نقل الغشاء
في علمِ الأَحياء الخَلوي، يُشير النَقل الغِشائيّ إلى مَجموعةِ الآليات التي تُنظم مُرور المواد المُذابَة مثل الأَيونات والجُزيئات الصَغيرة عَبر الأَغشية البَيولوجيّة، وَهي طَبقات ثُنائيّة الليبيد تَحتَوي على بروتينات مُضمَنَة فيها. يرجع تنظيم المرور عبر الغشاء إلى نفاذية الغشاء الانتقائي وهي خاصية للأغشية البَيولوجيّة تسمح لها بفصل المواد ذات الطبيعة الكيميائيّة المُميزة. بمعنى آخر، يمكن أن تكون نَفاذِيَّة لبعضِ المواد دون غيرها.[1]
تتم حركات معظم الْمَوَادّ المُذابَة عَبر الغِشاء بِوساطةِ بروتينات نَقل الغِشاء المُتَخصِصة بِدرجاتٍ مُتَفاوِِتَة في نقلِ جُزيئاتٍ مُعينة. نظراً لأن تَنوع الخلايا المُتَميزة وعلم وَظائِفها مُرتَبط ارتِباطاََ وَثيقاً بِقُدراتِها على جذبِ عناصرَ خارجيّة مُختَلفة، فقد تم نشر مَجموعَة مِنْ بروتينات النَقل المُحددة لِكلِ نوع خلية ولكل مَرحلة فِسْيولوجِيّة مُحددة.[1] يتم تنظيم هذا التعبير التَفاضُليّ مِن خلال النسخ التَفاضُليّ لترميز الجينات لهذه البروتينات وترجمتها، على سبيل المثال، من خلال الآليات الجُزيئيّة الجينيّة، ولكن أيضاً على مستوى بَيُولُوجْيا الخلية: يمكنُ تنشيط إنتاج هذه البروتينات عن طريق مسارات الإشارات الخَلويّة، على المستوى البَيو كيميائيّ، أو حتى عن طريقِ التَواجُد في الحُويصلات السيتوبلازميّة.[2]
خلفية
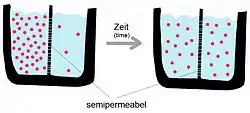
مِن الناحيةِ الدِينَامِيكيَّة الحَرارِيَّة، يمكن أن يحدث تَدفق الْمَوَادّ مِن حَيز إلى أخر في اتجاهِ التركيز أو التَدرج الكهرو كيميائيّ أو عكسه. إذا حدث تبادل المواد في اتجاه التدرج، أي في اتجاه تناقص الإمكانات، فلا توجد حاجة لمدخل للطاقة من خارج النظام؛ ومع ذلك، إذا كان النقل ضد التَدَرج، فَسوفَ يَتَطلب إدخال الطاقة، الطاقة الأَيضيّة في هَذهِ الحالة.[3] على سبيل المثال، آلية الفصل الكيميائيّة الكلاسيكيّة التي لا تتطلب إضافة طاقة خارجيّة هي غسيل الكلى. في هذا النظام، يفصل غشاء شبه منفذ بين محلولين بتركيز مختلف من نفس المادة المُذابة. إذا كان الغِشاء يسمح بمرور الماء ولكن ليس المذاب، فسوف ينتقل الماء إلى الحيز الذي يحتوي على أكبر تركيز مذاب من أجل إنشاء توازن تكون فيه طاقة النظام عند الحد الأدنى. يحدث هذا لأن الماء ينتقل من تركيز مُذيب عالي إلى تركيز منخفض (من حيث المذاب، يحدث العكس) ولأن الماء يتحرك على طول التدرج، فلا حاجة لمدخل خارجي للطاقة.
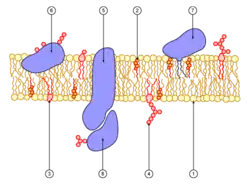
طبيعة الأغشيّة البيولوجيّة، وخاصةً الليبيد، هي مُزدوجة الألفة، لأنها تشكل طبقات ثُنائيّة تحتوي على طبقةٍ داخليّة كارهة للماء وطبقة خارجيّة مُحبة للماء. يجعل هذا الهيكل النقل مُمكناََ عن طريقِ الانتشار البسيط أو السلبي، والذي يتكون من انتشار المواد عبر الغشاء دون إنفاق الطاقة الأيضيّة ودون مساعدة بروتينات النقل. إذا كانت المادة المنقولة تحتوي على شحنة كهربائية صافية، فلن تتحرك فقط استجابةً لتدرج التركيز، ولكن أيضاََ إلى التدرج الكهر كيميائي بسبب جهد الغشاء.
| نوع المادة | أمثلة | سلوك |
|---|---|---|
| غازات | ثاني أكسيد الكربون، نيتروجين، أكسجين | منفذة |
| جزيئات قطبية صغيرة غير مشحونة | اليوريا، الماء، الإيثانول | قابل لنفاذ، كلياََ أو جزئياََ |
| جزيئات قطبية كبيرة غير مشحونة | الغلوكوز والفركتوز | غير منفذة |
| أيونات | K+، Na+، Cl−،HCO3 | غير منفذة |
| جزيئات قطبية مشحونة | أدينوسين ثلاثي الفوسفات، أحماض أمينية، غلوكوز-٦-فوسفات | غير منفذة |
نظراً لأن القليل من الجزيئات قادرة على الانتشار من خلال غشاء دهني، فإن غالبية عمليات النقل تتضمن بروتينات النقل. تمتلك بروتينات الغشاء هذه عدداََ كبيراً من لولب ألفا مغمورة في مَطْرِس الدهون. توجد هذه البروتينات في البكتيريا في شكل صحيفة بيتا.[4] من المحتمل أن تتضمن هذه البِنْيَة قناة عبر بيئات بروتينية محبة للماء تسبب اضطراباً في الوسط شديد الكراهية للماء الذي تشكله الدهون. يمكن أن تشارك هذه البروتينات في النقل بعدة طرق: فهي تعمل كمضخات مدفوعة بـ أدينوسين ثلاثي الفوسفات، أي بالطاقة الأيضية، أو كقنوات للانتشار المسهل.نظراً لأن القليل من الجزيئات قادرة على الانتشار من خلال غشاء دهني، فإن غالبية عمليات النقل تتضمن بروتينات النقل. تمتلك بروتينات الغشاء هذه عدداََ كبيراً من لولب ألفا مغمورة في مَطْرِس الدهون. توجد هذه البروتينات في البكتيريا في شكل صحيفة بيتا. من المحتمل أن تتضمن هذه البِنْيَة قناة عبر بيئات بروتينية محبة للماء تسبب اضطراباً في الوسط شديد الكراهية للماء الذي تشكله الدهون. يمكن أن تشارك هذه البروتينات في النقل بعدة طرق: فهي تعمل كمضخات مدفوعة بـ أدينوسين ثلاثي الفوسفات، أي بالطاقة الأيضية، أو كقنوات للانتشار المسهل.
الانتشار السلبي والانتشار النشط
كما ذكر أعلاه، فإن الانتشار السلبي هو ظاهرة تلقائية تزيد من إنتروبيا النظام وتقلل من الطاقة الحرة.[5] تتأثر عملية النقل بخصائص المادة المنقولة وطبيعة الطبقة الثنائية. سرعة الانتشار في غشاء الفسفوليبيد النقي ستعتمد على:
- تدرج التركيز.
- عدم الألفة للماء.
- الحجم.
- الشحنة. إذا كان للجزيء شحنة صافية.
- درجة الحرارة.
النقل النشط والنقل المشترك
في النقلِ النشط، يتم تحريك المُذاب مُقابل التركيز أو التدرج الكهركيميائي؛ عند القيام بذلك، تَستَهلك بروتينات النقل المَعنية الطاقة الأَيضيّة، عادةً أدينوسين ثلاثي الفوسفات. في النقلِ النشط الأولي، يحدث التحلل المائي لمُزود الطاقة (مثل أدينوسين ثلاثي الفوسفات) مباشرة من أجل نقل المُذاب المعني، على سبيل المثال، عندما تكون بروتينات النقل عبارة عن إنزيمات أدينوسين ثلاثي الفوسفات. عندما يكون التحلل المائي لمزود الطاقة غير مباشر كما هو الحال في النقل النشط الثانوي، يتم استخدام الطاقة المُخَزنَة في التدرج الكهر كيميائي. على سبيل المثال، في النقل المشترك، يتم استخدام تدرجات بعض المواد المذابة لنقل مركب مستهدف مقابل تدرجه، مما يتسبب في تبديد تدرج المذاب. قد يبدو في هذا المثال، أنه لا يوجد استخدام للطاقة، ولكن التحلل المائي لِمُزود الطاقة مطلوب لتحديد تدرج المذاب المنقول مع المركب المستهدف. سيتم إنشاء تدرج المادة المذابة المنقولة بشكل مشترك من خلال استخدام أنواع معينة من البروتينات تسمى المَضخات البيوكيميائيّة.[2]
جاءَ اكتشاف وجود هذا النوع مِن البروتين الناقلِ مِن دِراسة حَركية نقل الجُزيئات عَبر الغِشاء. بِالنسبَةِ لِبعضِ الْمَوادّ المُذابَة، لوحِظَ أن سرعة النقل وَصلت إلى الهضبة عِندَ تركيز مُعين أعلاه لم يَكن هناك زيادة كبيرة في مُعدلِ الامتِصاص، مما يُشير إلى استجابة نوع منحنى لوغاريتمي. فُسِرَ ذلك على أنه يُوضح أن النقل تَمّ بِوساطة تكوين مركب ناقل الركيزة، والذي هو من الناحيةِ النظرية نفس مجمع الركيزة الإنزيميّة لِحركيةِ الإنزيم. لذلك، يحتوي كل بروتين ناقل على ثابت تقارب للمُذاب الذي يساوي تركيز المُذاب عندما تكون سرعة النقل نصف قيمتها القصوى. وهذا يعادل في حالة إنزيم ثابت ميكايليس ومينتين.[6][7]
بعض المَيزات الهامة للنقلِ النشط بالإضافةِ إلى قُدرتِهِ على التدخل حتى ضد التدرج، حركته واستخدام أدينوسين ثلاثي الفوسفات، هي انْتِقائيَّة عالية وسهولة التثبيط الدوائي الانْتِقائي.[7]
بروتينات ناقلة نشطة ثانوية
تقوم البروتينات الناقلة الثانوية النشطة بنقل جزيئين في نفس الوقت: أحدهما عكس التدْرُج والآخر مع التدْرُج يتم تمييزها وفقاً لاتجاه الجزيئين:
- تنادل متعاكس: (يسمى أيضاََ الْمُبَادِل أو النَّاقِل المُعاكِس): حَرَكَ الجزيء عكس تدرجه وفي نفس الوقت يزيح واحداََ أو أكثر من الأيونات على طول تدرجه. تتحرك الجُزيئات في اتجاهين مُتعاكِسين.
- مُرَاحِل: تحريك الجُزيء عكس تَدْرجه مع إزاحة واحد أو أكثر من الأيونات المختلفة على طول تدرجها. تتحرك الجُزيئات في نفسِ الاتجاه.
مضخات

المِضَخّة عبارة عن بروتين يُحلل جُزيء أدينوسين ثلاثي الفوسفات، لِنقلِ مُذاب مُعين عَبر غشاء، وبذلك يولد جهد غشاء متدرج كهركيميائي. هذا التَدْرج مهم كَمؤشر لِحالةِ الخلية مِن خِلال معلمات مثل احتمال نرنست. فيما يتعلق بنقل الغشاء، فإن التدرج مهم لأنه يساهم في تقليل إنتروبيا النظام في النقل المشترك للمواد مقابل تدرجها. من أهم المِضَخّات في الخلايا الحيوانيّة هي مِضخّة الصوديوم والبوتاسيوم التي تعمل بالآلية التالية:
- ربط ثلاثة أيونات صوديوم بمواقعها النشطة على المضخة المرتبطة بـ ATP.
- يتم تحلل ATP مما يؤدي إلى فسفرة الجانب السيتوبلازمي للمِضَخة، وهذا يؤدي إلى تغيير هيكل البروتين. تنتج الفسفرة عن نقل المجموعة الطرفية من ATP إلى بقايا الأسبارتات في بروتين النقل والإفراج اللاحق عن أدينوسين ثنائي الفوسفات.
- تغيير الهيكل في المِضَخة يعرض أيونات الصوديوم إلى الخارج. يحتوي الشكل المفسفر للمِضَخة على ألفة مُنخَفِضة لأيونات الصوديوم لذلك يتم إطلاقها.
- بِمُجرد تحرير أيونات الصوديوم، تقوم المِضَخة بربط جُزيئين مِن الكالسيوم بمواقع الترابط الخاصة بهما على الوجه خارج الخلية لبروتين النقل. يُِؤدي هذا إلى نزع الفوسفات عن المضخة، وإعادتها إلى حالتها التوافقية السابقة، ونقل أيونات الكالسيوم إلى الخلية.
- يحتوي الشكل مَنزوع الفوسفات للمِضَخة على تقارب أعلى لأيونات الصوديوم أكثر من أيونات الكالسيوم، لذلك يتم إطلاق أيونات المرتبطة في العصارة الخلوية. يرتبط ATP، وتبدأ العملية مرة أخرى.
انتقائية الغشاء
نظراً لأن السِمَة الرئيسية للانتقال عبر الغِشاء البَيُولُوجْي هي انتِقائيته وسلوكه اللاحق كحاجز لبعض المَوادّ، تمت دراسة فسيولوجيا هذه الظاهرة على نطاقٍ واسع. تم تقسيم التحقيق في انتقائية الغشاء بشكل كلاسيكي إلى تلك المتعلقة بالإلكتروليت و اللاإلكتروليت.
انتقائية الإلكترولايت.
تحدد القنوات الأيونية قُطراً داخلياً يسمح بمرور أيونات صغيرة مرتبطة بخصائص مختلفة للأيونات التي يمكن نقلها. نظراً لأن حجم الأيون مرتبط بأنواعه الكيميائية، يمكن الافتراض مسبقاً أن القناة التي كان قُطْر مَسامِها كافٍ للسماح بمرور أيون واحد ستسمح أيضاً بنقل الآخرين بحجم أصغر، ومع ذلك، هذا لا يحدث في مُعظم الحالات. هناك سِمَتان إلى جانب الحجم مهمتان في تحديد انتِقائيّة مَسام الغِشاء: سُهولة الجفاف وتفاعل الأيونات مع الشحنات الداخلية للمسام.[7] لكي يمر أيون عبر المسام، يجب أن ينفصل عن جُزيئات الماء التي تُغطيه في طبقات متتالية مِن الذوبان. يرتبط الميل إلى الجفاف، أو التسهيلات للقيام بذلك، بحجم الأيون: يمكن للأيونات الأكبر أن تفعل ذلك بسهولة أكبر من الأيونات الأصغر، بحيث تسمح المسام ذات المراكز القُطبية الضعيفة بمرور الأيونات الأكبر حجماََ بشكل تفضيلي. أصغر حجماً. عندما يتكون الجزء الداخلي للقناة من مجموعات قُطبيّة من السلاسل الجانبية للأحماضِ الأمينية المكونة،[8] يمكن أن يكون تفاعل الأيونات المجففة مع هذه المراكز أكثر أهمية من وسيلة الجفاف في إضفاء خصوصية القناة. على سبيل المثال، القناة المكونة من الهيستيدينات والأرجينينات، مع مجموعات موجبة الشحنة، سوف تطرد بشكل انتقائي الأيونات من نفس القُطبية، ولكنها ستسهل مرور الأيونات سالبة الشحنة. أيضاََ، في هذه الحالة، ستكون الأيونات الأصغر قادرة على التفاعل. بشكل أوثق بسبب الترتيب المكاني للجزيء (الفراغية)، مما يزيد بشكل كبير من تفاعلات الشحنة والشحنة وبالتالي يبالغ في التأثير.[7]
انتقائية اللاإلكترولايت
المَوادّ اللاإلكترولايتية، المَوادّ التي تكون بشكل عام كارهة للماء ومُحبَة للدهون، تمر عادة عبر الغشاء عن طريق الذوبان في طبقة ثنائيّة الدهون، وبالتالي، عن طريق الانتشار السلبي. بالنسبة لأولئك الذين ليس لديهم إلكتروليت يتم نقله عبر الغشاء بواسطة بروتين نقل، فإن القدرة على الانتشار تعتمد بشكل عام على معامل التقسيم K. شحنة لا إلكتروليتية مشحونة جزئياً، والتي تكون قُطبية إلى حد ما، مثل الإيثانول أو الميثانول أو اليوريا، قادرة على المرور عبر الغشاء من خلال القنوات المائيّة المغمورة في الغشاء. لا توجد آلية تنظيم فعالة تحد من هذا النقل، مما يشير إلى ضعف جوهري للخلايا لاختراق هذه الجزيئات.[7]
تكوين بروتينات نقل الغشاء
هناك العديد من قواعد البيانات التي تحاول إنشاء أشجار النُشوء والتَطور التي توضح بالتفصيل إنشاء البروتينات الناقِلَة. أحد هذه الموارد هو قاعدة بيانات تصنيف الناقل.[9]
انظر أيضاََ
مراجع
- Lodish؛ وآخرون (2005)، Biología celular y molecular (ط. Buenos Aires: Médica Panamericana)، ISBN 950-06-1374-3.
- Alberts؛ وآخرون (2004)، Biología molecular de la célula (ط. Barcelona: Omega)، ISBN 84-282-1351-8.
- Cromer, A.H. (1996)، Física para ciencias de la vida (باللغة الإسبانية) (ط. Reverté ediciones)، ISBN 84-291-1808-X.
- Prescott, L.M. (1999)، Microbiología (ط. McGraw-Hill Interamericana de España, S.A.U.)، ISBN 84-486-0261-7.
- Mathews C. K.؛ Van Holde, K.E؛ Ahern, K.G (2003)، Bioquímica (ط. 3rd)، ISBN 84-7829-053-2.
- Randall D؛ Burggren, W.؛ French, K. (1998)، Eckert Fisiología animal (ط. 4th)، ISBN 84-486-0200-5.
- "A comprehensive approach to the mathematical modeling of mass transport in biological systems: Fundamental concepts and models"، International Journal of Heat and Mass Transfer، 158: 199777، مؤرشف من الأصل في 20 ديسمبر 2021.
- Lehninger, Albert (1993)، Principles of Biochemistry, 2nd Ed. (ط. Worth Publishers)، ISBN 0-87901-711-2، مؤرشف من الأصل في 16 ديسمبر 2019.
- "Transporter Classification Database"، مؤرشف من الأصل في 03 يناير 2014، اطلع عليه بتاريخ 15 يوليو 2010.
 بوابة علم الأحياء
بوابة علم الأحياء بوابة علم الأحياء الخلوي والجزيئي
بوابة علم الأحياء الخلوي والجزيئي بوابة الكيمياء الحيوية
بوابة الكيمياء الحيوية