تأشيب جسمي
التأشيب الجسمي (بالإنجليزية: V(D)J recombination) هي آلية فريدة من نوعها من إعادة التركيب الجيني الذي يحدث فقط في تطوير الخلايا الليمفاوية من خلال المراحل النضج الأولى للخلايا البائية والخلايا التائية.[1][2][3] هذه العملية تنتج جعبة متنوعة للغاية من الأجسام المضادة ومستقبلات الخلايا التائية التي توجد في الخلايا البائية والخلايا التائية على التوالي. هذه العملية هي السمة المميزة للنظام المناعة التكيفية وكان تطورها حدثا رئيسيا في تطور الفقاريات الفكية.
هي آلية إعادة التركيب الجسدي التي تحدث فقط في تطوير الخلايا الليمفاوية خلال المراحل المبكرة من نضوج الخلايا التائية والبائية.ينتج عنه ذخيرة متنوعة للغاية من الأجسام المضادة / الغلوبولين المناعي ومستقبلات الخلايا التائية (TCRs) الموجودة في الخلايا البائية والخلايا التائية، على التوالي.هذه العملية هي السمة المميزة لجهاز المناعة التكيفي.[بحاجة لمصدر]
يحدث إعادة التركيب V (D) J في الثدييات في الأعضاء اللمفاوية الأولية؛ (نخاع العظم للخلايا البائية والغدة الصعترية للخلايا التائية) وبطريقة عشوائية تقريبًا يعيد ترتيب المتغير (V)، وينضم (J)، وفي بعض الحالات، التنوع (D) شرائح الجين.تؤدي العملية في النهاية إلى تسلسلات جديدة للأحماض الأمينية في مناطق ارتباط المستضد من الغلوبولين المناعي و TCRs التي تسمح بالتعرف على المستضدات من جميع مسببات الأمراض تقريبًا بما في ذلك البكتيريا والفيروسات والطفيليات والديدان بالإضافة إلى «الخلايا الذاتية المتغيرة» كما شوهد في السرطان.يمكن أن يكون التعرف أيضًا حساسيًا بطبيعته (مثل حبوب اللقاح أو مسببات الحساسية الأخرى) أو قد يتطابق مع أنسجة المضيف ويؤدي إلى المناعة الذاتية.
في عام 1987، حصل سوسومو تونيغاوا على جائزة نوبل في علم وظائف الأعضاء أو الطب «لاكتشافه المبدأ الجيني لتوليد تنوع الأجسام المضادة».
خلفية
تتكون جزيئات الأجسام المضادة البشرية (بما في ذلك مستقبلات الخلايا البائية) من سلاسل ثقيلة وخفيفة، تحتوي كل منها على مناطق ثابتة (C) ومتغيرة (V)، مشفرة وراثيًا في ثلاثة مواضع جينية:[بحاجة لمصدر]
- الموضع الثقيل للجلوبيولين المناعي (IGH @) على الكروموسوم 14 ، والذي يحتوي على المقاطع الجينية للسلسلة الثقيلة للجلوبيولين المناعي.
- موضع الغلوبولين المناعي (κ) (IGK) على الكروموسوم 2 ، الذي يحتوي على مقاطع الجينات لجزء من السلسلة الخفيفة للغلوبولين المناعي.
- موضع الجلوبيولين المناعي لامدا (λ) (IGL) على الكروموسوم 22 ، الذي يحتوي على المقاطع الجينية لبقية سلسلة الغلوبولين المناعي الخفيفة.
تحتوي كل سلسلة ثقيلة أو جين سلسلة خفيفة على نسخ متعددة من ثلاثة أنواع مختلفة من المقاطع الجينية للمناطق المتغيرة من بروتينات الجسم المضاد.على سبيل المثال، تحتوي منطقة السلسلة الثقيلة للجلوبيولين المناعي البشري على جزأين جيني ثابت (C μ و Cδ) و 44 مقطعًا جينيًا متغيرًا (V)، بالإضافة إلى 27 مقطعًا جينيًا متنوعًا (D) و 6 مقاطع جينية متصلة (J).تمتلك السلاسل الخفيفة أيضًا مقطعين جيني ثابت (Cλ و Cκ) والعديد من مقاطع الجين V و J ، لكن لا تحتوي على مقاطع D جينية.تؤدي إعادة ترتيب الحمض النووي إلى انتقال نسخة واحدة من كل نوع من قطع الجينات إلى أي خلية لمفاوية معينة، مما يؤدي إلى إنتاج ذخيرة ضخمة من الأجسام المضادة؛ ما يقرب من 3 × 10^11 مجموعات ممكنة، على الرغم من إزالة بعضها بسبب التفاعل الذاتي.
تتكون معظم مستقبلات الخلايا التائية من سلسلة ألفا متغيرة وسلسلة بيتا.تتشابه جينات مستقبلات الخلايا التائية مع جينات الغلوبولين المناعي من حيث أنها تحتوي أيضًا على أجزاء متعددة من الجينات V و D و J في سلاسل بيتا الخاصة بها (وشرائح الجين V و J في سلاسل ألفا الخاصة بها) والتي يتم إعادة ترتيبها أثناء تطور الخلايا الليمفاوية إلى تزويد تلك الخلية بمستقبل فريد للمستضد.مستقبل الخلايا التائية بهذا المعنى هو المكافئ التوبولوجي لجزء ربط مستضد من الجسم المضاد، وكلاهما جزء من عائلة الغلوبولين المناعي.
يتم منع استجابة المناعة الذاتية عن طريق القضاء على الخلايا التي تتفاعل مع نفسها.يحدث هذا في الغدة الصعترية عن طريق اختبار الخلية ضد مجموعة من المستضدات الذاتية التي يتم التعبير عنها من خلال وظيفة منظم المناعة الذاتية (AIRE).يحتوي موضع السلسلة الخفيفة من الجلوبيولين المناعي لامدا على جينات مشفرة للبروتين يمكن فقدها مع إعادة ترتيبه.يعتمد هذا على آلية فسيولوجية وليس من العوامل الممرضة لسرطان الدم أو الأورام اللمفاوية.تستمر الخلية إذا قامت بإنشاء منتج ناجح لا يتفاعل مع نفسه، وإلا يتم تقليمها عن طريق موت الخلايا المبرمج.
مناعيات
سلسلة ثقيلة
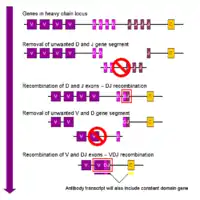
في الخلية البائية النامية، يكون حدث إعادة التركيب الأول بين جزء واحد من الجين D و J من موضع السلسلة الثقيلة.يتم حذف أي DNA بين هذين الجزأين الجينيين.يتبع إعادة التركيب D-J هذا انضمام جزء واحد من الجين V ، من منطقة المنبع لمجمع DJ الذي تم تشكيله حديثا، مما يشكل مقطعا جينيا معاد تركيبه VDJ.يتم الآن حذف جميع أجزاء الجينات الأخرى بين مقاطع V و D من جينوم الخلية.يتم إنشاء النسخة الأولية (الحمض النووي الريبي غير المقسم) تحتوي على منطقة VDJ من السلسلة الثقيلة وكلا من سلاسل مو ودلتا الثابتة (C μ و Cδ).(على سبيل المثال، يحتوي النص الأساسي على المقاطع: V-D-J-Cμ-Cδ).تتم معالجة RNA الأساسي لإضافة ذيل متعدد الأدينيلات (poly-A) بعد سلسلة C ولإزالة التسلسل بين مقطع VDJ وهذا الجزء الجيني الثابت.تؤدي ترجمة هذا الرنا المرسال إلى إنتاج بروتين IgM ثقيل السلسلة.
السلسلة خفيفة
تعيد سلاسل كابا (κ) و (λ) لامدا من مواضع السلسلة الخفيفة للجلوبيولين المناعي ترتيبًا متشابهًا للغاية، باستثناء أن السلاسل الخفيفة تفتقر إلى المقطع D.بعبارة أخرى، تتضمن الخطوة الأولى لإعادة تركيب السلاسل الخفيفة ربط السلاسل V و J لإعطاء مركب VJ قبل إضافة جين السلسلة الثابتة أثناء النسخ الأولي.تؤدي ترجمة الرنا المرسال المقسم إما لسلسلة كابا أو لامدا إلى تكوين بروتين Ig κ أو السلسلة الخفيفة Ig λ .
ينتج عن تجميع السلسلة الثقيلة Ig μ وإحدى السلاسل الخفيفة تكوين شكل مرتبط بالغشاء من الجلوبيولين المناعي IgM الذي يتم التعبير عنه على سطح الخلية البائية غير الناضجة.
مستقبل الخلية التائية
أثناء تطور الخلايا التوتية، تخضع سلاسل مستقبلات الخلايا التائية (TCR) بشكل أساسي لنفس تسلسل أحداث إعادة التركيب المطلوبة كما هو موصوف بالنسبة للجلوبيولينات المناعية. تحدث إعادة التركيب من D إلى J أولاً في سلسلة-بيتا من TCR.يمكن أن تتضمن هذه العملية إما انضمام مقطع الجين Dβ1 إلى واحد من ستة مقاطع Jβ1 أو انضمام جزء الجين Dβ2 إلى واحد من ستة مقاطع Jβ2.يتبع إعادة تركيب DJ (على النحو الوارد أعلاه) مع إعادة ترتيب Vβ-إلى-DβJβ.يتم حذف جميع أجزاء الجينات بين مقاطع الجين Vβ-Dβ-Jβ في المجمع المُشكل حديثًا ويتم تصنيع النسخة الأولية التي تتضمن جين المجال الثابت (Vβ-Dβ-Jβ-كربون ألفا).يقوم نسخ mRNA بتقسيم أي تسلسل متداخل ويسمح بترجمة البروتين كامل الطول لسلسلة TCR β.
يتبع إعادة ترتيب سلسلة ألفا (α) من TCR إعادة ترتيب السلسلة-بيتا، ويشبه إعادة ترتيب V إلى J الموصوف لسلاسل Ig الخفيفة (انظر أعلاه).ينتج عن تجميع السلاسل β- و α- تكوين αβ-TCR الذي يتم التعبير عنه في غالبية الخلايا التائية.
الآلية
الإنزيمات والمكونات الرئيسية
تتم عملية إعادة التركيب V (D) J بواسطة VDJ ريكومبيناز، وهي مجموعة متنوعة من الإنزيمات.الإنزيمات الرئيسية المعنية هي إعادة التركيب الجيني المنشط 1 و 2 (RAG)، وناقل ديوكسينوكليوتيديل الطرفي (TdT)، وارتميس نوكلياز، وهو عضو في مسار الانضمام غير المتماثل موجود في كل مكان (NHEJ) لإصلاح الحمض النووي.من المعروف أن العديد من الإنزيمات الأخرى تشارك في العملية وتشمل بروتين كيناز المعتمد على الحمض النووي (DNA-PK)، والبروتين التكميلي 4 (XRCC4) لإصلاح الأشعة السينية، و DNA ligase IV ، وعامل الانضمام غير المتماثل 1 (NHEJ1)؛ المعروف أيضًا باسم سرنونوس أو بارالوغ [XLF] مثل عامل XRCC-4، المكتشف مؤخرًا لـ XRCC4 و (XLF (PAXX ، وبوليميرات الحمض النووي λ وμ.بعض الإنزيمات المعنية خاصة بالخلايا الليمفاوية (على سبيل المثال، RAG ، TdT)، بينما يوجد البعض الآخر في أنواع الخلايا الأخرى وحتى في كل مكان (على سبيل المثال، مكونات NHEJ).
للحفاظ على خصوصية إعادة التركيب، يتعرف V (D) J ريكومبيناز على تسلسل إشارة إعادة التركيب (RSSs) الذي يحيط بالمتغير (V) والتنوع (D) والانضمام إلى (J) مقاطع الجينات.تتكون RSS من ثلاثة عناصر: هيبتامر من سبعة نيوكليوتيدات محفوظة، ومنطقة مباعدة من 12 أو 23 زوجًا قاعديًا في الطول، ونونامير من تسعة نيوكليوتيدات محفوظة.في حين أن غالبية RSSs تختلف في التسلسل، فإن متواليات هيبتامير ونونامير المتفق عليها هي CACAGTG و ACAAAAACC ، على التوالي؛ وعلى الرغم من عدم حفظ تسلسل منطقة المباعد بشكل جيد، إلا أن الطول محفوظ بدرجة عالية.يتوافق طول منطقة المباعد مع واحد تقريبًا (12 زوجًا أساسيًا) أو منعطفين (23 زوجًا أساسيًا) من حلزون الحمض النووي.باتباع ما يعرف بقاعدة 12/23، عادة ما تكون المقاطع الجينية التي سيتم إعادة تجميعها مجاورة لـ RSSs ذات أطوال مباعدة مختلفة (على سبيل المثال، يحتوي أحدهما على "12RSS" والآخر به "23RSS").هذه ميزة مهمة في تنظيم إعادة التركيب V (D) J.
العملية
يبدأ إعادة التركيب V (D) J عندما يربط V (D) J ريكومبيناز (RAG1 من خلال نشاط) RSS الذي يحيط بمقطع جين الترميز (V أو D أو J) وينشئ شقًا أحادي السلسلة في الحمض النووي بين الأول قاعدة RSS (قبل هبتامير مباشرة) وقطاع الترميز.هذا محايد بشكل أساسي (لا حاجة للتحلل المائي ATP) وينتج عنه تكوين مجموعة هيدروكسيل حرة 3 'ومجموعة 5' فوسفات على نفس الخيط.يتم وضع مجموعة الهيدروكسيل التفاعلية بواسطة ريكومبيناز لمهاجمة رابطة فوسفات ثنائي الاستر من الخيط المعاكس، وتشكيل طرفي DNA: دبوس شعر (حلقة جذعية) على مقطع الترميز ونهاية حادة على قطعة الإشارة.النموذج الحالي هو أن خدش الحمض النووي وتكوين دبوس الشعر يحدث على كلا الخيطين في وقت واحد (أو تقريبًا) في مجمع يُعرف باسم مركز إعادة التركيب.
يتم ربط نهايات الإشارة غير الحادة معًا لتشكيل قطعة دائرية من الحمض النووي تحتوي على جميع التسلسلات المتداخلة بين مقاطع التشفير المعروفة باسم مفصل الإشارة (على الرغم من كونها دائرية بطبيعتها، لا ينبغي الخلط بين هذا وبين البلازميد).بينما كان يُعتقد في الأصل أنه ضاع خلال الانقسامات الخلوية المتتالية، إلا أن هناك دليلًا على أن مفاصل الإشارة قد تدخل مرة أخرى في الجينوم وتؤدي إلى أمراض عن طريق تنشيط الجينات الورمية أو مقاطعة وظيفة الجين الكابت للورم.
تتم معالجة نهايات التشفير بشكل أكبر قبل ربطها من خلال العديد من الأحداث التي تؤدي في النهاية إلى التنوع الوصلي.تبدأ المعالجة عندما يرتبط DNA-PK بكل نهاية DNA مكسورة ويقوم بتجنيد العديد من البروتينات الأخرى بما في ذلك أرتيميس و XRCC4 و DNA ligase IV وسيرنونوس والعديد من بوليمرات الحمض النووي.يشكل DNA-PK مركبًا يؤدي إلى الفسفرة الذاتية، مما يؤدي إلى تنشيط أرتيميس.يتم فتح دبابيس الشعر الطرفية للترميز بواسطة نشاط أرتيميس.إذا تم فتحها في المركز، سينتج عن ذلك نهاية غير حادة للحمض النووي؛ ومع ذلك، في كثير من الحالات، يكون الفتح «خارج المركز» وينتج عنه بقاء قواعد إضافية على خصلة واحدة (نتوء).تُعرف هذه النيوكليوتيدات المتناوبة (P) بسبب الطبيعة المتناوبة للتسلسل الناتج عندما تحل إنزيمات إصلاح الحمض النووي المتراكمة.إن عملية فتح دبوس الشعر بواسطة أرتيميس هي خطوة حاسمة لإعادة التركيب V (D) J وهي معيبة في نموذج الفأر الذي يعاني من نقص المناعة الشديد (scid).
بعد ذلك، يقوم XRCC4 وسيرنوسيس و DNA-PK بمحاذاة نهايات الحمض النووي وتجنيد ديوكسينوكليوتيديل ترانسفيراز (TdT)، وهو بوليميريز DNA مستقل عن القالب يضيف نيوكليوتيدات غير مقولبة (N) إلى نهاية الترميز.تكون الإضافة عشوائية في الغالب، لكن TdT يُظهر تفضيلًا لنيوكليوتيدات G / C.كما هو الحال مع جميع بوليمرات الحمض النووي المعروفة، يضيف TdT النيوكليوتيدات إلى حبل واحد في اتجاه 5 'إلى 3'.
أخيرًا، يمكن للنوكليازات الخارجية إزالة القواعد من نهايات التشفير (بما في ذلك أي نيوكليوتيدات P أو N قد تكون قد تكونت).تقوم بوليميرات الحمض النووي λ وμ بإدخال نيوكليوتيدات إضافية حسب الحاجة لجعل الطرفين متوافقين للانضمام.هذه عملية عشوائية، لذلك يمكن أن يحدث أي مزيج من إضافة نيوكليوتيدات P و N وإزالة الحالة الخارجية (أو لا يحدث أي شيء على الإطلاق).أخيرًا، يتم ربط نهايات التشفير المعالجة معًا بواسطة DNA ligase IV.
تؤدي كل أحداث المعالجة هذه إلى منطقة ارتباط بمولد الضد شديدة التباين، حتى عند إعادة تجميع نفس المقاطع الجينية.يسمح إعادة التركيب V (D) J بتوليد الغلوبولين المناعي ومستقبلات الخلايا التائية لمولدات المضادات التي لا يحتاج الكائن الحي ولا سلفه (أسلافه) إلى مواجهتها سابقًا، مما يسمح باستجابة مناعية تكيفية لمسببات الأمراض الجديدة التي تتطور أو تلك التي تتطور بشكل متكرر. التغيير (مثل الأنفلونزا الموسمية).ومع ذلك، فإن التحذير الرئيسي لهذه العملية هو أن تسلسل الحمض النووي يجب أن يظل في الإطار من أجل الحفاظ على تسلسل الأحماض الأمينية الصحيحة في منتج البروتين النهائي.إذا كان التسلسل الناتج خارج الإطار، فسيتم إيقاف تطور الخلية، ولن تستمر الخلية حتى النضج.لذلك فإن إعادة التركيب V (D) J هي عملية مكلفة للغاية ويجب أن تخضع (وهي) لتنظيم ومراقبة صارمة.
انظر أيضًا
مراجع
- "معلومات عن تأشيب جسمي على موقع amigo.geneontology.org"، amigo.geneontology.org، مؤرشف من الأصل في 14 أكتوبر 2017.
- "معلومات عن تأشيب جسمي على موقع meshb.nlm.nih.gov"، meshb.nlm.nih.gov، مؤرشف من الأصل في 18 سبتمبر 2019.
- "معلومات عن تأشيب جسمي على موقع jstor.org"، jstor.org، مؤرشف من الأصل في 11 أبريل 2020.
 بوابة علم الأحياء
بوابة علم الأحياء بوابة طب
بوابة طب بوابة الكيمياء الحيوية
بوابة الكيمياء الحيوية