Marginal-zone B cell
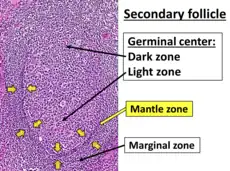
Marginal-zone B cells (MZ B cells) are noncirculating mature B cells that in humans segregate anatomically into the marginal zone (MZ) of the spleen[1] and certain other types of lymphoid tissue.[2] The MZ B cells within this region typically express low-affinity polyreactive B-cell receptors (BCR), high levels of IgM, Toll-like receptors (TLRs), CD21, CD1, CD9, CD27 with low to negligible levels of secreted-IgD, CD23, CD5, and CD11b that help to distinguish them phenotypically from follicular (FO) B cells and B1 B cells.[2][3]
MZ B cells are innate-like B cells specialized to mount rapid T-independent, but also T-dependent responses against blood-borne pathogens.[4] They are also known to be the main producers of IgM antibodies in humans.[5]
Development and differentiation
The spleen's marginal zone contains multiple subtypes of macrophages and dendritic cells interlaced with the MZ B cells; it is not fully formed until 2 to 3 weeks after birth in rodents and 1 to 2 years in humans.[6] In humans, but not rodents, marginal zone B cells are also located in the inner wall of the subcapsular sinus of lymph nodes, the epithelium of tonsillar crypts, and the sub-epithelial area of mucosa-associated lymphoid tissues including the sub-epithelial dome of intestinal Peyer's patches.[2] Human MZ B cells are also present in peripheral blood, suggesting that they recirculate.[7] However, in mice they seem to be noncirculating and only limited to follicular shuttling.[2]
In rodents, MZ B cells are recognized as IgMhighIgDlowCD21highCD23low population of B cells. They are furthermore distinguished by the expression of CD9[3] and CD27 (in humans).[2] In mice, MZ B cells characteristically express high levels of CD1d, which is an MHC class I-like molecule involved in the presentation of lipid molecules to NKT cells.[8] Unlike FO B cells, MZ B cells express polyreactive BCRs that bind to multiple microbial molecular patterns. Additionally, they express high levels of TLRs.[2]
In specimens where the tyrosine kinase for Pyk-2 has been knocked-out, marginal zone B-cells will fail to develop while B-1 cells will still be present. MZ B-cells are the only B-cells dependent on NOTCH2 signaling for proliferation.[9]
Activation and function
Similar to B1 B cells, MZ B cells can be rapidly recruited into the early adaptive immune responses in a T cell-independent manner.[9] The MZ B cells are especially well-positioned as the first line of defense against systemic blood-borne antigens that enter the circulation and become trapped in the spleen.[10] While large blood-borne antigens are captured by dendritic cells, circulating granulocytes or MZ macrophages, smaller blood-borne antigens may directly interact with MZ B cells situated on the exterior of the marginal sinus.[2][4] MZ B cells shuttle between the blood-filled marginal zone for antigen collection and the follicle for antigen delivery to follicular dendritic cells. In mice, it has been shown that these cells shear flow via the LFA-1 integrin ligand ICAM-1 and adhere or migrate down the flow via the VLA-4 integrin ligand VCAM-1. While CXCR5/CXCL13 signaling is required for MZ B cells to enter the follicle, Sphingosine-1-phosphate signaling is required for them to exit from the follicle.[11]
MZ B cells respond to a wide spectrum of T-independent, but also T-dependent antigens. It is believed that MZ B cells are especially reactive to microbial polysaccharide antigens of encapsulated bacteria such as Streptococcus pneumoniae, Haemophilus influenzae and Neisseria meningitidis. TLRs often activate MZ B cells after recognizing microbial molecular structures in cooperation with the BCR.[7] These innate-like B cells provide a rapid first line of defense against blood-borne pathogens and produce low-affinity antibodies of wide specificity before the induction of T-cell-dependent high-affinity antibody responses. Therefore MZ B cells may play an important role in the prevention of sepsis.[8] MZ B cells also display a lower activation threshold than their FO B cell counterparts, with a heightened propensity for plasma cell differentiation that contributes further to the accelerated primary antibody response.[2][12] They have been acknowledged as the main producers of IgM antibodies in humans.[5]
They are important for antibody-response toward invading pathogens and maintaining homeostasis via opsonization of dead cells and cellular debris.[5] Moreover, MZ B cells are potent antigen-presenting cells, that are able to activate CD4+ T cells more effectively than FO B cells due to their elevated expression levels of MHC class II, CD80 and CD86 molecules.[2][7]
Deficiencies of MZ B cells are associated with a higher risk of pneumococcal infection, meningitis and insufficient antibody response to capsular polysaccharides.[2][4]
Memory
In humans the splenic marginal zone B cells have evidence of somatic hypermutation in their immunoglobulin genes, indicating that they have been generated through a germinal centre reaction to become memory cells. While naive MZ B cells produce low-affinity IgM antibodies, memory MZ B cells express high-affinity Ig molecules. Besides unswitched cells (IgM+), class-switched B cells can be found in the human and rodent marginal zone (IgG+ and IgA+). In humans, MZ B cells express CD27, which is a member of the TNF-receptor family expressed by human memory B cells.[8]
Role in autoimmune diseases
Many of MZ B cell-receptors are self-reactive, which may be a factor that contributes to their expansion in some autoimmune diseases. On the other hand, aiding in the clearance of self-antigens is considered an important mechanism to prevent the development of autoimmune diseases. The role of expanded self-reactive MZ B cells has been observed on mice models of lupus, diabetes and arthritis.[7] However, their levels in human vasculitis are reduced.[5]
Role in tumors
Marginal zone B cells are the malignant cells in marginal zone lymphomas, a heterogeneous group of generally indolent lymphomas.[13]
References
- ↑ Martin F, Kearney JF (2002). "Marginal-zone B cells". Nat Rev Immunol. 2 (5): 323–335. doi:10.1038/nri799. PMID 12033738. S2CID 22573840.
- 1 2 3 4 5 6 7 8 9 10 Cerutti A, Cols M, Puga I (February 2013). "Marginal zone B cells: virtues of innate-like antibody-producing lymphocytes". Nature Reviews. Immunology. 13 (2): 118–32. doi:10.1038/nri3383. PMC 3652659. PMID 23348416.
- 1 2 Won, Woong-Jai; Kearney, John F. (2002-06-01). "CD9 is a unique marker for marginal zone B cells, B1 cells, and plasma cells in mice". Journal of Immunology. 168 (11): 5605–5611. doi:10.4049/jimmunol.168.11.5605. ISSN 0022-1767. PMID 12023357.
- 1 2 3 Turner, Vivian M.; Mabbott, Neil A. (July 2017). "Ageing adversely affects the migration and function of marginal zone B cells". Immunology. 151 (3): 349–362. doi:10.1111/imm.12737. ISSN 1365-2567. PMC 5461100. PMID 28369800.
- 1 2 3 4 Appelgren, Daniel; Eriksson, Per; Ernerudh, Jan; Segelmark, Mårten (2018-10-02). "Marginal-Zone B-Cells Are Main Producers of IgM in Humans, and Are Reduced in Patients With Autoimmune Vasculitis". Frontiers in Immunology. 9: 2242. doi:10.3389/fimmu.2018.02242. ISSN 1664-3224. PMC 6190848. PMID 30356862.
- ↑ MacLennan IC, Bazin H, Chassoux D; et al. (1985). "Comparative Analysis of the Development of B Cells in Marginal Zones and Follicles". Microenvironments in the Lymphoid System. Advances in Experimental Medicine and Biology. Vol. 186. pp. 139–144. doi:10.1007/978-1-4613-2463-8_17. ISBN 978-1-4612-9495-5. PMID 3876699.
{{cite book}}: CS1 maint: multiple names: authors list (link) - 1 2 3 4 Palm, Anna-Karin E.; Friedrich, Heike C.; Kleinau, Sandra (2016-06-09). "Nodal marginal zone B cells in mice: a novel subset with dormant self-reactivity". Scientific Reports. 6 (1): 27687. Bibcode:2016NatSR...627687P. doi:10.1038/srep27687. ISSN 2045-2322. PMC 4899733. PMID 27277419.
- 1 2 3 Hendricks, Jacobus; Bos, Nicolaas A.; Kroese, Frans G.M. (2018). "Heterogeneity of Memory Marginal Zone B Cells". Critical Reviews in Immunology. 38 (2): 145–158. doi:10.1615/CritRevImmunol.2018024985. ISSN 1040-8401. PMC 5989013. PMID 29887727.
- 1 2 Hardy, Richard (2008). "Chapter 7: B Lymphocyte Development and Biology". In Paul, William (ed.). Fundamental Immunology (Book) (6th ed.). Philadelphia: Lippincott Williams & Wilkins. pp. 237–269. ISBN 978-0-7817-6519-0.
- ↑ Balazs M, Martin F, Zhou T; et al. (2002). "Blood dendritic cells interact with splenic marginal zone B cells to initiate T-independent immune responses". Immunity. 17 (3): 341–352. doi:10.1016/s1074-7613(02)00389-8. PMID 12354386.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ↑ Tedford, Kerry; Steiner, Michael; Koshutin, Stanislav; Richter, Karin; Tech, Laura; Eggers, Yannik; Jansing, Inga; Schilling, Kerstin; Hauser, Anja Erika; Korthals, Mark; Fischer, Klaus-Dieter (2017-12-22). "The opposing forces of shear flow and sphingosine-1-phosphate control marginal zone B cell shuttling". Nature Communications. 8 (1): 2261. Bibcode:2017NatCo...8.2261T. doi:10.1038/s41467-017-02482-4. ISSN 2041-1723. PMC 5741619. PMID 29273735.
- ↑ Lopes-Carvalho T, Foote J, Kearney JF (2005). "Marginal zone B cells in lymphocyte activation and regulation". Curr Opin Immunol. 17 (3): 244–250. doi:10.1016/j.coi.2005.04.009. PMID 15886113.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ↑ Bron D, Meuleman N (September 2019). "Marginal zone lymphomas: second most common lymphomas in older patients". Current Opinion in Oncology. 31 (5): 386–393. doi:10.1097/CCO.0000000000000554. PMID 31246587. S2CID 195765608.