Meisterfeld R, Knoll AH (May 2003). "Vase-shaped microfossils from the Neoproterozoic Chuar Group, Grand Canyon: a classification guided by modern testate amoebae" (PDF). Journal of Paleontology. 77 (3): 409–429. doi:10.1666/0022-3360(2003)077<0409:VMFTNC>2.0.CO;2. Archived (PDF) from the original on 2018-03-08. Retrieved 2023-09-24.
- ↑ Tingle KE, Porter SM, Raven MR, Czaja AD, Webb SM, Bloeser B (2023). "Organic preservation of vase-shaped microfossils from the late Tonian Chuar Group, Grand Canyon, Arizona, USA". Geobiology. 21 (3): 290–309. doi:10.1111/gbi.12544. PMID 36651474.
- ↑ Leidy J (1874). "Notice of some new freshwater rhizopods" (PDF). Annals and Magazine of Natural History. 14 (83): 383–385. doi:10.1080/00222937408680997.
- ↑ Jung W (1942). "Illustrierte Thekamöben-Bestimmungstabellen I. Die Systematik der Nebelinen" [Illustrated identification tables of thecate amoebae I. The systematics of the nebelines]. Archiv für Protistenkunde (in German). 95: 357–390.
- ↑ "ICZN Code Art. 13". Archived from the original on 5 October 2021. Retrieved 25 September 2023.
- ↑ Loeblich, Alfred Richard; Tappan, Helen Niña (1961). "Remarks on the systematics of the Sarkodina (Protozoa), renamed homonyms and new and validated genera". Proceedings of the Biological Society of Washington. 74: 213–234. Archived from the original on 2023-09-30. Retrieved 2023-09-24.
- ↑ Vucetich MC (1974). "Comentarios criticos sobre Argynnia Jung, 1942 (Rhizopoda, Testacea)" [Critical comments on Argynnia Jung, 1942 (Rhizopoda, Testaeca)]. Neotropica (in Spanish). 20 (63): 126–128.
- ↑ Ogden CG (1979). "Siliceous structures secreted by members of the subclass Lobosia (Rhizopodea, Protozoa)". Bulletin of the British Museum (Natural History) Zoology. 36: 203–207. Archived from the original on 2023-10-03. Retrieved 2023-10-22.
- ↑ Meisterfeld R (2002). "Order Arcellinida Kent, 1880" (PDF). In Lee JJ, Leedale GF, Bradbury P (eds.). An illustrated guide to the protozoa: organisms traditionally referred to as protozoa, or newly discovered groups (2nd ed.). Lawrence, Kansas, USA: Society of protozoologists. pp. 827–860. Archived from the original on 2023-09-29. Retrieved 2023-09-25.
- ↑ Nikolaev SI, Mitchell EA, Petrov NB, Berney C, Fahrni J, Pawlowski J (2005). "The testate lobose amoebae (order Arcellinida Kent, 1880) finally find their home within Amoebozoa". Protist. 156 (2): 191–202. doi:10.1016/j.protis.2005.03.002. PMID 16171186.
- 1 2 3 4 5 6 7 8 Kosakyan, Anush; Lahr, Daniel J. G.; Mulot, Matthieu; Meisterfeld, Ralf; Mitchell, Edward A. D.; Lara, Enrique (2016). "Phylogenetic reconstruction based on COI reshuffles the taxonomy of hyalosphenid shelled (testate) amoebae and reveals the convoluted evolution of shell plate shapes". Cladistics. 32 (6): 606–623. doi:10.1111/cla.12167. PMID 34727671.
- 1 2 3 Duckert C, Blandenier Q, Kupferschmid FAL, Kosakyan A, Mitchell EAD, Lara E, Singer D (2018). "En garde! Redefinition of Nebela militaris (Arcellinida, Hyalospheniidae) and erection of Alabasta gen. nov" (PDF). European Journal of Protistology. 66: 156–165. doi:10.1016/j.ejop.2018.08.005. PMID 30366198. S2CID 53116247. Archived (PDF) from the original on 2020-06-10. Retrieved 2023-01-03.
- 1 2 Bobrov, Anatoly; Kosakyan, Anush. "A New Species from Mountain Forest Soils in Japan: Porosia paracarinata sp. nov., and Taxonomic Concept of the Genus Porosia Jung, 1942". Acta Protozoologica. 54 (4): 289–294. doi:10.4467/16890027AP.15.024.3538.
- ↑ Duckert C, Blandenier Q, McKeown M, Hohaia H, Luketa S, Wilmshurst J, et al. (2021). "Superficially described and ignored for 92 years, rediscovered and emended: Apodera angatakere (Amoebozoa: Arcellinida: Hyalospheniformes) is a new flagship testate amoeba taxon from Aotearoa (New Zealand)". Journal of Eukaryotic Microbiology. 68 (6): e12867. doi:10.1111/jeu.12867. PMC 9292727. PMID 34351666.
- ↑ González-Miguéns, Rubén; Todorov, Milcho; Blandenier, Quentin; Duckert, Clément; Porfirio-Sousa, Alfredo L.; Ribeiro, Giulia M.; Ramos, Diana; Lahr, Daniel J.G.; Buckley, David; Lara, Enrique (2022). "Deconstructing Difflugia: The tangled evolution of lobose testate amoebae shells (Amoebozoa: Arcellinida) illustrates the importance of convergent evolution in protist phylogeny". Molecular Phylogenetics and Evolution. 175: 107557. doi:10.1016/j.ympev.2022.107557. ISSN 1055-7903. PMID 35777650.
- 1 2 Jung W (1942). "Südchilenische Thekamöben". Archiv für Protistenkunde. 95: 253–356.
- ↑ Luketa, Stefan (2015). "Description of the family Padaungiellidae and morphological variability of Padaungiella lageniformis (Amoebozoides: Arcellinida) from the Vlasina Lake area, Serbia". Archives of Biological Sciences, Belgrade. 67 (4): 1331–1337. doi:10.2298/ABS150312110L.
- ↑ Duckert C, Blandenier Q, McKeown M, Hohaia H, Luketa S, Wilmshurst J, et al. (2021). "Superficially described and ignored for 92 years, rediscovered and emended: Apodera angatakere (Amoebozoa: Arcellinida: Hyalospheniformes) is a new flagship testate amoeba taxon from Aotearoa (New Zealand)". Journal of Eukaryotic Microbiology. 68 (6): e12867. doi:10.1111/jeu.12867. PMC 9292727. PMID 34351666.
- ↑ Bobrov, Anatoly; Duckert, Clément; Mitchell, Edward A. D. (2021). "Certesella larai (Amoebozoa: Arcellinida: Hyalospheniformes) a new soil testate amoeba species from the Dominican Republic and Chile challenges the definition of genera Certesella and Porosia". Acta Protozoologica. 60: 61–75. doi:10.4467/16890027AP.21.007.15381.
- ↑ Vucetich, María Cristina (1978). "Comentarios sobre el género Certesella Loeblich & Tappan, 1961 y estudio de la estéreo ultraestructura tecal de tres especies austroamericanas (Rhizopoda Testaceolobosa)" [Comments on the genus Certesella Loeblich & Tappan, 1961 and the study of the thecate stereo ultrastructure of three Austroamerican species (Rhizopoda Testacealobosa)] (PDF). Obra Centenario, Museo de la Plata, Zoología (in Spanish). 6: 305–313. Archived (PDF) from the original on 2022-02-20. Retrieved 2023-09-23.
- ↑ Luketa, Stefan (2017). "Morphological variability of Gibbocarina galeata and G. penardiana comb. nov. (Arcellinida: Hyalospheniidae) from East Herzegovina". Protistology. 11 (1): 37–47. doi:10.21685/1680-0826-2017-11-1-3.
- ↑ Todorov, Milcho; Bankov, Nikola; Ganeva, Anna (2018). "Longinebela ampulla sp. n. (Arcellinida: Hyalospheniidae), a New Testate Amoeba from Sphagnum Peatlands in Bulgaria" (PDF). Acta Zoologica Bulgarica. 70 (3): 285–292. Archived (PDF) from the original on 2023-03-13. Retrieved 2023-10-22.