مستقبلات الدوبامين
مستقبلات الدوبامين هي فئة من مجموعة المستقبلات المقترنة بالبروتين ج البارزة في الجهاز العصبي المركزي الفقاري (الجهاز العصبي المركزي). والناقلات العصبية الدوبامينية هي الربيطات الداخلية الأولية لمستقبلات الدوبامين.
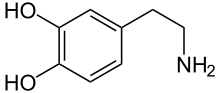
و تضلع مستقبلات الدوبامين في العديد من العمليات العصبية، بما في ذلك الدوافع والاستمتاع والإدراك والذاكرة والتعلم والتحكم الحركي الجيد، فضلا عن تعديل إشارات الخلايا العصبية الصماء. ان إشارات مستقبلات الدوبامين غير الطبيعية والوظيفة العصبية الدوبامينية تتضمن العديد من الاضطرابات العصبية والنفسية.[1] وهكذا، فان مستقبلات الدوبامين اهداف شائعة لعقاقير الجهاز العصبي؛ اما الأدوية المضادة للذهان فهي في كثير من الأحيان مضادات مستقبلات الدوبامين بينما المنشطات عادة ما تكون ناهضات غير مباشرة لمستقبلات الدوبامين.
الأنواع الفرعية لمستقبلات الدوبامين
هناك على الأقل خمسة أنواع فرعية من مستقبلات الدوبامين وهي D1 و D2 و D3 وD4 و D5.
مستقبلات D1 و D5 هي أعضاء في شبه عائلة D1 لمستقبلات الدوبامين، في حين أن مستقبلات D2 و D3 و D4 هي أعضاء في شبه عائلة D2. وهناك أيضا بعض الدلائل التي تشير إلى وجود محتمل لمستقبلات الدوبامين D6 و D7، ولكن لم يتم تحديد هذه المستقبلات بصورة قاطعة.[2]
على المستوى العالمي، فان مستقبلات D1 لها اصطلاحات واسعة النطاق خلال الدماغ. وعلاوة على ذلك، تم العثور على أنواع فرعية من مستقبلات D1-2 على المستويات من 10-100 مرة من الأنواع الفرعية لمستقبلات D3-5.[3]
شبه عائلة-D1
يقترن تنشيط مستقبلات شبه عائلة D1 مع بروتين ج Gαs، الذي ينشط فيما بعد إنزيم محلقة الأدينيلات، وبزيادة التركيز داخل الخلايا لنظام المرسال الثاني لمركب حلقي أدينوزين أحادي الفوسفات cAMP.
- يتم ترميز D1 من قبل جين مستقبل الدوبامين D1.
- يتم ترميز D5 من قبل جين مستقبل الدوبامين D5.
شبه عائلة -D2
يقترن تنشيط مستقبلات شبه عائلة D2 مع بروتين ج Gαi، والذي يثبط بشكل مباشر تشكيل حلقي أدينوزين أحادي الفوسفات cAMP عن طريق تثبيط انزيم محلقة الأدينيلات.[4]
- يتم ترميز D1 من قبل جين مستقبل الدوبامين D1، والذي يوجد منه نوعان : D2Sh (قصيرة) وD2Lh (طويلة) :
- نموذج D2Sh هو يقع ما قبل التشابك، وله وظائف تغييرية (بمعنى ان المستقبلات التلقائية، التي تنظم الناقلات العصبية بواسطة آليات التغذية المرتجعة، تؤثر على تجميع وتخزين وإطلاق الدوبامين في الشق المشبكي).
- نموذج D2Lh قد يعمل بمثابة مستقبلات تقليدية لما بعد التشابك العصبي، أي نقل المعلومات (اما بطريقة استثارية أو بطريقة مثبطة) إلا إذا منعت من قبل مستقبلات مضادة أو محرضات اصطناعية جزئية.
- يتم ترميز D3 من قبل جين مستقبل الدوبامين D3. ويلاحظ الاصطلاح الأعلى من مستقبلات الدوبامين D3 في جزر كاليخا والنواة المتكئة.[5]
- يتم ترميز D4 بواسطة جين مستقبل الدوبامين D4. جينات مستقبلات D4 تعرض أشكال متعددة تختلف في عدد ظهورها متغيرة ومكررة جنبا إلى جنب ضمن سلسلة ترميز إكسون 3.[6] بعض هذه الأليلات المقترنة مع زيادة معدل انتشار أمراض معينة. على سبيل المثال، الأليلات D4.7 انشأت جمعيتها مع اضطراب قصور الانتباه وفرط الحركة.[7][8][9]
دور مستقبلات الدوبامين في نظام الجهاز العصبي المركزي
مستقبلات الدوبامين تتحكم بالإشارات العصبية التي تنظم الكثير من السلوكيات المهمة، من قبيل الذاكرة العاملة المكانية.[10] وعلى الرغم من أن مستقبلات الدوبامين تتوزع بشكل واسع في الدماغ، إلا أن مناطق مختلفة لها مستقبلات مختلفة في كثافة أنواعها مما يعكس افتراضا أدوارا وظيفية مختلفة.
مستقبلات الدوبامين لغير الجهاز العصبي المركزي
نظام القلب والرئتين
في البشر، يوضح الشريان الرئوي D1، D2، D4 و D5 وأنواع فرعية من المستقبلات، والتي تؤثر في الدم عن طريق الدوبامين وتوسع الاوعية.[11] اما في الجرذان فان مستقبلات - D1 موجودة حول العضلات الملساء في الأوعية الدموية في معظم الأعضاء الرئيسية.[12]
وقد تم تحديد مستقبلات D4 في الأذينين في قلوب الجرذان وقلوب البشر. زيادات الدوبامين [13] تؤدي إلى انقباض عضلة القلب والنتاج القلبي، من دون تغيير معدل ضربات القلب عن طريق إشارات عبر مستقبلات الدوبامين.[2]
النظام الكلوي
مستقبلات الدوبامين توجد على طول الكليون في الكلى مع نبيبات صغيرة قريبة من الخلايا الظهارية والتي تبين أعلى كثافة.[12] في الجرذان فان مستقبلات - D1 توجد على الأجهزة المجاورة للكبيبة والنبيبات الكلوية بينما مستقبلات - D2 توجد على النبيبات الكلوية والكبيبات وخلف اطراف عقد الجهاز العصبي الودي وفي ذات منطقة الخلايا الكبيبية في القشرة الكظرية.[12] ولذا فان الدوبامين يؤثر على إدرار البول وإفراز الصوديوم في البول بتأثيره على طريقة عمل الكلى.[2]
مستقبلات الدوبامين في المرض
يتضمن الاختلال الوظيفي للناقلات العصبية التي تعمل على الدوبامين في الجهاز العصبي المركزي مجموعة متنوعة من الاضطرابات العصبية والنفسية، بما في ذلك الرهاب الاجتماعي [14]، ومتلازمة توريت [15]، ومرض باركنسون[16]، والفصام [15] متلازمة الذهان الخبيثة,[17] قصور الانتباه وفرط الحركة [18]، وإدمان المخدرات والكحول.[15][19]
قصور الانتباه وفرط الحركة
و على مدى سنوات عديدة تم الاعتراف بمستقبلات الدوبامين باعتبارها مسببات هامة لقصور الانتباه وفرط الحركة ADHD. ان العقاقير المستخدمة لعلاج اعاقة ADHD، بما في ذلك ميثيل فينيدات والأمفيتامين، لها آثار هامة على إشارات الدوبامين في الدماغ. وقد تضمنت دراسات الجمعيات مختصة عدة جينات ضمن مسارات إشارات الدوبامين ؛ وبصفة خاصة كان D4.7 وهو البديل ل D4 يظهر على الدوام ليكون أكثر تكررا في المرضى الذين يعانون من اعاقة ADHD.[20] مرضى اعاقة ADHD مع الأليل 4,7 يميلون ليكون ادائهم الإدراكي أفضل ونتائجهم أفضل على المدى البعيد مقارنة مع المرضى دون الاليل 4,7، مما يوحي بأن الاليل يرتبط مع شكل أكثر اعتدالا لاعاقة ADHD.[20]
وقد ثبط أليل D4.7 التعبير الجيني بالمقارنة مع المتغيرات الأخرى.[21]
الترفيه بتعاطي المخدرات واساءة الاستخدام
الدوبامين هو ناقل عصبي أساسي يشارك في مسارات الإثابة في الدماغ. وهكذا، فالعقاقير التي تزيد إشارات الدوبامين تنتج تأثيرات بهيجة. الكثير من العقاقير الترفيهية مثل الأمفيتامينات تغير الأداء الوظيفي لنقل الدوبامين (DAT)، والبروتينات المسؤولة عن إزالة الدوبامين من المشبك العصبي. عندما يتم حظر نشاط DAT، فان كيمياء الأعصاب تتدفق مع الدوبامين وتزيد إشارات الدوبامين. عند حدوث ذلك، لا سيما في النواة المتكئة [22] تزداد إشارات مستقبلات D1[19] و D2[22] مما يقوم بالوساطة في «مكافأة» التحفيز لاستهلاك المخدرات.[22]
الفصام
بينما هناك دلائل على أن منظومة الدوبامين تشترك في حدوث الفصام، إلا أن النظرية القائلة بأن النشاط المفرط لتوصيل الإشارة الدوباميني يحث المرض نظرية مثيرة للجدل. المنشطات، مثل الأمفيتامين والكوكايين تحرض تغييرات جذرية في إشارات الدوبامين؛ فالجرعات الكبيرة والاستخدام المطول يمكن ان يحفز أعراض تشبه الفصام. بالإضافة إلى ذلك، العديد من العقاقير المضادة للذهان تستهدف مستقبلات الدوبامين، بخاصة مستقبلات D2.
ارتفاع ضغط الدم الجيني
يمكن لطفرات مستقبلات الدوبامين ان تسبب ارتفاع ضغط الدم الجيني لدى البشر.[23] وهذا يمكن أن يحدث في النماذج الحيوانية والبشر مع خلل في نشاط مستقبلات الدوبامين، لا سيما D1.[12]
تنظيم الدوبامين
تكون مستقبلات الدوبامين في حالة مستقرة عادة، ولكن الازدياد أو النقصان الحاد (و أحيانا لفترات طويلة) في مستويات الدوبامين (بصورة رئيسية عن طريق المنشطات أو مضادات الذهان) يمكن أن تؤدي إلى تقليل ضبط (تقليل أعداد) أو زيادة ضبط (زيادة أعداد) مستقبلات الدوبامين. مع المنشطات، يرتيط تقليل الضبط عادة مع فقدان الرغبة في الأنشطة الممتعة، وتقصير مدى الانتباه، وسلوكيات السعي لتعاطي المخدرات. اما مع مضادات الذهان، فان زيادة الضبط يمكن ان تؤدي إلى خلل الحركة المتأخر (العضلات الجيدة، مثل الوجه، الاختلاج اللا ارادي)، أو خلل الحركة العادي فقط (نفسه ولكنه مؤقت فقط).
هالوبيريدول، وبعض مضادات الذهان الأخرى، قد اظهرت زيادة نشاط مستقبلات D2.[24] هالوبيريدول تستحث الأنشطة لتصل إلى 98 ٪ أعلى من خط الأساس في ما يقرب من فترة اسبوعين، ولكنها اسفرت عن أعراض جانبية كبيرة في خلل الحركة. دراسة أخرى [25] أظهرت انه في الكائنات الحية، تزيد مستقبلات الدوبامين 2 تقريبا 90بنسبة ٪ بعد العلاج المضاد للذهان.
توجد تقارير مختلفة في سوء استعمال المنشطات، والتنظيم الأعلى / الأسفل. فطبقا لإحدى الدراسات،[26] فان الكوكايين والهيروين والمنشطات، والكحول، والنيكوتين تسبب جميعها نقصانا في كمية مستقبلات الدوبامين 2. وهذا الاقتران قد تم ربطه مع إدمان المواد الغذائية، لأن قلة توافر مستقبلات الدوبامين موجودة في الأشخاص الأكثر استهلاكا للمواد الغذائية.[27][28] مقالة الأخبار الأخيرة [29] بين ملخص دراسة بركهافن المخبرية الوطنية في وزارة الطاقة الأمريكية أن زيادة مستقبلات الدوبامين مع المعالجات الوراثية يؤدي إلى انخفاض في استهلاك الكوكايين بنسبة تصل إلى 75 ٪، ولكنها دامت لمدة ستة أيام فقط.
في دراسة مقارنة للكوكايين [30] اظهرت ان زيادة ضبط (زيادة أعداد) الكوكايين في مستقبلات D3 في النواة المتكئة، قد يسهم في سلوك السعي لتعاطي المخدرات.
انظر أيضًا
وصلات خارجية
- "مستقبلات الدوبامين"، IUPHAR Database of Receptors and Ion Channels، International Union of Basic and Clinical Pharmacology، مؤرشف من الأصل في 01 فبراير 2017.
- Zimmerberg, B., "مستقبلات الدوبامين : العائلة الممثلة لمستقبلات الميتابوتروبيك, Multimedia Neuroscience Education Project (2002)
- مادة السكولاربيديا في علم تشريح الدوبامين
المراجع
- Girault J, Greengard P (2004)، "The neurobiology of dopamine signaling"، Arch Neurol، 61 (5): 641–4، doi:10.1001/archneur.61.5.641، PMID 15148138.
- Contreras F, Fouillioux C, Bolívar A, Simonovis N, Hernández-Hernández R, Armas-Hernandez M, Velasco M (2002)، "Dopamine, hypertension and obesity"، J Hum Hypertens، 16 Suppl 1: S13–7، doi:10.1038/sj.jhh.1001334، PMID 11986886.
{{استشهاد بدورية محكمة}}: صيانة CS1: أسماء متعددة: قائمة المؤلفون (link) - Hurley MJ, Jenner P (2006)، "What has been learnt from study of dopamine receptors in Parkinson's disease?"، Pharmacol Ther.، 111 (3): 715، doi:10.1016/j.pharmthera.2005.12.001، PMID 16458973.
- Neves SR, Ram PT, Ravi Iyengar (مايو 2002)، "G protein pathways"، Science (journal)، 296 (5573): 1636–9، doi:10.1126/science.1071550، PMID 12040175.
{{استشهاد بدورية محكمة}}: صيانة CS1: أسماء متعددة: قائمة المؤلفون (link) - Suzuki M, Hurd YL, Sokoloff P, Schwartz JC, Sedvall G (يناير 1998)، "D3 dopamine receptor mRNA is widely expressed in the human brain"، Brain Res.، 779 (1–2): 58–74، doi:10.1016/S0006-8993(97)01078-0، PMID 9473588.
{{استشهاد بدورية محكمة}}: صيانة CS1: أسماء متعددة: قائمة المؤلفون (link) - NCBI Database نسخة محفوظة 04 فبراير 2015 على موقع واي باك مشين.
- Manor I, Tyano S, Eisenberg J, Bachner-Melman R, Kotler M, Ebstein RP (2002)، "The short DRD4 repeats confer risk to attention deficit hyperactivity disorder in a family-based design and impair performance on a continuous performance test (TOVA)"، Mol. Psychiatry، 7 (7): 790–4، doi:10.1038/sj.mp.4001078، PMID 12192625.
{{استشهاد بدورية محكمة}}: صيانة CS1: أسماء متعددة: قائمة المؤلفون (link) - Langley K, Marshall L, van den Bree M, Thomas H, Owen M, O'Donovan M, Thapar A (يناير 2004)، "Association of the dopamine D4 receptor gene 7-repeat allele with neuropsychological test performance of children with ADHD"، Am J Psychiatry، 161 (1): 133–8، doi:10.1176/appi.ajp.161.1.133، PMID 14702261.
{{استشهاد بدورية محكمة}}: صيانة CS1: أسماء متعددة: قائمة المؤلفون (link) - Kustanovich V, Ishii J, Crawford L, Yang M, McGough JJ, McCracken JT, Smalley SL, Nelson SF (يوليو 2004)، "Transmission disequilibrium testing of dopamine-related candidate gene polymorphisms in ADHD: confirmation of association of ADHD with DRD4 and DRD5"، Mol. Psychiatry، 9 (7): 711–7، doi:10.1038/sj.mp.4001466، PMID 14699430.
{{استشهاد بدورية محكمة}}: صيانة CS1: أسماء متعددة: قائمة المؤلفون (link) - Williams G, Castner S (2006)، "Under the curve: critical issues for elucidating D1 receptor function in working memory"، Neuroscience، 139 (1): 263–76، doi:10.1016/j.neuroscience.2005.09.028، PMID 16310964.
- Ricci A, Mignini F, Tomassoni D, Amenta F (2006)، "Dopamine receptor subtypes in the human pulmonary arterial tree"، Auton Autacoid Pharmacol، 26 (4): 361–9، doi:10.1111/j.1474-8673.2006.00376.x، PMID 16968475.
{{استشهاد بدورية محكمة}}: صيانة CS1: أسماء متعددة: قائمة المؤلفون (link) - Hussain T, Lokhandwala M (2003)، "Renal dopamine receptors and hypertension"، Exp Biol Med (Maywood)، 228 (2): 134–42، PMID 12563019.
- Ricci A, Bronzetti E, Fedele F, Ferrante F, Zaccheo D, Amenta F (1998)، "Pharmacological characterization and autoradiographic localization of a putative dopamine D4 receptor in the heart"، J Auton Pharmacol، 18 (2): 115–21، doi:10.1046/j.1365-2680.1998.1820115.x، PMID 9730266.
{{استشهاد بدورية محكمة}}: صيانة CS1: أسماء متعددة: قائمة المؤلفون (link) - Schneier FR, Liebowitz MR, Abi-Dargham A, Zea-Ponce Y, Lin SH, Laruelle M (2000)، "Low dopamine D(2) receptor binding potential in social phobia"، Am J Psychiatry، 157 (3): 457–459، doi:10.1176/appi.ajp.157.3.457، PMID 10698826.
{{استشهاد بدورية محكمة}}: صيانة CS1: أسماء متعددة: قائمة المؤلفون (link) - Kienast T, Heinz A (2006)، "Dopamine and the diseased brain"، CNS Neurol Disord Drug Targets، 5 (1): 109–31، doi:10.2174/187152706784111560، PMID 16613557.
- Fuxe K, Manger P, Genedani S, Agnati L (2006)، "The nigrostriatal DA pathway and Parkinson's disease"، J Neural Transm Suppl، 70 (70): 71–83، doi:10.1007/978-3-211-45295-0_13، PMID 17017512.
{{استشهاد بدورية محكمة}}: صيانة CS1: أسماء متعددة: قائمة المؤلفون (link) - Mihara K, Kondo T, Suzuki A؛ وآخرون (2003)، "Relationship between functional dopamine D2 and D3 receptors gene polymorphisms and neuroleptic malignant syndrome"، Am. J. Med. Genet. B Neuropsychiatr. Genet.، 117 (1): 57–60، doi:10.1002/ajmg.b.10025، PMID 12555236.
{{استشهاد بدورية محكمة}}: Explicit use of et al. in:|مؤلف=(مساعدة)صيانة CS1: أسماء متعددة: قائمة المؤلفون (link) - Faraone S, Khan S (2006)، "Candidate gene studies of attention-deficit/hyperactivity disorder"، J Clin Psychiatry، 67 Suppl 8: 13–20، PMID 16961425.
- Hummel M, Unterwald E (2002)، "D1 dopamine receptor: a putative neurochemical and behavioral link to cocaine action"، J Cell Physiol، 191 (1): 17–27، doi:10.1002/jcp.10078، PMID 11920678.
- Gornick M, Addington A, Shaw P, Bobb A, Sharp W, Greenstein D, Arepalli S, Castellanos F, Rapoport J (2007)، "Association of the dopamine receptor D4 (DRD4) gene 7-repeat allele with children with attention-deficit/hyperactivity disorder (ADHD): An update"، Am J Med Genet B Neuropsychiatr Genet، 144 (3): 379–82، doi:10.1002/ajmg.b.30460، PMID 17171657.
{{استشهاد بدورية محكمة}}: صيانة CS1: أسماء متعددة: قائمة المؤلفون (link) - Schoots O, Van Tol H (2003)، "The human dopamine D4 receptor repeat sequences modulate expression"، Pharmacogenomics J، 3 (6): 343–8، doi:10.1038/sj.tpj.6500208، PMID 14581929.
- Di Chiara G, Bassareo V, Fenu S, De Luca M, Spina L, Cadoni C, Acquas E, Carboni E, Valentini V, Lecca D (2004)، "Dopamine and drug addiction: the nucleus accumbens shell connection"، Neuropharmacology، 47 Suppl 1: 227–41، doi:10.1016/j.neuropharm.2004.06.032، PMID 15464140.
{{استشهاد بدورية محكمة}}: صيانة CS1: أسماء متعددة: قائمة المؤلفون (link) - Jose P, Eisner G, Felder R (2003)، "Regulation of blood pressure by dopamine receptors"، Nephron Physiol، 95 (2): p19–27، doi:10.1159/000073676، PMID 14610323.
{{استشهاد بدورية محكمة}}:|صفحات=has extra text (مساعدة)صيانة CS1: أسماء متعددة: قائمة المؤلفون (link) - Silvestri S, Seeman MV, Negrete JC؛ وآخرون (أكتوبر 2000)، "Increased dopamine D2 receptor binding after long-term treatment with antipsychotics in humans: a clinical PET study"، Psychopharmacology، 152 (2): 174–80، doi:10.1007/s002130000532، PMID 11057521.
{{استشهاد بدورية محكمة}}: Explicit use of et al. in:|مؤلف=(مساعدة)صيانة CS1: أسماء متعددة: قائمة المؤلفون (link) - Farde L, Hall H, Ehrin E, Sedvall G (يناير 1986)، "Quantitative analysis of D2 dopamine receptor binding in the living human brain by PET"، Science (New York, N.Y.)، 231 (4735): 258–61، doi:10.1126/science.2867601، PMID 2867601.
{{استشهاد بدورية محكمة}}: صيانة CS1: أسماء متعددة: قائمة المؤلفون (link) - Fehr C, Yakushev I, Hohmann N؛ وآخرون (أبريل 2008)، "Association of low striatal dopamine d2 receptor availability with nicotine dependence similar to that seen with other drugs of abuse"، The American journal of psychiatry، 165 (4): 507–14، doi:10.1176/appi.ajp.2007.07020352، PMID 18316420.
{{استشهاد بدورية محكمة}}: Explicit use of et al. in:|مؤلف=(مساعدة)صيانة CS1: أسماء متعددة: قائمة المؤلفون (link) - Paul Park (09 أغسطس 2007)، "Food Addiction: From Drugs to Donuts, Brain Activity May be the Key"، مؤرشف من الأصل في 20 سبتمبر 2018.
- Paul M Johnson& Paul J Kenny (28 مارس 2010)، "Dopamine D2 receptors in addiction-like reward dysfunction and compulsive eating in obese rats"، مؤرشف من الأصل في 1 أبريل 2010. Nature Neuroscience Year published:(2010)DOI:doi:10.1038/nn.2519
- "Gene Therapy For Addiction: Flooding Brain With 'Pleasure Chemical' Receptors Works On Cocaine, As On Alcohol"، 18 أبريل 2008، مؤرشف من الأصل في 27 سبتمبر 2018.
- Staley JK, Mash DC (أكتوبر 1996)، "Adaptive increase in D3 dopamine receptors in the brain reward circuits of human cocaine fatalities"، The Journal of neuroscience : the official journal of the Society for Neuroscience، 16 (19): 6100–6، PMID 8815892، مؤرشف من الأصل في 24 أكتوبر 2010.
 بوابة علم الأحياء الخلوي والجزيئي
بوابة علم الأحياء الخلوي والجزيئي بوابة الكيمياء الحيوية
بوابة الكيمياء الحيوية