L-selectin
L-selectin, also known as CD62L, is a cell adhesion molecule found on the cell surface of leukocytes, and the blastocyst. It is coded for in the human by the SELL gene. L-selectin belongs to the selectin family of proteins, which recognize sialylated carbohydrate groups containing a Sialyl LewisX (sLeX) determinant.[5] L-selectin plays an important role in both the innate and adaptive immune responses by facilitating leukocyte-endothelial cell adhesion events.[6] These tethering interactions are essential for the trafficking of monocytes and neutrophils into inflamed tissue as well as the homing of lymphocytes to secondary lymphoid organs. L-selectin is also expressed by lymphoid primed hematopoietic stem cells and may participate in the migration of these stem cells to the primary lymphoid organs.[6] In addition to its function in the immune response, L-selectin is expressed on embryonic cells and facilitates the attachment of the blastocyst to the endometrial endothelium during human embryo implantation.[7]
| SELL | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Identifiers | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Aliases | SELL, CD62L, LAM1, LECAM1, LEU8, LNHR, LSEL, LYAM1, PLNHR, TQ1, selectin L | ||||||||||||||||||||||||||||||||||||||||||||||||||
| External IDs | OMIM: 153240 MGI: 98279 HomoloGene: 539 GeneCards: SELL | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Wikidata | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||


L-selectin is composed of multiple structural regions: an N-terminus C-type lectin domain, an adjacent epidermal growth factor-like domain, two to the consensus repeat units homologous to those found in C3/C4-binding proteins, an extracellular cleavage site, a short transmembrane domain, and a cytoplasmic tail. It is cleaved by ADAM17.[6][8]
Ligands
The nature of the interactions between L-selectin and ligand depends on many circumstances, primarily the location of anatomically defined sites in the high vessel venules (perivascular, extravascular and intravascular).
Because of the diversity of L-selectin ligands, signals that propagate downstream of L-selectin provide information about the position of the leukocyte within the multistep adhesion cascade (binding, rolling, adhesion, and transmigration).
While L-selectin ligands on the apical side of the endothelium have long been characterized as receptors for binding and rolling, glycans enriched on the basolateral side and in the basement membrane likely control quite different signals. The binding lifetime of L-selectin with apical ligands will be on the order of milliseconds, so in contrast, L-selectin-dependent adhesion in a microenvironment without hydrodynamic shear stress (e.g., within transmigrating pseudopods) will take seconds to minutes.
- GlyCAM-1, found in the high endothelial venules of the lymph nodes.
- CD34, found on endothelial cells.
- MadCAM-1, found on endothelial cells of gut-associated lymphoid tissue.
- PSGL-1, binds with low affinity.
Expression
L-selectin is expressed constitutively on most circulating leukocytes.[6] Over time, these molecules are released through the process of ectodomain shedding and are replaced by newly synthesized L-selectin proteins. Ectodomain shedding is largely accomplished through cleavage by ADAM17.
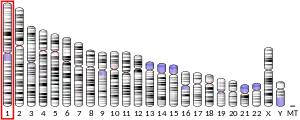
The human L-selectin gene (sell) is located on the long arm of chromosome 1 (1q24.2), and is arranged in tandem with its family members (in the order: L-, P-, and E-selectin). Human sell consists of 10 exons and its transcription factor is FOXO 1,[9] on the other hand the mouse sell gene is composed of 9 exons. [6]
Subsequent splicing of exons into mature mRNA translates to a protein product with a predicted molecular mass of 30 kDa. L-selectin varies between cell types, has ranging molecular weight from 65 kDa in lymphocytes to 100 kDa in neutrophils, and is due to cell type-specific glycosylation. Most glycoproteins undergo either N- or O-linked glycosylation, and it is very likely that the type of L-selectin glycosylation determines the specific functions of individual cells, but this has not yet been investigated in detail. [6]
L-selectin is expressed on naive T cells and is rapidly shed following T cell priming.[6] L-selectin expression is re-activated in cytotoxic T cells once they exit the lymph node. Mature central memory T cells express L-selectin while effector memory cells do not. L-selectin is also expressed by naive B cells, with the loss of L-selectin distinguishing activated B cells destined to differentiate to antibody-secreting cells
L-selectin is expressed on circulating neutrophils and is shed following neutrophil priming.[6] Expression of L-selectin in neutrophils decreases with neutrophil aging. Classical monocytes express high levels of L-selectin while in circulation. Shedding of L-selectin from monocytes occurs during trans-endothelial migration.
L-selectin expression is also observed on oocytes and early-stage embryos. Blastocysts express L-selectin following, but not prior to emergence from the zona pellucida. An increase in L-selectin expression is observed when both the blastocyst and cytotrophoblast attach to the endometrium. L-selectin expression decreases by the 17th week of pregnancy, and remains low or non-existent until term (2017).[7]
Function
Lymphocytes
L-selectin acts as a "homing receptor" for lymphocytes to enter secondary lymphoid tissues via high endothelial venules. Ligands present on endothelial cells will bind to lymphocytes expressing L-selectin, slowing lymphocyte trafficking through the blood, and facilitating entry into a secondary lymphoid organ at that point.[10] The receptor is commonly found on the cell surfaces of T cells. Naive T-lymphocytes, which have not yet encountered their specific antigen, need to enter secondary lymph nodes to encounter their antigen. Central memory T-lymphocytes, which have encountered antigen, express L-selectin to localize in secondary lymphoid organs. Here they reside ready to proliferate upon re-encountering antigen. Effector memory T-lymphocytes do not express L-selectin, as they circulate in the periphery and have immediate effector functions upon encountering antigen. High expression of L-selectin on human bone marrow progenitor cells is an early sign of cells becoming committed to lymphoid differentiation.[11]
Neutrophils and monocytes
Similar to its role in homing lymphocytes to secondary lymphoid tissues, L-selectin expressed on the surface of monocytes and neutrophils is essential for facilitating the first stage of adhesion to venule epithelial cells (known as the “rolling stage”).[6][5] Adhesion to activated epithelial cells is a critical step in the immune response as it allows these immune cells to emigrate from the bloodstream into inflamed tissue. Prolonged rolling and transmigration of neutrophils can trigger shedding of L-selectin from the neutrophil plasma membrane.[5] The membrane-bound fragment left behind following cleavage of L-selectin has also been suggested to play a critical role in the interstitial chemotaxis of neutrophils along a cytokine gradient.[6] L-selectin on neutrophils can result in its own ectodomain shedding, drived by activation of p38 MAPK followed by antibody-mediated clustering (AMC), after which L-selectin can behave as a cell adhesion molecule and signaling receptor. L-selectin shedding is not strictly consequence of neutrohpil transmigration, because it was observed that there is differences between neutrophil migration toward acute or chronic inflammation could differ in the expression and turnover of adhesion molecules.[12]
L-selectin shedding also occurs in monocytes; however, in these cells shedding is triggered only during trans-endothelial and not by earlier stages of the adhesion process.[6] The specific shedding of L-selectin from the leading migratory fronts of transmigrating monocytes suggests that this process plays a role in facilitating the directional migration of these cells (2019).[6]
Embryo
L-selectin is also present on the surface of human embryo trophoblasts prior to implantation into the uterus. Similar to its function in lymphocytes, L-selectin acts as a receptor to facilitate adhesion of the embryo to the site of invasion on the surface epithelium of the uterine endometrium. The embryo secretes human chorionic gonadotropin (hCG), which downregulates anti-adhesion factor, MUC-1, located on the uterine epithelium at the site of invasion. Removal of MUC-1 exposes the oligosaccharide ligands of the uterine epithelium, thus allowing binding by the L-selectin receptor of the trophoblast cell, followed by embryo adhesion and invasion.[13]
Clinical significance
Human immunodeficiency virus (HIV)
L-selectin expressed on CD4 T lymphocytes has been implicated in mediating adhesion and entry of HIV. L-selectin binds gp120, one of the many glycans present on the HIV envelope. This binding allows for rolling adhesion to T cells and thus facilitates the binding of HIV to its target receptors.[14] Infection of the cell triggers shedding of L-selectin. The loss of L-selectin likely aids in the release of new virus from the cell.
Abnormal pregnancy and infertility
The binding of L-selectin to its ligands plays an important role in embryo implantation during human pregnancy. Deficiency epithelial expression of L-selectin ligands has been associated with infertility, while increased expression has been implicated in ectopic pregnancies[7]
Cancer
The adhesive properties of L-selectin have been shown to contribute to cancer progression. L-selectin interactions participate in trafficking of chronic lymphocytic leukemia cells to the lymph nodes where they are able to proliferate and evolve. Additionally, L-selectin interactions may play a role in metastasis.[15]
References
- GRCh38: Ensembl release 89: ENSG00000188404 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000026581 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- Ivetic A (March 2018). "A head-to-tail view of L-selectin and its impact on neutrophil behaviour". Cell and Tissue Research. 371 (3): 437–453. doi:10.1007/s00441-017-2774-x. PMC 5820395. PMID 29353325.
- Ivetic A, Hoskins Green HL, Hart SJ (2019-05-14). "L-selectin: A Major Regulator of Leukocyte Adhesion, Migration and Signaling". Frontiers in Immunology. 10: 1068. doi:10.3389/fimmu.2019.01068. PMC 6527602. PMID 31139190.
- Feng Y, Ma X, Deng L, Yao B, Xiong Y, Wu Y, et al. (May 2017). "Role of selectins and their ligands in human implantation stage". Glycobiology. 27 (5): 385–391. doi:10.1093/glycob/cwx009. PMID 28115423.
- Tvaroška I, Selvaraj C, Koča J (June 2020). "Selectins-The Two Dr. Jekyll and Mr. Hyde Faces of Adhesion Molecules-A Review". Molecules. 25 (12): 2835. doi:10.3390/molecules25122835. PMC 7355470. PMID 32575485.
- Kerdiles YM, Beisner DR, Tinoco R, Dejean AS, Castrillon DH, DePinho RA, Hedrick SM (February 2009). "Foxo1 links homing and survival of naive T cells by regulating L-selectin, CCR7 and interleukin 7 receptor". Nature Immunology. 10 (2): 176–184. doi:10.1038/ni.1689. PMC 2856471. PMID 19136962.
- Robbins SL, Cotran RS, Kumar V, Collins T (1998). Robbins Pathologic Basis of Disease. Philadelphia: W.B Saunders Company. ISBN 0-7216-7335-X.
- Kohn LA, Hao QL, Sasidharan R, Parekh C, Ge S, Zhu Y, et al. (October 2012). "Lymphoid priming in human bone marrow begins before expression of CD10 with upregulation of L-selectin". Nature Immunology. 13 (10): 963–971. doi:10.1038/ni.2405. PMC 3448017. PMID 22941246.
- Björkman L, Christenson K, Davidsson L, Mårtensson J, Amirbeagi F, Welin A, et al. (June 2019). "Neutrophil recruitment to inflamed joints can occur without cellular priming". Journal of Leukocyte Biology. 105 (6): 1123–1130. doi:10.1002/JLB.3AB0918-369R. PMID 30570778. S2CID 58603580.
- James JL, Carter AM, Chamley LW (May 2012). "Human placentation from nidation to 5 weeks of gestation. Part I: What do we know about formative placental development following implantation?". Placenta. 33 (5): 327–334. doi:10.1016/j.placenta.2012.01.020. PMID 22374510.
- Segura J, He B, Ireland J, Zou Z, Shen T, Roth G, Sun PD (2021-09-29). "The Role of L-Selectin in HIV Infection". Frontiers in Microbiology. 12: 725741. doi:10.3389/fmicb.2021.725741. PMC 8511817. PMID 34659153.
- Natoni A, Macauley MS, O'Dwyer ME (2016). "Targeting Selectins and Their Ligands in Cancer". Frontiers in Oncology. 6: 93. doi:10.3389/fonc.2016.00093. PMC 4834419. PMID 27148485.
Further reading
- Ryan US, Worthington RE (February 1992). "Cell-cell contact mechanisms". Current Opinion in Immunology. 4 (1): 33–37. doi:10.1016/0952-7915(92)90120-4. PMID 1375831.
- Nicholson IC (2003). "CD62L (L-selectin)". Journal of Biological Regulators and Homeostatic Agents. 16 (2): 144–146. PMID 12144128.
- Ivetic A, Ridley AJ (December 2004). "The telling tail of L-selectin". Biochemical Society Transactions. 32 (Pt 6): 1118–1121. doi:10.1042/BST0321118. PMID 15506984.
- Lasky LA, Singer MS, Dowbenko D, Imai Y, Henzel WJ, Grimley C, et al. (June 1992). "An endothelial ligand for L-selectin is a novel mucin-like molecule". Cell. 69 (6): 927–938. doi:10.1016/0092-8674(92)90612-G. PMID 1376638. S2CID 9517058.
- Ord DC, Ernst TJ, Zhou LJ, Rambaldi A, Spertini O, Griffin J, Tedder TF (May 1990). "Structure of the gene encoding the human leukocyte adhesion molecule-1 (TQ1, Leu-8) of lymphocytes and neutrophils". The Journal of Biological Chemistry. 265 (14): 7760–7767. doi:10.1016/S0021-9258(19)38994-X. PMID 1692315.
- Bevilacqua M, Butcher E, Furie B, Furie B, Gallatin M, Gimbrone M, et al. (October 1991). "Selectins: a family of adhesion receptors". Cell. 67 (2): 233. doi:10.1016/0092-8674(91)90174-W. hdl:2027.42/29086. PMID 1717161. S2CID 35258400.
- Tedder TF, Isaacs CM, Ernst TJ, Demetri GD, Adler DA, Disteche CM (July 1989). "Isolation and chromosomal localization of cDNAs encoding a novel human lymphocyte cell surface molecule, LAM-1. Homology with the mouse lymphocyte homing receptor and other human adhesion proteins". The Journal of Experimental Medicine. 170 (1): 123–133. doi:10.1084/jem.170.1.123. PMC 2189363. PMID 2473156.
- Camerini D, James SP, Stamenkovic I, Seed B (November 1989). "Leu-8/TQ1 is the human equivalent of the Mel-14 lymph node homing receptor". Nature. 342 (6245): 78–82. Bibcode:1989Natur.342...78C. doi:10.1038/342078a0. PMID 2509939. S2CID 4342053.
- Bowen BR, Nguyen T, Lasky LA (July 1989). "Characterization of a human homologue of the murine peripheral lymph node homing receptor". The Journal of Cell Biology. 109 (1): 421–427. doi:10.1083/jcb.109.1.421. PMC 2115458. PMID 2663882.
- Siegelman MH, Weissman IL (July 1989). "Human homologue of mouse lymph node homing receptor: evolutionary conservation at tandem cell interaction domains". Proceedings of the National Academy of Sciences of the United States of America. 86 (14): 5562–5566. Bibcode:1989PNAS...86.5562S. doi:10.1073/pnas.86.14.5562. PMC 336895. PMID 2664786.
- Bajorath J, Aruffo A (November 1995). "A template for generation and comparison of three-dimensional selectin models". Biochemical and Biophysical Research Communications. 216 (3): 1018–1023. doi:10.1006/bbrc.1995.2722. PMID 7488174.
- Dianzani U, Bragardo M, Buonfiglio D, Redoglia V, Funaro A, Portoles P, et al. (May 1995). "Modulation of CD4 lateral interaction with lymphocyte surface molecules induced by HIV-1 gp120". European Journal of Immunology. 25 (5): 1306–1311. doi:10.1002/eji.1830250526. PMID 7539755. S2CID 37717142.
- Maruyama K, Sugano S (January 1994). "Oligo-capping: a simple method to replace the cap structure of eukaryotic mRNAs with oligoribonucleotides". Gene. 138 (1–2): 171–174. doi:10.1016/0378-1119(94)90802-8. PMID 8125298.
- Brenner B, Gulbins E, Schlottmann K, Koppenhoefer U, Busch GL, Walzog B, et al. (December 1996). "L-selectin activates the Ras pathway via the tyrosine kinase p56lck". Proceedings of the National Academy of Sciences of the United States of America. 93 (26): 15376–15381. Bibcode:1996PNAS...9315376B. doi:10.1073/pnas.93.26.15376. PMC 26412. PMID 8986819.
- Zöllner O, Lenter MC, Blanks JE, Borges E, Steegmaier M, Zerwes HG, Vestweber D (February 1997). "L-selectin from human, but not from mouse neutrophils binds directly to E-selectin". The Journal of Cell Biology. 136 (3): 707–716. doi:10.1083/jcb.136.3.707. PMC 2134294. PMID 9024699.
- Suzuki Y, Yoshitomo-Nakagawa K, Maruyama K, Suyama A, Sugano S (October 1997). "Construction and characterization of a full length-enriched and a 5'-end-enriched cDNA library". Gene. 200 (1–2): 149–156. doi:10.1016/S0378-1119(97)00411-3. PMID 9373149.
- Prakobphol A, Thomsson KA, Hansson GC, Rosen SD, Singer MS, Phillips NJ, et al. (April 1998). "Human low-molecular-weight salivary mucin expresses the sialyl lewisx determinant and has L-selectin ligand activity". Biochemistry. 37 (14): 4916–4927. doi:10.1021/bi972612a. PMID 9538010.
- Sassetti C, Tangemann K, Singer MS, Kershaw DB, Rosen SD (June 1998). "Identification of podocalyxin-like protein as a high endothelial venule ligand for L-selectin: parallels to CD34". The Journal of Experimental Medicine. 187 (12): 1965–1975. doi:10.1084/jem.187.12.1965. PMC 2212365. PMID 9625756.
- Malhotra R, Ward M, Sim RB, Bird MI (July 1999). "Identification of human complement Factor H as a ligand for L-selectin". The Biochemical Journal. 341 (1): 61–69. doi:10.1042/0264-6021:3410061. PMC 1220330. PMID 10377245.
- Bradley LM, Duncan DD, Tonkonogy S, Swain SL (September 1991). "Characterization of antigen-specific CD4+ effector T cells in vivo: immunization results in a transient population of MEL-14-, CD45RB- helper cells that secretes interleukin 2 (IL-2), IL-3, IL-4, and interferon gamma". The Journal of Experimental Medicine. 174 (3): 547–559. doi:10.1084/jem.174.3.547. PMC 2118927. PMID 1678774.
External links
- L-Selectin at the U.S. National Library of Medicine Medical Subject Headings (MeSH)
- Immunology at MCG 1/physresp
- Overview of all the structural information available in the PDB for UniProt: P14151 (L-selectin) at the PDBe-KB.
This article incorporates text from the United States National Library of Medicine, which is in the public domain.