Low-affinity nerve growth factor receptor
The p75 neurotrophin receptor (p75NTR) was first identified in 1973 as the low-affinity nerve growth factor receptor (LNGFR)[5][6] before discovery that p75NTR bound other neurotrophins equally well as nerve growth factor.[7][8] p75NTR is a neurotrophic factor receptor. Neurotrophic factor receptors bind Neurotrophins including Nerve growth factor, Neurotrophin-3, Brain-derived neurotrophic factor, and Neurotrophin-4. All neurotrophins bind to p75NTR. This also includes the immature pro-neurotrophin forms.[9][10] Neurotrophic factor receptors, including p75NTR, are responsible for ensuring a proper density to target ratio of developing neurons, refining broader maps in development into precise connections. p75NTR is involved in pathways that promote neuronal survival and neuronal death.[7]
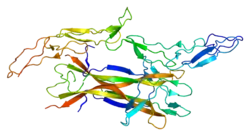


Receptor family
p75NTR is a member of the tumor necrosis factor receptor superfamily. p75NTR/LNGFR was the first member of this large family of receptors to be characterized,[5][6][11] that now contains about 25 receptors, including tumor necrosis factor 1 (TNFR1) and TNFR2, Fas, RANK, and CD40. All members of the TNFR superfamily contain structurally related cysteine-rich modules in their ECDs. p75NTR is an unusual member of this family due to its propensity to dimerize rather than trimerize, because of its ability to act as a tyrosine kinase co-receptor, and because the neurotrophins are structurally unrelated to the ligands, which typically bind TNFR family members. Indeed, with the exception of p75NTR, essentially all members of the TNFR family preferentially bind structurally related trimeric Type II transmembrane ligands, members of the TNF ligand superfamily.[12]
Structure
p75NTR is a type I transmembrane protein, with a molecular weight of 75 kDa, determined by glycosylation through both N- and O-linkages in the extracellular domain.[13] It consists of an extracellular domain, a transmembrane domain and an intracellular domain. The extracellular domain consists of a stalk domain connecting the transmembrane domain and four cysteine-rich repeat domains, CRD1, CRD2, CRD3, and CRD4; which are negatively charged, a property that facilitates Neurotrophin binding. The intracellular part is a global-like domain, known as a death domain, which consists of two sets of perpendicular helixes arranged in sets of three. It connects the transmembrane domain through a flexible linker region N-terminal domain.[14] It is important to say that, in contrast to the type I death domain found in other TNFR proteins, the type II intracellular death domain of p75NTR does nor self-associated. This was an early indication that p75NTR does nor signal death through the same mechanism as the TNFR death domains, although the ability of the p75NTR death domain to activate other second messengers is conserved.[13]
The p75ECD-binding interface to NT-3 can be divided into three main contact sites, two in the case of NGF, that are stabilized by hydrophobic interactions, salt bridges, and hydrogen bonds. The junction regions between CDR1 and CDR2 form the site 1 that contains five hydrogen bonds and one salt bridge. Site 2 is formed by equal contributions from CDR3 and CRD4 and involves two salt bridges and two hydrogen bonds. Site 3, in the CRD4, includes only one salt bridge.[15]
Function
Interactions with neurotrophins
Neurotrophins that interact with p75NTR include NGF, NT-3, BDNF, and NT-4/5.[7] Neurotrophins activating p75NTR may initiate apoptosis (for example, via c-Jun N-terminal kinases signaling, and subsequent p53, Jax-like proteins and caspase activation).[13] This effect can be counteracted by anti-apoptotic signaling by TrkA.[16] Neurotrophin binding to p75NTR, in addition to apoptotic signaling, can also promote neuronal survival (for example, via NF-kB activation).[17] There are multiple targets of Akt that could play a role in mediating p75NTR-dependent survival, but one of the more intriguing possibilities is that Ant-induced phosphorylation of IkB kinase 1 (IKK1) plays a role in the induction of NF-kB.[12]
Interactions with proneurotrophins
Proforms of NGF and BNDF (proNGF and proBNDF) are precursors to NGF and BNDF. proNGF and proBNDF interact with p75NTR and cause p75NTR-mediated apoptosis without activating TrkA-mediated survival mechanisms. Cleavage of proforms into mature Neurotrophins allows the mature NGF and BDNF to activate TrkA-mediated survival mechanisms.[18][19]
Sensory development
Recent research has suggested a number of roles for the LNGFR, including in development of the eyes and sensory neurons,[20][21] and in repair of muscle and nerve damage in adults.[22][23][24] Two distinct subpopulations of Olfactory ensheathing glia have been identified[25] with high or low cell surface expression of low-affinity nerve growth factor receptor (p75).
Interactions with other receptors
Sortilin
Sortilin is required for many apoptosis-promoting p75NTR reactions, functioning as a co-receptor for the binding of neurotrophins such as BDNF. pro-neurotrophins (such as proBDNF) bind especially well to p75NTR when sortilin is present.[26]
Crosstalk with Trk receptors
When p75NTR initiates apoptosis, NGF binding to Tropomyosin receptor kinase A (TrkA) can negate p75NTR apoptotic effects. p75NTR c-Jun kinase pathway activation (which causes apoptosis) is suppressed when NGF binds to TrkA. p75NTR activation of NF-kB, which promotes survival, is unaffected by NGF binding to TrkA.[26]
Nogo-66 receptor (NgR1)
p75NTR functions in a complex with Nogo-66 receptor (NgR1) to mediate RhoA-dependent inhibition of growth of regenerating axons exposed to inhibitory proteins of CNS myelin, such as Nogo, MAG or OMgP. Without p75NTR, OMgP can activate RhoA and inhibit CNS axon regeneration. Coexpression of p75NTR and OMgP suppress RhoA activation. A complex of NgR1, p75NTR and LINGO1 can activate RhoA.[27]
p75NTR-mediated signaling pathways
NF-kB activation
NF-kB is a transcription factor that can be activated by p75NTR. Nerve growth factor (NGF) is a neurotrophin that promotes neuronal growth, and, in the absence of NGF, neurons die. Neuronal death in the absence of NGF can be prevented by NF-kB activation. Phosphorylated IκB kinase binds to and activates NF-kB before separating from NF-kB. After separation, IκB degrades and NF-kB continues to the nucleus to initiate pro-survival transcription. NF-kB also promotes neuronal survival in conjunction with NGF.[17]
NF-kB activity is activated by p75NTR, and is not activated via Trk receptors. NF-kB activity does not effect Brain-derived neurotrophic factor promotion of neuronal survival.[17]
RhoGDI and RhoA
p75NTR serves as a regulator for actin assembly. Ras homolog family member A (RhoA) causes the actin cytoskeleton to become rigid which limits growth cone mobility and inhibits neuronal elongation in the developing nervous system. p75NTR without a ligand bound activates RhoA and limits actin assembly, but neurotrophin binding to p75NTR can inactivate RhoA and promote actin assembly.[28] p75NTR associates with the Rho GDP dissociation inhibitor (RhoGDI), and RhoGDI associates with RhoA. Interactions with Nogo can strengthen the association between p75NTR and RhoGDI. Neurotrophin binding to p75NTR inhibits the association of RhoGDI and p75NTR, thereby suppressing RhoA release and promoting growth cone elongation (inhibiting RhoA actin suppression).[29]
JNK signaling pathway
Neurotrophin binding to p75NTR activates the c-Jun N-terminal kinases (JNK) signaling pathway causing apoptosis of developing neurons. JNK, through a series of intermediates, activates p53 and p53 activates Bax which initiates apoptosis. TrkA can prevent p75NTR-mediated JNK pathway apoptosis.[30]
JNK-Bim-EL signaling pathway
JNK can directly phosphorylate Bim-EL, a splicing isoform of Bcl-2 interacting mediator of cell death (Bim), which activates Bim-EL apoptotic activity. JNK activation is required for apoptosis but c-jun, a protein in the JNK signaling pathway, is not always required.[16]
Caspase-dependent signaling
LNGFR also activates a caspase-dependent signaling pathway that promotes developmental axon pruning, and axon degeneration in neurodegenerative disease.[31]
In the apoptosis pathway, members of the TNF receptor superfamily assemble a death-inducing signaling complex (DISC) in which TRADD or FADD bind directly to the receptor's death domain, thereby allowing aggregation and activation of Caspase 8 and subsequent activation of the Caspase cascade. However, Caspase 8 induction does not appear to be involved in p75NTR-mediated apoptosis, but Caspase 9 is activated during p75NTR-mediated killing.[12]
Role in disease
Huntington's disease
Huntington's disease is characterized by cognitive impairments. There is increased expression of p75NTR in the hippocampus of Huntington's disease patients (including mice models and humans). Over expression of p75NTR in mice causes cognitive impairments similar to Huntington's disease. p75NTR is linked to reduced numbers of dendritic spines in the hippocampus, likely through p75NTR interactions with Transforming protein RhoA. Modulating p75NTR function could be a future direction in treating Huntington's disease.[32]
Amyotrophic lateral sclerosis
Amyotrophic lateral sclerosis ALS is a neurodegenerative disease characterized by progressive muscular paralysis reflecting degeneration of motor neurons in the primary motor cortex, corticospinal tracts, brainstem and spinal cord. One study using the superoxide dismutase 1 (SOD1) mutant mouse, an ALS model which develops severe neurodegeneration, the expression of p75NTR correlated with the extent of degeneration and p75NTR knockdown delayed disease progression.[33][34][35]
Alzheimer's disease
Alzheimer's disease (AD) is the most common cause of dementia in the elderly. AD is a neurodegenerative disease characterized by the loss of cognitive functioning - thinking, remembering and reasoning- and behavioral abilities to such an extent that it interferes with a person's daily life and activities. The neuropathological hallmarks of AD include amyloid plaques and neurofibrillary tangles, which lead to neuronal death. Studies in animal models of AD have shown that p75NTR contributes to amyloid β-induced neuronal damage.[36] In humans with AD, increases in p75NTR expression relative to TrkA have been suggested to be responsible for the loss of cholinergic neurons.[37][38] Increases in proNGF in AD [39] indicate that the Neurotrophin environment is favorable for p75NTR/sortilin signaling and supports the theory that age-related neural damage is facilitated by a shift toward proNGF-mediated signaling.[35] A recent study found that activation of Ngfr signaling in astroglia of Alzheimer's disease mouse model enhanced neurogenesis and reduced two hallmarks of Alzheimer's disease.[40] This study also found that NGFR signaling in humans is age-related and correlates with proliferative potential of neural progenitors.
Role in cancer stem cells
p75NTR has been implicated as a marker for cancer stem cells in melanoma and other cancers. Melanoma cells transplanted into an immunodeficient mouse model were shown to require expression of CD271 in order to grow a melanoma.[41] Gene knockdown of CD271 has also been shown to abolish neural crest stem cell properties of melanoma cells and decrease genomic stability leading to a reduced migration, tumorigenicity, proliferation and induction of apoptosis.[42][43][44] Furthermore, increased levels of CD271 were observed in brain metastatic melanoma cells whereas resistance to the BRAF inhibitor vemurafenib supposedly selects for highly malignant brain and lung-metastasizing melanoma cells.[45][44][46][47] Recently, expression of p75NTR (NGFR) was associated with progressive intracranial disease in melanoma patients [48]
Interactions
Low-affinity nerve growth factor receptor has been shown to interact with:
References
- GRCh38: Ensembl release 89: ENSG00000064300 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000000120 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- Johnson D, Lanahan A, Buck CR, Sehgal A, Morgan C, Mercer E, et al. (November 1986). "Expression and structure of the human NGF receptor". Cell. 47 (4): 545–554. doi:10.1016/0092-8674(86)90619-7. PMID 3022937. S2CID 22472119.
- Radeke MJ, Misko TP, Hsu C, Herzenberg LA, Shooter EM (1987). "Gene transfer and molecular cloning of the rat nerve growth factor receptor". Nature. 325 (6105): 593–597. Bibcode:1987Natur.325..593R. doi:10.1038/325593a0. PMID 3027580. S2CID 4342838.
- Bibel M, Hoppe E, Barde YA (February 1999). "Biochemical and functional interactions between the neurotrophin receptors trk and p75NTR". The EMBO Journal. 18 (3): 616–622. doi:10.1093/emboj/18.3.616. PMC 1171154. PMID 9927421.
- Frade JM, Barde YA (February 1998). "Nerve growth factor: two receptors, multiple functions". BioEssays. 20 (2): 137–145. doi:10.1002/(SICI)1521-1878(199802)20:2<137::AID-BIES6>3.0.CO;2-Q. PMID 9631659. S2CID 38060583.
- Barker PA (May 2004). "p75NTR is positively promiscuous: novel partners and new insights". Neuron. 42 (4): 529–533. doi:10.1016/j.neuron.2004.04.001. PMID 15157416. S2CID 15080734.
- Chao MV, Bothwell M (January 2002). "Neurotrophins: to cleave or not to cleave". Neuron. 33 (1): 9–12. doi:10.1016/s0896-6273(01)00573-6. PMID 11779474. S2CID 17364992.
- He XL, Garcia KC (May 2004). "Structure of nerve growth factor complexed with the shared neurotrophin receptor p75". Science. 304 (5672): 870–875. Bibcode:2004Sci...304..870H. doi:10.1126/science.1095190. PMID 15131306. S2CID 46060440.
- Roux PP, Barker PA (June 2002). "Neurotrophin signaling through the p75 neurotrophin receptor". Progress in Neurobiology. 67 (3): 203–233. doi:10.1016/S0301-0082(02)00016-3. PMID 12169297. S2CID 8659757.
- Underwood CK, Coulson EJ (30 June 2007). "The p75 neurotrophin receptor". The International Journal of Biochemistry & Cell Biology. 40 (9): 1664–1668. doi:10.1016/j.biocel.2007.06.010. PMID 17681869.
- Chen Y, Zeng J, Cen L, Chen Y, Wang X, Yao G, et al. (2009). "Multiple roles of the p75 neurotrophin receptor in the nervous system". The Journal of International Medical Research. 37 (2): 281–288. doi:10.1177/147323000903700201. PMID 19383220. S2CID 40720230.
- Vilar M (29 November 2016). "Structural Characterization of the p75 Neurotrophin Receptor: A Stranger in the TNFR Superfamily". Vitamins and Hormones. 104: 57–87. doi:10.1016/bs.vh.2016.10.007. PMID 28215307.
- Becker EB, Howell J, Kodama Y, Barker PA, Bonni A (October 2004). "Characterization of the c-Jun N-terminal kinase-BimEL signaling pathway in neuronal apoptosis". The Journal of Neuroscience. 24 (40): 8762–8770. doi:10.1523/JNEUROSCI.2953-04.2004. PMC 6729963. PMID 15470142.
- Hamanoue M, Middleton G, Wyatt S, Jaffray E, Hay RT, Davies AM (July 1999). "p75-mediated NF-kappaB activation enhances the survival response of developing sensory neurons to nerve growth factor". Molecular and Cellular Neurosciences. 14 (1): 28–40. doi:10.1006/mcne.1999.0770. PMID 10433815. S2CID 25648122.
- Lee R, Kermani P, Teng KK, Hempstead BL (November 2001). "Regulation of cell survival by secreted proneurotrophins". Science. 294 (5548): 1945–1948. Bibcode:2001Sci...294.1945L. doi:10.1126/science.1065057. PMID 11729324. S2CID 872149.
- Feng D, Kim T, Ozkan E, Light M, Torkin R, Teng KK, et al. (March 2010). "Molecular and structural insight into proNGF engagement of p75NTR and sortilin". Journal of Molecular Biology. 396 (4): 967–984. doi:10.1016/j.jmb.2009.12.030. PMC 2847487. PMID 20036257.
- Nicol GD (October 2008). "Nerve growth factor, sphingomyelins, and sensitization in sensory neurons". Sheng Li Xue Bao. 60 (5): 603–604. PMID 18958367.
- Di Girolamo N, Sarris M, Chui J, Cheema H, Coroneo MT, Wakefield D (December 2008). "Localization of the low-affinity nerve growth factor receptor p75 in human limbal epithelial cells". Journal of Cellular and Molecular Medicine. 12 (6B): 2799–2811. doi:10.1111/j.1582-4934.2008.00290.x. PMC 3828893. PMID 19210757.
- Chen LW, Yung KK, Chan YS, Shum DK, Bolam JP (December 2008). "The proNGF-p75NTR-sortilin signalling complex as new target for the therapeutic treatment of Parkinson's disease". CNS & Neurological Disorders Drug Targets. 7 (6): 512–523. doi:10.2174/187152708787122923. PMID 19128208.
- Kocsis JD, Lankford KL, Sasaki M, Radtke C (June 2009). "Unique in vivo properties of olfactory ensheathing cells that may contribute to neural repair and protection following spinal cord injury". Neuroscience Letters. 456 (3): 137–142. doi:10.1016/j.neulet.2008.08.093. PMC 2713444. PMID 19429149.
- Deponti D, Buono R, Catanzaro G, De Palma C, Longhi R, Meneveri R, et al. (August 2009). "The low-affinity receptor for neurotrophins p75NTR plays a key role for satellite cell function in muscle repair acting via RhoA". Molecular Biology of the Cell. 20 (16): 3620–3627. doi:10.1091/mbc.E09-01-0012. PMC 2777922. PMID 19553472.
- Honoré A, Le Corre S, Derambure C, Normand R, Duclos C, Boyer O, et al. (March 2012). "Isolation, characterization, and genetic profiling of subpopulations of olfactory ensheathing cells from the olfactory bulb". Glia. 60 (3): 404–413. doi:10.1002/glia.22274. PMID 22161947. S2CID 31230806.
- Chen ZY, Ieraci A, Teng H, Dall H, Meng CX, Herrera DG, et al. (June 2005). "Sortilin controls intracellular sorting of brain-derived neurotrophic factor to the regulated secretory pathway". The Journal of Neuroscience. 25 (26): 6156–6166. doi:10.1523/JNEUROSCI.1017-05.2005. PMC 1201519. PMID 15987945.
- Mi S, Lee X, Shao Z, Thill G, Ji B, Relton J, et al. (March 2004). "LINGO-1 is a component of the Nogo-66 receptor/p75 signaling complex". Nature Neuroscience. 7 (3): 221–228. doi:10.1038/nn1188. PMID 14966521. S2CID 2344794.
- Yamashita T, Tucker KL, Barde YA (November 1999). "Neurotrophin binding to the p75 receptor modulates Rho activity and axonal outgrowth". Neuron. 24 (3): 585–593. doi:10.1016/s0896-6273(00)81114-9. PMID 10595511. S2CID 17271817.
- Yamashita T, Tohyama M (May 2003). "The p75 receptor acts as a displacement factor that releases Rho from Rho-GDI". Nature Neuroscience. 6 (5): 461–467. doi:10.1038/nn1045. PMID 12692556. S2CID 10865814.
- Aloyz RS, Bamji SX, Pozniak CD, Toma JG, Atwal J, Kaplan DR, Miller FD (December 1998). "p53 is essential for developmental neuron death as regulated by the TrkA and p75 neurotrophin receptors". The Journal of Cell Biology. 143 (6): 1691–1703. doi:10.1083/jcb.143.6.1691. PMC 2132983. PMID 9852160.
- Simon DJ, Weimer RM, McLaughlin T, Kallop D, Stanger K, Yang J, et al. (December 2012). "A caspase cascade regulating developmental axon degeneration". The Journal of Neuroscience. 32 (49): 17540–17553. doi:10.1523/JNEUROSCI.3012-12.2012. PMC 3532512. PMID 23223278.
- Brito V, Giralt A, Enriquez-Barreto L, Puigdellívol M, Suelves N, Zamora-Moratalla A, et al. (October 2014). "Neurotrophin receptor p75(NTR) mediates Huntington's disease-associated synaptic and memory dysfunction". The Journal of Clinical Investigation. 124 (10): 4411–4428. doi:10.1172/JCI74809. PMC 4191006. PMID 25180603.
- Shepheard SR, Chataway T, Schultz DW, Rush RA, Rogers ML (January 2014). "The extracellular domain of neurotrophin receptor p75 as a candidate biomarker for amyotrophic lateral sclerosis". PLOS ONE. 9 (1): e87398. Bibcode:2014PLoSO...987398S. doi:10.1371/journal.pone.0087398. PMC 3903651. PMID 24475283.
- Turner BJ, Cheah IK, Macfarlane KJ, Lopes EC, Petratos S, Langford SJ, Cheema SS (November 2003). "Antisense peptide nucleic acid-mediated knockdown of the p75 neurotrophin receptor delays motor neuron disease in mutant SOD1 transgenic mice". Journal of Neurochemistry. 87 (3): 752–763. doi:10.1046/j.1471-4159.2003.02053.x. PMID 14535957. S2CID 5901529.
- Meeker RB, Williams KS (May 2015). "The p75 neurotrophin receptor: at the crossroad of neural repair and death". Neural Regeneration Research. 10 (5): 721–725. doi:10.4103/1673-5374.156967. PMC 4468762. PMID 26109945.
- Yang T, Knowles JK, Lu Q, Zhang H, Arancio O, Moore LA, et al. (21 July 2008). "Small molecule, non-peptide p75 ligands inhibit Abeta-induced neurodegeneration and synaptic impairment". PLOS ONE. 3 (11): e3604. Bibcode:2008PLoSO...3.3604Y. doi:10.1371/journal.pone.0003604. PMC 2575383. PMID 18978948.
- Costantini C, Scrable H, Puglielli L (May 2006). "An aging pathway controls the TrkA to p75NTR receptor switch and amyloid beta-peptide generation". The EMBO Journal. 25 (9): 1997–2006. doi:10.1038/sj.emboj.7601062. PMC 1456930. PMID 16619032.
- Counts SE, Nadeem M, Wuu J, Ginsberg SD, Saragovi HU, Mufson EJ (October 2004). "Reduction of cortical TrkA but not p75(NTR) protein in early-stage Alzheimer's disease". Annals of Neurology. 56 (4): 520–531. doi:10.1002/ana.20233. PMID 15455399. S2CID 38106502.
- Fahnestock M, Michalski B, Xu B, Coughlin MD (August 2001). "The precursor pro-nerve growth factor is the predominant form of nerve growth factor in brain and is increased in Alzheimer's disease". Molecular and Cellular Neurosciences. 18 (2): 210–220. doi:10.1006/mcne.2001.1016. PMID 11520181. S2CID 8443739.
- Siddiqui T, Cosacak MI, Popova S, Bhattarai P, Yilmaz E, Lee AJ, Min Y, Wang X, Allen M, İş O, Atasavum ZT, Rodriguez-Muela N, Vardarajan BN, Flaherty D, Teich AF, Santa-Maria I, Freudenberg U, Werner C, Tosto G, Mayeux R, Ertekin-Taner N, Kizil C (July 2023). "Nerve growth factor receptor (Ngfr) induces neurogenic plasticity by suppressing reactive astroglial Lcn2/Slc22a17 signaling in Alzheimer's disease". npj Regenerative Medicine. 8 (1): 33. doi:10.1038/s41536-023-00311-5. PMC 10333226. PMID 37429840.
- name="pmid20596026">Boiko AD, Razorenova OV, van de Rijn M, Swetter SM, Johnson DL, Ly DP, et al. (July 2010). "Human melanoma-initiating cells express neural crest nerve growth factor receptor CD271". Nature. 466 (7302): 133–137. Bibcode:2010Natur.466..133B. doi:10.1038/nature09161. PMC 2898751. PMID 20596026.
- Redmer T, Welte Y, Behrens D, Fichtner I, Przybilla D, Wruck W, et al. (2014). "The nerve growth factor receptor CD271 is crucial to maintain tumorigenicity and stem-like properties of melanoma cells". PLOS ONE. 9 (5): e92596. Bibcode:2014PLoSO...992596R. doi:10.1371/journal.pone.0092596. PMC 4010406. PMID 24799129.
- Redmer T, Walz I, Klinger B, Khouja S, Welte Y, Schäfer R, Regenbrecht C (January 2017). "The role of the cancer stem cell marker CD271 in DNA damage response and drug resistance of melanoma cells". Oncogenesis. 6 (1): e291. doi:10.1038/oncsis.2016.88. PMC 5294251. PMID 28112719.
- Radke J, Roßner F, Redmer T (August 2017). "CD271 determines migratory properties of melanoma cells". Scientific Reports. 7 (1): 9834. Bibcode:2017NatSR...7.9834R. doi:10.1038/s41598-017-10129-z. PMC 5574914. PMID 28852061.
- Guo R, Fierro-Fine A, Goddard L, Russell M, Chen J, Liu CZ, et al. (2014). "Increased expression of melanoma stem cell marker CD271 in metastatic melanoma to the brain". International Journal of Clinical and Experimental Pathology. 7 (12): 8947–8951. PMC 4313973. PMID 25674270.
- Zubrilov I, Sagi-Assif O, Izraely S, Meshel T, Ben-Menahem S, Ginat R, et al. (May 2015). "Vemurafenib resistance selects for highly malignant brain and lung-metastasizing melanoma cells". Cancer Letters. 361 (1): 86–96. doi:10.1016/j.canlet.2015.02.041. PMID 25725450.
- Redmer T (July 2018). "Deciphering mechanisms of brain metastasis in melanoma - the gist of the matter". Molecular Cancer. 17 (1): 106. doi:10.1186/s12943-018-0854-5. PMC 6064184. PMID 30053879.
- Radke J, Schumann E, Onken J, Koll R, Acker G, Bodnar B, et al. (November 2022). "Decoding molecular programs in melanoma brain metastases". Nature Communications. 13 (1): 7304. Bibcode:2022NatCo..13.7304R. doi:10.1038/s41467-022-34899-x. PMC 9701224. PMID 36435874.
- Shonukan O, Bagayogo I, McCrea P, Chao M, Hempstead B (June 2003). "Neurotrophin-induced melanoma cell migration is mediated through the actin-bundling protein fascin". Oncogene. 22 (23): 3616–3623. doi:10.1038/sj.onc.1206561. PMID 12789270.
- Tcherpakov M, Bronfman FC, Conticello SG, Vaskovsky A, Levy Z, Niinobe M, et al. (December 2002). "The p75 neurotrophin receptor interacts with multiple MAGE proteins". The Journal of Biological Chemistry. 277 (51): 49101–49104. doi:10.1074/jbc.C200533200. PMID 12414813.
- Kuwako K, Taniura H, Yoshikawa K (January 2004). "Necdin-related MAGE proteins differentially interact with the E2F1 transcription factor and the p75 neurotrophin receptor". The Journal of Biological Chemistry. 279 (3): 1703–1712. doi:10.1074/jbc.M308454200. PMID 14593116.
- Bronfman FC, Tcherpakov M, Jovin TM, Fainzilber M (April 2003). "Ligand-induced internalization of the p75 neurotrophin receptor: a slow route to the signaling endosome". The Journal of Neuroscience. 23 (8): 3209–3220. doi:10.1523/JNEUROSCI.23-08-03209.2003. PMC 6742322. PMID 12716928.
- Mukai J, Hachiya T, Shoji-Hoshino S, Kimura MT, Nadano D, Suvanto P, et al. (June 2000). "NADE, a p75NTR-associated cell death executor, is involved in signal transduction mediated by the common neurotrophin receptor p75NTR". The Journal of Biological Chemistry. 275 (23): 17566–17570. doi:10.1074/jbc.C000140200. PMID 10764727.
- Nykjaer A, Lee R, Teng KK, Jansen P, Madsen P, Nielsen MS, et al. (February 2004). "Sortilin is essential for proNGF-induced neuronal cell death". Nature. 427 (6977): 843–848. Bibcode:2004Natur.427..843N. doi:10.1038/nature02319. PMID 14985763. S2CID 4343450.
- Higuchi H, Yamashita T, Yoshikawa H, Tohyama M (April 2003). "PKA phosphorylates the p75 receptor and regulates its localization to lipid rafts". The EMBO Journal. 22 (8): 1790–1800. doi:10.1093/emboj/cdg177. PMC 154469. PMID 12682012.
- Ye X, Mehlen P, Rabizadeh S, VanArsdale T, Zhang H, Shin H, et al. (October 1999). "TRAF family proteins interact with the common neurotrophin receptor and modulate apoptosis induction". The Journal of Biological Chemistry. 274 (42): 30202–30208. doi:10.1074/jbc.274.42.30202. PMID 10514511.
- Krajewska M, Krajewski S, Zapata JM, Van Arsdale T, Gascoyne RD, Berern K, et al. (June 1998). "TRAF-4 expression in epithelial progenitor cells. Analysis in normal adult, fetal, and tumor tissues". The American Journal of Pathology. 152 (6): 1549–1561. PMC 1858434. PMID 9626059.
Further reading
- Buxser S, Puma P, Johnson GL (February 1985). "Properties of the nerve growth factor receptor. Relationship between receptor structure and affinity". The Journal of Biological Chemistry. 260 (3): 1917–1926. doi:10.1016/S0021-9258(18)89680-6. PMID 2981877.
- Glass DJ, Nye SH, Hantzopoulos P, Macchi MJ, Squinto SP, Goldfarb M, Yancopoulos GD (July 1991). "TrkB mediates BDNF/NT-3-dependent survival and proliferation in fibroblasts lacking the low affinity NGF receptor". Cell. 66 (2): 405–413. doi:10.1016/0092-8674(91)90629-D. PMID 1649703. S2CID 43626580.
- Ibáñez CF (June 2002). "Jekyll-Hyde neurotrophins: the story of proNGF". Trends in Neurosciences. 25 (6): 284–286. doi:10.1016/S0166-2236(02)02169-0. PMID 12086739. S2CID 9449831.
- Radeke MJ, Misko TP, Hsu C, Herzenberg LA, Shooter EM (1987). "Gene transfer and molecular cloning of the rat nerve growth factor receptor". Nature. 325 (6105): 593–597. Bibcode:1987Natur.325..593R. doi:10.1038/325593a0. PMID 3027580. S2CID 4342838.
External links
- Nerve+Growth+Factor+Receptor,+Low-Affinity at the U.S. National Library of Medicine Medical Subject Headings (MeSH)
PDB gallery | |
|---|---|
|
