Leishmania
| Leishmania | |
|---|---|
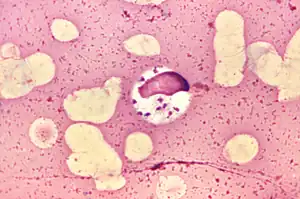 | |
| L. donovani in bone marrow cell | |
| Scientific classification | |
| Domain: | Eukaryota |
| Phylum: | Euglenozoa |
| Class: | Kinetoplastea |
| Order: | Trypanosomatida |
| Genus: | Leishmania Borovsky 1898[1] (Ross 1903) |
| Species | |
|
L. aethiopica | |
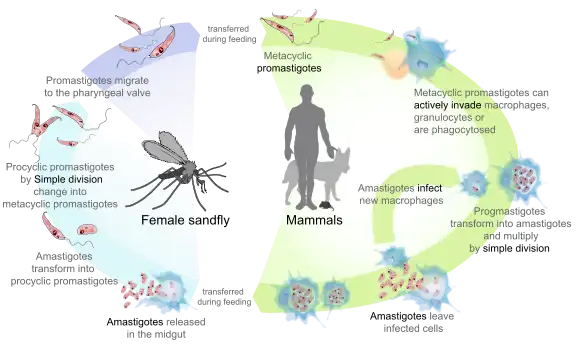
Leishmania /liːʃˈmeɪniə, -ˈmæn-/[2] is a genus of trypanosomes that are responsible for the disease leishmaniasis.[3][4][5] They are spread by sandflies of the genus Phlebotomus in the Old World, and of the genus Lutzomyia in the New World. At least 93 sandfly species are proven or probable vectors worldwide.[6] Their primary hosts are vertebrates; Leishmania commonly infects hyraxes, canids, rodents, and humans.
History

The first written reference to the conspicuous symptoms of cutaneous leishmaniasis surfaced in the Paleotropics within oriental texts dating back to the 7th century BC (allegedly transcribed from sources several hundred years older, between 1500 and 2000 BC[7]). Due to its broad and persistent prevalence throughout antiquity as a mysterious disease of diverse symptomatic outcomes, leishmaniasis has been dubbed with various names ranging from "white leprosy" to "black fever". Some of these names suggest links to negative cultural beliefs or mythology, which still feed into the social stigmatization of leishmaniasis today.[8]
Members of an ancient genus of the Leishmania parasite, Paleoleishmania, have been detected in fossilized sand flies dating back to the early Cretaceous period,[9] however, the causative agent for the disease was only discovered in 1901 as a concurrent finding by William Boog Leishman and Charles Donovan. They independently visualised microscopic single-celled parasites (later called Leishman-Donovan bodies) living within the cells of infected human organs. The parasitic genus would later be classed as trypanosomatid protozoans under the phylogenetic designation, Leishmania donovani. Several species have since been classified and grouped under two major subgenera i.e. Leishmania Viannia (generally located in the Neotropics) or Leishmania Leishmania (generally located in the Paleotropics, with the major exception of the L. mexicana subgroup).
Epidemiology
Leishmania currently affects 6 million people in 98 countries. About 0.9-1.6 million new cases occur each year, and 21 species are known to cause disease in humans: it is considered a zoonosis.
Structure
Leishmania species are unicellular eukaryotes having a well-defined nucleus and other cell organelles including kinetoplasts and flagella. Depending on the stage of their life cycle, they exist in two structural variants, as:[10][11]
- The amastigote form is found in the mononuclear phagocytes and circulatory systems of humans. It is an intracellular and nonmotile form, being devoid of external flagella. The short flagellum is embedded at the anterior end without projecting out. It is oval in shape, and measures 3–6 µm in length and 1–3 µm in breadth. The kinetoplast and basal body lie towards the anterior end.
- The promastigote form is found in the alimentary tract of sandflies. It is an extracellular and motile form. It is considerably larger and highly elongated, measuring 15-30 µm in length and 5 µm in width. It is spindle-shaped, tapering at both ends. A long flagellum (about the body length) is projected externally at the anterior end. The nucleus lies at the centre, and in front of it are the kinetoplast and the basal body.
Evolution
The details of the evolution of this genus are debated, but Leishmania apparently evolved from an ancestral trypanosome lineage. The oldest lineage is that of the Bodonidae, followed by Trypanosoma brucei, the latter being confined to the African continent. Trypanosoma cruzi groups with trypanosomes from bats, South American mammals, and kangaroos suggest an origin in the Southern Hemisphere. These clades are only distantly related.
The remaining clades in this tree are Blastocrithidia, Herpetomonas, and Phytomonas. The four genera Leptomonas, Crithidia, Leishmania, and Endotrypanum form the terminal branches, suggesting a relatively recent origin. Several of these genera may be polyphyletic and may need further division.[12]
The origins of genus Leishmania itself are unclear.[13][14] One theory proposes an African origin, with migration to the Americas. Another proposes migration from the Americas to the Old World via the Bering Strait land bridge around 15 million years ago. A third theory proposes a Palearctic origin.[15] Such migrations would entail subsequent migration of vector and reservoir or successive adaptations along the way. A more recent migration is that of L. infantum from Mediterranean countries to Latin America (known as L. chagasi), since European colonization of the New World, where the parasites picked up their current New World vectors in their respective ecosystems.[16] This is the cause of the epidemics now evident. One recent New World epidemic concerns foxhounds in the USA.[17]
Although it was suggested that Leishmania might have evolved in the Neotropics.[18] this is probably true for species belonging to the subgenera Viannia and Endotrypanum. However, there is evidence that the primary evolution of the subgenera Leishmania and Sauroleishmania is the Old World. While the Mundinia species appear to be more universal in their evolution. One theory is that different lineages became isolated geographically during different periods and it is this that gave rise to this evolutionary mosaicism. But there is no doubt that the Leishmaniinae are a monophyletic group.
A large data set analysis suggests that Leishmania evolved 90 to 100 million years ago in Gondwana.[19] The reptile infecting species originated in mammalian clades.
Sauroleishmania species were originally defined on the basis that they infected reptiles (lizards) rather than mammals. Based on molecular evidences, they have been moved to subgenus status within Leishmania. This subgenus probably evolved from a group that originally infected mammals.[20]
Taxonomy
53 species are recognised in this genus. The status of several of these is disputed, so the final number may differ. At least 20 species infect humans. To make things more complex, hybrids might be involved, as it has been reported in Brazil with a hybrid between Leishmania (V.) guyanensis and Leishmania (V.) shawi shawi.[21]
The genus is presently divided into 4 subgenera: Leishmania, Sauroleishmania, Mundinia and Viannia. The division into the two subgenera (Leishmania and Viannia) was made by Lainson and Shaw in 1987 on the basis of their location within the insect gut. The species in the Viannia subgenus develop in the hind gut: L. (V.) braziliensis has been proposed as the type species for this subgenus. This division has been confirmed by all subsequent studies. Shaw, Camargo and Teixeira created the subgenus Mundinia while revising Leishmaniinae in 2016.[22]
Endotrypanum is closely related to Leishmania. Some Endotypanum species are unique in that they infect the erythrocytes of their hosts (sloths). All species are confined to Central and South America.[23] E. colombiensis infections have been found in man.
Sauroleishmania was originally described by Ranquein 1973 as a separate genus, but molecular studies suggest this is actually a subgenus rather than a separate genus.
The proposed division of the Leishmania into Euleishmania and Paraleishmania groups in 2000 emphasized the deep phylogenic distance between parasites, some of which had been named as Leishmania species.[24] The Euleishmania included species currently placed in the subgenera Leishmania, Sauroleishmania, Mundinia and Viannia. The proposed Paraleishmania included species of Endotypanum, Leishmamnia-L. colomubensis, L. herreri, L. hertigiand L. deanei and L. equatorensis. In a recent revision these species were given different generic status.
Four subgenera of Leishmania are now recognised - Leishmania, Sauroleishmania, Viannia and Mundinia (the L. enriettii complex). The genus Endotrypanum and Porcisia belong to the Paraleishmania.
There are four Mundinia species - L. (Mundinia) enriettii, L. (Mundinia) martiniquensis, L. (Mundinia) macropodum, and L. (Mundinia) orientalis, which is found in Thailand.[25]
L. archibaldi's specific status is unsettled but it is closely related to L. donovani.
L. herreri belongs to the genus Endotypanum rather than to Leishmania.
L. donovani and L. infantum are closely related.
The selenoenzyme Seltryp appears to be unique to this order.[26] It has been removed from the subgenus Viannia.
L. deanei and L. hertigi, both of which infect porcupines have been moved to the genus Porcisia.
Classification
Subgenus Leishmania Ross, 1903 sensu Saf'janova, 1982
- Leishmania aethiopica Bray, Ashford & Bray, 1973
- Leishmania amazonensis Lainson & Shaw, 1972 (includes garnhami Scorza et al., 1979)
- Leishmania arabica Peters, Elbihari & Evans, 1986
- Leishmania aristidesi Lainson & Shaw, 1979
- Leishmania donovani (Laveran & Mesnil, 1903)
- Leishmania forattinii Yoshida, Cuba, Pacheco, Cupolillo, Tavares, Machado, Homen & Grimaldi, 1993
- Leishmania gerbilli Wang, Qu & Guan, 1964
- Leishmania infantum Nicolle, 1908 (subspecies chagasi Cunha & Chagas, 1937)
- Leishmania killicki Rioux, Lamotte & Pratlong, 1986
- Leishmania major Yakimoff & Schokhor, 1914
- Leishmania mexicana Biagi, 1953
- Leishmania pifanoi Medina & Romero, 1959
- Leishmania tropica (Wright, 1903)
- Leishmania turanica Strelkova, Shurkhal, Kellina, Eliseev, Evans, Peters, Chapman, Le Blancq & van Eys, 1990
- Leishmania venezeulensis Bonfante-Garrido, 1980
- Leishmania waltoni Shaw, Pratlong & Dedet 2015
Subgenus Mundinia Shaw,Camargo and Teixeira 2016
- Leishmania enriettiiMuniz & Medina, 1948
- Leishmania macropodum Barratt, Kaufer, Peters, Craig, Lawrence, Roberts, Lee, McAuliffe, Stark, Ellis, 2017
- Leishmania martiniquensis Desbois, Pratlong, Quist and Dedet,2014[27]
- Leishmania orientalis Jariyapan, Daroontum, Jaiwong, Chanmol,.Intakhan, Sor-Suwan, Siriyasatien, Somboon, Bates, Bates, 2018
Subgenus Sauroleishmania Ranque, 1973 sensu Saf'janova, 1982
- Leishmania adleri Heisch, 1958
- Leishmania agamae David, 1929
- Leishmania ceramodactyli Adler & Theodor, 1929
- Leishmania gulikae Ovezmukhammedov & Saf'janova, 1987
- Leishmania gymnodactyli † Khodukin & Sofiev, 1940
- Leishmania helioscopi † Chodukin & Sofiev, 1940
- Leishmania hemidactyli Mackie, Gupta & Swaminath, 1923
- Leishmania hoogstraali McMillan, 1965
- Leishmania nicollei Chodukin & Sofieff, 1940
- Leishmania platycephala Telford, 2009
- Leishmania phrynocephali Chodukin & Sofieff, 1940
- Leishmania senegalensis Ranque, 1973
- Leishmania sofieffi ↑ Markov, Lukina & Markova, 1964
- Leishmania tarentolae Wenyon, 1921
- Leishmania zmeevi ↑ Andruchko & Markov 1955
- Leishmania zuckermani Paperna, Boulard, Hering-Hagenbeck & Landau, 2001
↑ Species described as Sauroleishmania. Their development is not like other members of the subgenus and so their taxonomic position is doubtful.
Subgenus Viannia Lainson & Shaw 1987
- Leishmania braziliensis Vianna, 1911
- Leishmania guyanensis Floch, 1954
- Leishmania lainsoni Silveira, Shaw, Braga & Ishikawa, 1987
- Leishmania lindenbergi Silveira, Ishikawa, De Souza & Lainson, 2002
- Leishmania naiffi Lainson & Shaw, 1989
- Leishmania panamensis Lainson & Shaw, 1972
- Leishmania peruviana Velez, 1913
- Leishmania shawi Lainson, Braga & de Souza, 1989
- Leishmania utingensis Braga, Lainson, Ishikawa & Shaw 2003
Related genera
The relationships between these genera and Leishmania is presently unclear but they are closely related.
Genus Endotrypanum
- Endotrypanum colombiensis (Kreutzer, Corredor, Grimaldi, Grogl, Rowton, Young, Morales, McMahon-Pratt, Guzman & Tesh, 1991)
- Endotrypanum equatorensis (Grimaldi, Kreutzer, Hashiguchi, Gomet, Mimory & Tesh, 1992)
- Endotrypanum herreri (Zeledon, Ponce & Murillo, 1979)
- Endotrypanum monterogeii Shaw, 1969
- Endotrypanum schaudinni Mesnil and Brimont, 1908
Genus Novymonas Kostygov and Yurchenko 2016
- Novymonas esmeraldas Votýpka, Kostygov, Maslov and Lukeš 2016
Genus Porcisia Shaw, Camargo and Teixeira 2016
- Porcisia deanei (Lainson & Shaw 1977)
- Porcisia hertigi (Herrer 1971)
Genus Zelonia Shaw, Camargo and Teixeira 2016
- Zelonia australiensis Barratt, Kaufer, Peters, Craig, Lawrence, Roberts, Lee, McAuliffe, Stark, Ellis 2017
- Zelonia costaricensis (Yurchenko, Lukes, Jirku, Zeledon, Maslov 2006)
Biochemistry and cell biology
The biochemistry and cell biology of Leishmania is similar to that of other kinetoplastids. They share the same main morphological features, including a single flagellum which has an invagination, the flagellar pocket, at its base, a kinetoplast, which is found in the single mitochondrion, and a subpelicular array of microtubules, which make up the main part of the cytoskeleton.
Lipophosphoglycan coat
Leishmania possesses a lipophosphoglycan coat over the outside of the cell. Lipophosphoglycan is a trigger for toll-like receptor 2, a signalling receptor involved in triggering an innate immune response in mammals.
The precise structure of lipophosphoglycan varies depending on the species and lifecycle stage of the parasite. The glycan component is particularly variable and different lipophosphoglycan variants can be used as a molecular marker for different lifecycle stages. Lectins, a group of proteins which bind different glycans, are often used to detect these lipophosphoglycan variants. For example, peanut agglutinin binds a particular lipophosphoglycan found on the surface of the infective form of L. major.
Lipophosphoglycan is used by the parasite to promote its survival in the host and the mechanisms by which the parasite does this center around modulating the immune response of the host. This is vital, as the Leishmania parasites live within macrophages and need to prevent the macrophages from killing them. Lipophosphoglycan has a role in resisting the complement system, inhibiting the oxidative burst response, inducing an inflammation response and preventing natural killer T cells recognising that the macrophage is infected with the Leishmania parasite.
| Type | Pathogen | Location |
|---|---|---|
| Cutaneous leishmaniasis (localised and diffuse) infections appear as obvious skin reactions. | The most common is the Oriental Sore (caused by Old World species L. major, L. tropica, and L. aethiopica). In the New World, the most common culprits is L. mexicana. | Cutaneous infections are most common in Afghanistan, Brazil, Iran, Peru, Saudi Arabia and Syria. |
| Mucocutaneous leishmaniasis infections start off as a reaction at the bite, and can go by metastasis into the mucous membrane and become fatal. | L. braziliensis | Mucocutaneous infections are most common in Bolivia, Brazil and Peru. Mucocutaneous infections are also found in Karamay, China Xinjiang Uygur Autonomous Region. |
| Visceral leishmaniasis infections are often recognised by fever, swelling of the liver and spleen, and anemia. They are known by many local names, of which the most common is probably kala azar,[28][29] | Caused exclusively by species of the L. donovani complex (L. donovani, L. infantum syn. L. chagasi).[3] | Found in tropical and subtropical areas of all continents except Australia, visceral infections are most common in Bangladesh, Brazil, India, Nepal, and Sudan.[3] Visceral leishmaniasis also found in part of China, such as Sichuan Province, Gansu Province, and Xinjiang Uygur Autonomous Region. |
Intracellular mechanism of infection
In order to avoid destruction by the immune system and thrive, the Leishmania 'hides' inside its host's cells. This location enables it to avoid the action of the humoral immune response (because the pathogen is safely inside a cell and outside the open bloodstream), and furthermore it may prevent the immune system from destroying its host through nondanger surface signals which discourage apoptosis. The primary cell types Leishmania infiltrates are phagocytotic cells such as neutrophils and macrophages.[30]
Usually, a phagocytotic immune cell like a macrophage will ingest a pathogen within an enclosed endosome and then fill this endosome with enzymes which digest the pathogen. However, in the case of Leishmania, these enzymes have no effect, allowing the parasite to multiply rapidly. This uninhibited growth of parasites eventually overwhelms the host macrophage or other immune cell, causing it to die.[31]
Transmitted by the sandfly, the protozoan parasites of L. major may switch the strategy of the first immune defense from eating/inflammation/killing to eating/no inflammation/no killing of their host phagocyte and corrupt it for their own benefit. They use the willingly phagocytosing polymorphonuclear neutrophil granulocytes (PMNs) rigorously as a tricky hideout, where they proliferate unrecognized from the immune system and enter the long-lived macrophages to establish a "hidden" infection.
Uptake and survival
Upon microbial infection, PMNs move out from the bloodstream through the vessels’ endothelial layer, to the site of the infected tissue (dermal tissue after fly bite). They immediately initiate the first immune response and phagocytize the invader by recognition of foreign and activating surfaces on the parasite. Activated PMN secrete chemokines, IL-8 particularly, to attract further granulocytes and stimulate phagocytosis. Further, L. major increases the secretion of IL-8 by PMNs. This mechanism is observed during infection with other obligate intracellular parasites, as well. For microbes like these, multiple intracellular survival mechanisms exist. Surprisingly, the coinjection of apoptotic and viable pathogens causes by far a more fulminate course of disease than injection of only viable parasites. When the anti-inflammatory signal phosphatidylserine usually found on apoptotic cells, is exposed on the surface of dead parasites, L. major switches off the oxidative burst, thereby preventing killing and degradation of the viable pathogen.
In the case of Leishmania, progeny are not generated in PMNs, but in this way they can survive and persist untangled in the primary site of infection. The promastigote forms also release Leishmania chemotactic factor (LCF) to actively recruit neutrophils, but not other leukocytes, for instance monocytes or NK cells. In addition to that, the production of interferon gamma (IFNγ)-inducible protein 10 (IP10) by PMNs is blocked in attendance of Leishmania, what involves the shut down of inflammatory and protective immune response by NK and Th1 cell recruitment. The pathogens stay viable during phagocytosis since their primary hosts, the PMNs, expose apoptotic cell-associated molecular pattern (ACAMP) signaling "no pathogen".
Persistency and attraction
The lifespan of neutrophil granulocytes is quite short. They circulate in bloodstream for about 6 to 10 hours after leaving bone marrow, whereupon they undergo spontaneous apoptosis. Microbial pathogens have been reported to influence cellular apoptosis by different strategies. Obviously because of the inhibition of caspase3-activation, L. major can induce the delay of neutrophils apoptosis and extend their lifespan for at least 2–3 days. The fact of extended lifespan is very beneficial for the development of infection because the final host cells for these parasites are macrophages, which normally migrate to the sites of infection within two or three days. The pathogens are not dronish; instead they take over the command at the primary site of infection. They induce the production by PMNs of the chemokines MIP-1α and MIP-1β (macrophage inflammatory protein) to recruit macrophages.[32]
An important factor in prolonging infection is the inhibition of the adaptive immune system. This occurs especially during the intercellular phases, when amastigotes search for new macrophages to infect and are more susceptible to immune responses. Nearly all types of phagocytes are targeted.[33] For example, mincle has been shown to be targeted by L. major. Interaction between mincle and a protein released by the parasite results in a weakened immune response in dendritic cells.[34]
Silent phagocytosis theory
To save the integrity of the surrounding tissue from the toxic cell components and proteolytic enzymes contained in neutrophils, the apoptotic PMNs are silently cleared by macrophages. Dying PMNs expose the "eat me"-signal phosphatidylserine which is transferred to the outer leaflet of the plasma membrane during apoptosis. By reason of delayed apoptosis, the parasites that persist in PMNs are taken up into macrophages, employing an absolutely physiological and nonphlogistic process. The strategy of this "silent phagocytosis" has the following advantages for the parasite:
- Taking up apoptotic cells silences macrophage killing activity leading to a survival of the pathogens.
- Pathogens inside of PMNs have no direct contact to the macrophage surface receptors, because they can not see the parasite inside the apoptotic cell. So, the activation of the phagocyte for immune activation does not occur.
However, studies have shown this is unlikely, as the pathogens are seen to leave apoptopic cells and no evidence is known of macrophage uptake by this method.
Molecular biology
An important aspect of the Leishmania protozoan is its glycoconjugate layer of lipophosphoglycan (LPG). This is held together with a phosphoinositide membrane anchor, and has a tripartite structure consisting of a lipid domain, a neutral hexasaccharide, and a phosphorylated galactose-mannose, with a termination in a neutral cap. Not only do these parasites develop postphlebotomus digestion, but it is also thought to be essential to oxidative bursts, thus allowing passage for infection. Characteristics of intracellular digestion include an endosome fusing with a lysosome, releasing acid hydrolases which degrade DNA, RNA, proteins and carbohydrates.
Genomics
The genomes of four Leishmania species (L. major, L. infantum, L. donovani and L. braziliensis) have been sequenced, revealing more than 8300 protein-coding and 900 RNA genes. Almost 40% of protein-coding genes fall into 662 families containing between two and 500 members. Most of the smaller gene families are tandem arrays of one to three genes, while the larger gene families are often dispersed in tandem arrays at different loci throughout the genome. Each of the 35 or 36 chromosomes is organized into a small number of gene clusters of tens-to-hundreds of genes on the same DNA strand. These clusters can be organized in head-to-head (divergent) or tail-to-tail (convergent) fashion, with the latter often separated by tRNA, rRNA and/or snRNA genes. Transcription of protein-coding genes initiates bidirectionally in the divergent strand-switch regions between gene clusters and extends polycistronically through each gene cluster before terminating in the strand-switch region separating convergent clusters. Leishmania telomeres are usually relatively small, consisting of a few different types of repeat sequence. Evidence can be found for recombination between several different groups of telomeres. The L. major and L. infantum genomes contain only about 50 copies of inactive degenerated Ingi/L1Tc-related elements (DIREs), while L. braziliensis also contains several telomere-associated transposable elements and spliced leader-associated retroelements. The Leishmania genomes share a conserved core proteome of about 6200 genes with the related trypanosomatids Trypanosoma brucei and Trypanosoma cruzi, but around 1000 Leishmania-specific genes are known, which are mostly randomly distributed throughout the genome. Relatively few (about 200) species-specific differences in gene content exist between the three sequenced Leishmania genomes, but about 8% of the genes appear to be evolving at different rates between the three species, indicative of different selective pressures that could be related to disease pathology. About 65% of protein-coding genes currently lack functional assignment.[4]
Leishmania species produce several different heat shock proteins. These include Hsp83, a homolog of Hsp90. A regulatory element in the 3' UTR of Hsp83 controls translation of Hsp83 in a temperature-sensitive manner. This region forms a stable RNA structure which melts at higher temperatures.[35]
Sexual reproduction
A microbial pathogen's reproductive system is one of the basic biologic processes that condition the microorganism's ecology and disease spread.[36] In 2009 Akopyants et al.[37] demonstrated that L. major has a sexual cycle, including a meiotic process. Hybrid progeny are formed that have full genomic complements from both parents. Mating only occurs in the sand fly vector, and hybrids can be transmitted to the mammalian host by sand fly bite. In L. braziliensis matings in nature are predominantly between related individuals resulting in extreme inbreeding.[38] The rate of outcrossing between different strains of Leishmania in the sand fly vector depends on the frequency of co-infection. Such outcrossing events appear to be rare in L. major [37] and L. donovani.[39]
L. infantum produces proteins BRCA1 and RAD51 that interact with each other to promote homologous recombinational repair.[40] These proteins play a key role in meiosis. Thus, meiotic events provide the adaptive advantage of efficient recombinational repair of DNA damages even when they do not lead to outcrossing[41]
See also
- Canine leishmaniasis
- List of parasites (human)
References
- ↑ Schnur, L. F (1989). "On the Clinical Manifestations and Parasites of Old World Leishmaniases and Leishmania Tropica Causing Visceral Leishmaniasis". In Hart, D. T. (ed.). Leishmaniasis. pp. 939–43. doi:10.1007/978-1-4613-1575-9_119. ISBN 978-1-4612-8862-6.
- ↑ "leishmania". The Chambers Dictionary (9th ed.). Chambers. 2003. ISBN 0-550-10105-5.
- 1 2 3 Ryan KJ; Ray CG (editors) (2004). Sherris Medical Microbiology (4th ed.). McGraw Hill. pp. 749–54. ISBN 0-8385-8529-9.
{{cite book}}: CS1 maint: multiple names: authors list (link) - 1 2 Myler P; Fasel N (editors) (2008). Leishmania: After The Genome. Caister Academic Press. ISBN 978-1-904455-28-8.
{{cite book}}: CS1 maint: multiple names: authors list (link) - ↑ Ansari MY, Equbal A, Dikhit MR, Mansuri R, Rana S, Ali V, Sahoo GC, Das P (Nov 2015). "Establishment of Correlation between In-Silico &In-Vitro Test Analysis against Leishmania HGPRT to inhibitors". International Journal of Biological Macromolecules. 83: 78–96. doi:10.1016/j.ijbiomac.2015.11.051. PMID 26616453.
- ↑ WHO (2010) Annual report. Geneva
- ↑ Cox, F. E. G (2002). "History of Human Parasitology". Clinical Microbiology Reviews. 15 (4): 595–612. doi:10.1128/CMR.15.4.595-612.2002. PMC 126866. PMID 12364371.
- ↑ Yanik, M.; et al. (2004). "The psychological impact of cutaneous leishmaniasis". Clinical and Experimental Dermatology. 29 (5): 464–467. doi:10.1111/j.1365-2230.2004.01605.x. PMID 15347324. S2CID 11543741.
- ↑ Poinar, G (2008). "Lutzomyia adiketis sp. n. (Diptera: Phlebotomidae), a vector of Paleoleishmania neotropicum sp. n. (Kinetoplastida: Trypanosomatidae) in Dominican amber". Parasit Vectors. 1 (1): 2. doi:10.1186/1756-3305-1-22. PMC 2491605. PMID 18627624.
- ↑ "Morphology and Life Cycle". UCLA. Archived from the original on 13 March 2014. Retrieved 24 January 2014.
- ↑ Pulvertaft, RJ; Hoyle, GF (1960). "Stages in the life-cycle of Leishmania donovani". Transactions of the Royal Society of Tropical Medicine and Hygiene. 54 (2): 191–6. doi:10.1016/0035-9203(60)90057-2. PMID 14435316.
- ↑ Hughes, AL; Piontkivska, H (2003). "Phylogeny of Trypanosomatidae and Bodonidae (Kinetoplastida) based on 18S rRNA: evidence for paraphyly of Trypanosoma and six other genera". Mol Biol Evol. 20 (4): 644–652. doi:10.1093/molbev/msg062. PMID 12679543.
- ↑ Momen H, Cupolillo E (2000). "Speculations on the origin and evolution of the genus Leishmania". Mem. Inst. Oswaldo Cruz. 95 (4): 583–8. doi:10.1590/S0074-02762000000400023. PMID 10904419.
- ↑ Noyes HA, Morrison DA, Chance ML, Ellis JT (2000). "Evidence for a neotropical origin of Leishmania". Mem. Inst. Oswaldo Cruz. 95 (4): 575–8. doi:10.1590/S0074-02762000000400021. PMID 10904417.
- ↑ Kerr SF (2000). "Palaearctic origin of Leishmania". Mem. Inst. Oswaldo Cruz. 95 (1): 75–80. doi:10.1590/S0074-02762000000100011. PMID 10656708.
- ↑ Kuhls, Katrin; Alam, Mohammad Zahangir; Cupolillo, Elisa; Ferreira, Gabriel Eduardo M.; Mauricio, Isabel L.; Oddone, Rolando; Feliciangeli, M. Dora; Wirth, Thierry; Miles, Michael A.; Schönian, Gabriele; Kamhawi, Shaden (7 June 2011). "Comparative Microsatellite Typing of New World Leishmania infantum Reveals Low Heterogeneity among Populations and Its Recent Old World Origin". PLOS Neglected Tropical Diseases. 5 (6): e1155. doi:10.1371/journal.pntd.0001155. PMC 3110170. PMID 21666787.
- ↑ Duprey, Z. H.; Steurer, F. J.; Rooney, J. A.; Kirchhoff, L. V.; Jackson, J. E.; Rowton, E. D.; Schantz, P. M. (2006). "Canine Visceral Leishmaniasis, United States and Canada, 2000–2003". Emerging Infectious Diseases. 12 (3): 440–446. doi:10.3201/eid1203.050811. PMC 3291440. PMID 16704782.
- ↑ Noyes, HA; Arana, BA; Chance, ML; Maingon, R (1997). "The Leishmania hertigi (Kinetoplastida; Trypanosomatidae) complex and the lizard Leishmania: their classification and evidence for a neotropical origin of the Leishmania-Endotrypanum clade". J Eukaryot Microbiol. 44 (5): 511–557. doi:10.1111/j.1550-7408.1997.tb05732.x. PMID 9304821. S2CID 27460253.
- ↑ Harkins, Kelly M; Schwartz, Rachel S; Cartwright, Reed A; Stone, Anne C (2016). "Phylogenomic reconstruction supports supercontinent origins for Leishmania". Infection, Genetics and Evolution. 38: 101–9. doi:10.1016/j.meegid.2015.11.030. PMID 26708057.
- ↑ Croan, David G; Morrison, David A; Ellis, John T (1997). "Evolution of the genus Leishmania revealed by comparison of DNA and RNA polymerase gene sequences". Molecular and Biochemical Parasitology. 89 (2): 149–59. doi:10.1016/S0166-6851(97)00111-4. PMID 9364962.
- ↑ Jennings, Y. L.; de Souza, A. A. A.; Ishikawa, E. A.; Shaw, J.; Lainson, R.; Silveira, F. (2014). "Phenotypic characterization of Leishmania spp. causing cutaneous leishmaniasis in the lower Amazon region, western Pará state, Brazil, reveals a putative hybrid parasite, Leishmania (Viannia) guyanensis × Leishmania (Viannia) shawi shawi". Parasite. 21: 39. doi:10.1051/parasite/2014039. PMC 4118625. PMID 25083790.
- ↑ Espinosa, O.A.; Serrano, M.G.; Camargo, E.P.; M.M.G., Teixeira; J.J., Shaw (2016). "An appraisal of the taxonomy and nomenclature of trypanosomatids presently classified as Leishmania' 'and' 'Endotrypanum". Parasitology. 145 (4): 430–442. doi:10.1017/S0031182016002092. PMID 27976601. S2CID 41544524.
- ↑ Franco, AM; Grimaldi, G Jr (1999). "Characterization of Endotrypanum (Kinetoplastida: Trypanosomatidae), a unique parasite infecting the neotropical tree sloths (Edentata)". Mem Inst Oswaldo Cruz. 94 (2): 261–268. doi:10.1590/s0074-02761999000200026. PMID 10224540.
- ↑ Momen, H; Cupolillo, E (2000). "Speculations on the origin and evolution of the genus Leishmania". Mem Inst Oswaldo Cruz. 95 (4): 583–588. doi:10.1590/s0074-02762000000400023. PMID 10904419.
- ↑ Jariyapan, N; Daroontum, T; Jaiwong, K; Chanmol, W; Intakhan, N; Sor-Suwan, S; Siriyasatien, P; Somboon, P; Bates, MD; Bates, PA (2018). "Leishmania (Mundinia) orientalis n. sp. (Trypanosomatidae), a parasite from Thailand responsible for localised cutaneous leishmaniasis". Parasit Vectors. 11 (1): 351. doi:10.1186/s13071-018-2908-3. PMC 6006788. PMID 29914526.
- ↑ Bonilla, Mariana; Krull, Erika; Irigoín, Florencia; Salinas, Gustavo; Comini, Marcelo A. (2016). "Selenoproteins of African trypanosomes are dispensable for parasite survival in a mammalian host". Molecular and Biochemical Parasitology. 206 (1–2): 13–19. doi:10.1016/j.molbiopara.2016.03.002. PMID 26975431.
- ↑ Desbois, Nicole; Pratlong, Francine; Quist, Danièle; Dedet, Jean-Pierre (2014). "Leishmania (Leishmania) martiniquensis n. sp. (Kinetoplastida: Trypanosomatidae), description of the parasite responsible for cutaneous leishmaniasis in Martinique Island (French West Indies)". Parasite. 21: 12. doi:10.1051/parasite/2014011. ISSN 1776-1042. PMC 3952653. PMID 24626346.

- ↑ Visceral leishmniasis: The disease Archived 2005-04-28 at the Wayback Machine
- ↑ kala-azar Archived 2009-02-10 at the Wayback Machine. The American Heritage Dictionary of the English Language
- ↑ Vannier-Santos, MA; Martiny A; de Souza W. (August 2002). "Cell biology of Leishmania spp.: invading and evading". Current Pharmaceutical Design. 8 (4): 297–318. doi:10.2174/1381612023396230. PMID 11860368.
- ↑ Paul, William E. (September 1993). "Infectious Diseases and the Immune System". Scientific American. 269 (3): 94–95. Bibcode:1993SciAm.269c..90P. doi:10.1038/scientificamerican0993-90. PMID 8211095.
- ↑ Laskay T, et al. (2003). "Neutrophil granulocytes – Trojan horses for Leishmania major and other intracellular microbes?". Trends in Microbiology. 11 (5): 210–4. doi:10.1016/S0966-842X(03)00075-1. PMID 12781523.
- ↑ Martínez-López M, et al. (2018). "Leishmania Hijacks Myeloid Cells for Immune Escape". Frontiers in Microbiology. 9: 883. doi:10.3389/fmicb.2018.00883. PMC 5949370. PMID 29867798.
- ↑ Iborra S, et al. (2016). "Leishmania uses Mincle to target an inhibitory ITAM signaling pathway in dendritic cells that dampens adaptive immunity to infection". Immunity. 45 (4): 788–801. doi:10.1016/j.immuni.2016.09.012. PMC 5074365. PMID 27742545.
- ↑ David, M; Gabdank, I; Ben-David, M; Zilka, A; Orr, I; Barash, D; Shapira, M (February 2010). "Preferential translation of Hsp83 in Leishmania requires a thermosensitive polypyrimidine-rich element in the 3' UTR and involves scanning of the 5' UTR". RNA. 16 (2): 364–74. doi:10.1261/rna.1874710. PMC 2811665. PMID 20040590.
- ↑ Rougeron V, De Meeûs T, Kako Ouraga S, Hide M, Bañuls AL (2010). ""Everything you always wanted to know about sex (but were afraid to ask)" in Leishmania after two decades of laboratory and field analyses". PLOS Pathog. 6 (8): e1001004. doi:10.1371/journal.ppat.1001004. PMC 2924324. PMID 20808896.
- 1 2 Akopyants NS, Kimblin N, Secundino N, Patrick R, Peters N, Lawyer P, Dobson DE, Beverley SM, Sacks DL (April 2009). "Demonstration of genetic exchange during cyclical development of Leishmania in the sand fly vector". Science. 324 (5924): 265–8. Bibcode:2009Sci...324..265A. doi:10.1126/science.1169464. PMC 2729066. PMID 19359589.
- ↑ Rougeron V, De Meeûs T, Hide M, Waleckx E, Bermudez H, Arevalo J, Llanos-Cuentas A, Dujardin JC, De Doncker S, Le Ray D, Ayala FJ, Bañuls AL (June 2009). "Extreme inbreeding in Leishmania braziliensis". Proc. Natl. Acad. Sci. U.S.A. 106 (25): 10224–9. Bibcode:2009PNAS..10610224R. doi:10.1073/pnas.0904420106. PMC 2700931. PMID 19497885.
- ↑ Rogers MB, Downing T, Smith BA, Imamura H, Sanders M, Svobodova M, Volf P, Berriman M, Cotton JA, Smith DF (January 2014). "Genomic confirmation of hybridisation and recent inbreeding in a vector-isolated Leishmania population". PLOS Genet. 10 (1): e1004092. doi:10.1371/journal.pgen.1004092. PMC 3894156. PMID 24453988.
- ↑ Genois MM, Mukherjee A, Ubeda JM, Buisson R, Paquet E, Roy G, Plourde M, Coulombe Y, Ouellette M, Masson JY (August 2012). "Interactions between BRCA2 and RAD51 for promoting homologous recombination in Leishmania infantum". Nucleic Acids Res. 40 (14): 6570–84. doi:10.1093/nar/gks306. PMC 3413117. PMID 22505581.
- ↑ Bernstein H, Byerly HC, Hopf FA, Michod RE. Genetic damage, mutation, and the evolution of sex. Science. 1985 Sep 20;229(4719):1277-81. doi: 10.1126/science.3898363. PMID: 3898363
Further reading
- Van Zandbergen, G; Bollinger, A; Wenzel, A; Kamhawi, S; Voll, R; Klinger, M; Muller, A; Holscher, C; Herrmann, M; Sacks, D; Solbach, W; Laskay, T (2006). "Leishmania disease development depends on the presence of apoptotic promastigotes in the virulent inoculum". Proceedings of the National Academy of Sciences. 103 (37): 13837–42. Bibcode:2006PNAS..10313837V. doi:10.1073/pnas.0600843103. PMC 1564231. PMID 16945916.
- Shaw, Jeffrey Jon (1969). The Haemoflagellates of Sloths Archived 2020-03-18 at the Wayback Machine. H. K. Lewis: London.
- Ansari MY, Dikhit MR, Sahoo GC, Das P (2012). "Comparative modeling of HGPRT enzyme of L. donovani and binding affinities of different analogs of GMP". Int J Biol Macromol. 50 (3): 637–49. doi:10.1016/j.ijbiomac.2012.01.010. PMID 22327112.
External links
- The International Leishmania Network (ILN) Archived 2017-09-22 at the Wayback Machine has basic information on the disease and links to many aspects of the disease and its vector.
- A discussion list (Leish-L) Archived 2021-05-11 at the Wayback Machine is also available with over 600 subscribers to the list, ranging from molecular biologists to public health workers, from many countries both inside and outside endemic regions. Comments and questions are welcomed.
- KBD: Kinetoplastid Biology and Disease Archived 2016-01-10 at the Wayback Machine, is a website devoted to leishmaniasis, sleeping sickness and Chagas disease (American trypanosomiasis). It contains free access to full text peer-reviewed articles on these subjects. The site contains many articles relating to the unique kinetoplastid organelle and genetic material therein.
- Sexual reproduction in leishmania parasites, short review of a "science"-paper Archived 2016-03-04 at the Wayback Machine
- Drug Search for Leishmaniasis Archived 2021-07-16 at the Wayback Machine World Community Grid