CD22
CD22, or cluster of differentiation-22, is a molecule belonging to the SIGLEC family of lectins.[4] It is found on the surface of mature B cells and to a lesser extent on some immature B cells. Generally speaking, CD22 is a regulatory molecule that prevents the overactivation of the immune system and the development of autoimmune diseases.[5]
| CD22 | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Identifiers | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Aliases | CD22, SIGLEC-2, SIGLEC2, CD22 molecule | ||||||||||||||||||||||||||||||||||||||||||||||||||
| External IDs | OMIM: 107266 MGI: 88322 HomoloGene: 31052 GeneCards: CD22 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Wikidata | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||


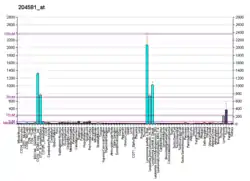
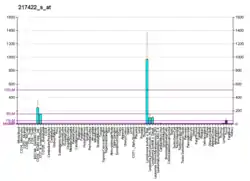
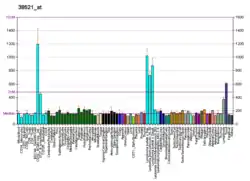
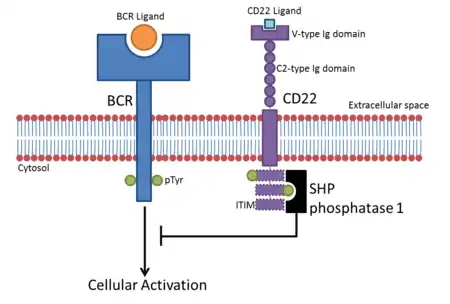
CD22 is a sugar binding transmembrane protein, which specifically binds sialic acid with an immunoglobulin (Ig) domain located at its N-terminus. The presence of Ig domains makes CD22 a member of the immunoglobulin superfamily. CD22 functions as an inhibitory receptor for B cell receptor (BCR) signaling. It is also involved in the B cell trafficking to Peyer's patches in mice.[6] In mice, it has been shown that CD22 blockade restores homeostatic microglial phagocytosis in aging brains.[7]
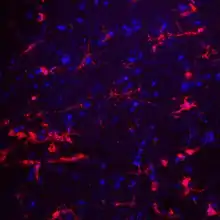
Structure
CD22 is a transmembrane protein with a molecular weight of 140 kDa. The extracellular part of CD22 consists of seven immunoglobulin domains and the intracellular part is formed by 141-amino acid cytoplasmic tail.[8]
Extracellular part
The binding site for ligands is located at the extracellular N-terminus, specifically at the last immunoglobulin domain called the V-like domain. This domain binds to ligands containing sialic acid via α2,6-linkage to the galactose residue. Such ligands are commonly expressed on the surface of erythrocytes, monocytes, cytokine-activated endothelial cells, T cells and B cells. To a lesser extent they are present on soluble IgM and on the soluble plasmatic glycoprotein called haptoglobin. Therefore, CD22 can bind ligands in the cis configuration, when they are on the surface of B cells, or in the trans configuration, when they are on the surface of other cells or on soluble glycoproteins or attached to a cell-associated antigen. However, CD22 is masked on most B-cell surfaces, meaning that it cannot bind exogenous ligands, so cis interaction with glycoprotein ligands on the same cell is preferred.[9]
Trans ligands
Trans interactions between CD22 and its ligands are important for B cell adhesion and migration. Specifically, CD22-deficient mice have been shown to have reduced numbers of recirculating B cells and reduced numbers of IgM-secreting plasma cells in the bone marrow. Together, this implies that CD22 interacting with trans ligands is crucial for the homing of mature, recirculating B cells to the bone marrow.[10]
BCR signaling
The intracellular part of CD22 consists of 6 tyrosine residues which contain both ITIM and ITAM motifs suggesting both inhibitory and activation role in signaling.[11] Because of the tyrosine residues, the cytoplasmic domain of CD22 can be phosphorylated. This happens when the BCR is cross-linked by the antigen. Phosphorylation is mediated by Lyn, a protein tyrosine kinase (PTK) of the Src family found in lipid rafts.[9]
Inhibitory role
After CD22 is phosphorylated, the ITIM motifs provide docking sites for the SH2 domain containing protein tyrosine phosphatase called SHP-1. SHP-1 inhibits mitogen-activated protein kinase (MAPK) and dephosphorylates components of BCR signaling. That means that association of CD22 with SHP-1 leads to the inhibition of BCR signaling.[12][9]
Activation role
After CD22 is phosphorylated, the ITAM motifs provide docking sites for the SH2 domain of Lyn or other Syk kinase or Src-family tyrosine kinases. Thus, CD22 positively regulates BCR signaling and thereby promotes B cell survival.[9]
Autoimmunity
Single-nucleotide polymorphisms in the CD22 gene lead to a higher likelihood of autoimmune disease. Specifically, some studies show that polymorphisms in the CD22 gene are associated with susceptibility to systemic lupus erythematosus (SLE) and cutaneous systemic sclerosis. In addition, mutations in enzymes involved in the glycosylation of the CD22 ligand may also lead to the susceptibility to autoimmune diseases. Specifically, mutations in the sialic acid esterase were frequently found in patients with rheumatoid arthritis and SLE. This enzyme is essential for deacetylation of the N-glycan sialic acid present in CD22 ligands and is therefore crucial for ligand binding.[13]
As a drug target
Because CD22 is restricted to B cells, it is an excellent target for immunotherapy of B cell malignancies. There are several mechanisms by which this can be achieved, namely monoclonal antibodies, bispecific antibodies, antibody-drug conjugates, radioimmunoconjugates or CAR-T cells.[14]
An immunotoxin, BL22 (CAT-3888), that targets this receptor was developed at the NIH.[15] BL22 was superseded by moxetumomab pasudotox (HA22, CAT-8015).[16] Moxetumomab pasudotox is approved in the EU and USA for treatment of relapsed or refractory hairy cell leukemia.[17][18]
Inotuzumab
It was shown that antibody-drug conjugates work better than naked antibodies. The reason is that CD22 is rapidly internalized rather than being exposed to the extracellular environment making it more suitable for specific delivery of these conjugates.[19] One of such therapeutics is Inotuzumab, which was approved by the FDA for the treatment of relapsed or refractory B cell acute lymphoblastic leukemia in August 2017.[20] Inotuzumab consists of a CD22-targeting immunoglobulin G4 humanized monoclonal antibody conjugated to calicheamicin. The mechanism by which calicheamicin destroys malignant cells is that it binds to DNA, causing DNA double-strand breaks, and this in turn leads to transcription inhibition.[19]
Interactions
CD22 has been shown to interact with Grb2,[21][22] PTPN6,[22][23][24][25][26] LYN,[21][24] SHC1[21] and INPP5D.[21]
References
- GRCm38: Ensembl release 89: ENSMUSG00000030577 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- Crocker PR, Clark EA, Filbin M, Gordon S, Jones Y, Kehrl JH, et al. (February 1998). "Siglecs: a family of sialic-acid binding lectins". Glycobiology. 8 (2): v. doi:10.1093/glycob/8.2.0. PMID 9498912.
- Hatta Y, Tsuchiya N, Matsushita M, Shiota M, Hagiwara K, Tokunaga K (April 1999). "Identification of the gene variations in human CD22". Immunogenetics. 49 (4): 280–6. doi:10.1007/s002510050494. PMID 10079291. S2CID 22947237.
- Lee M, Kiefel H, LaJevic MD, Macauley MS, Kawashima H, O'Hara E, et al. (October 2014). "Transcriptional programs of lymphoid tissue capillary and high endothelium reveal control mechanisms for lymphocyte homing". Nature Immunology. 15 (10): 982–95. doi:10.1038/ni.2983. PMC 4222088. PMID 25173345.
- Pluvinage JV, Wyss-Coray T, et al. (April 11, 2019). "CD22 blockade restores homeostatic microglial phagocytosis in aging brains". Nature. 568 (7751): 187–192. Bibcode:2019Natur.568..187P. doi:10.1038/s41586-019-1088-4. PMC 6574119. PMID 30944478.
- Tedder, T. F.; Tuscano, J.; Sato, S.; Kehrl, J. H. (1997). "CD22, a B lymphocyte-specific adhesion molecule that regulates antigen receptor signaling". Annual Review of Immunology. 15: 481–504. doi:10.1146/annurev.immunol.15.1.481. ISSN 0732-0582. PMID 9143697.
- Walker, Jennifer A.; Smith, Kenneth G. C. (March 2008). "CD22: an inhibitory enigma". Immunology. 123 (3): 314–325. doi:10.1111/j.1365-2567.2007.02752.x. ISSN 1365-2567. PMC 2433339. PMID 18067554.
- Nitschke, Lars (July 2009). "CD22 and Siglec-G: B-cell inhibitory receptors with distinct functions". Immunological Reviews. 230 (1): 128–143. doi:10.1111/j.1600-065X.2009.00801.x. ISSN 1600-065X. PMID 19594633. S2CID 205825220.
- Poe, J. C.; Fujimoto, M.; Jansen, P. J.; Miller, A. S.; Tedder, T. F. (2000-06-09). "CD22 forms a quaternary complex with SHIP, Grb2, and Shc. A pathway for regulation of B lymphocyte antigen receptor-induced calcium flux". The Journal of Biological Chemistry. 275 (23): 17420–17427. doi:10.1074/jbc.M001892200. ISSN 0021-9258. PMID 10748054.
- Sato, S.; Tuscano, J. M.; Inaoki, M.; Tedder, T. F. (August 1998). "CD22 negatively and positively regulates signal transduction through the B lymphocyte antigen receptor". Seminars in Immunology. 10 (4): 287–297. doi:10.1006/smim.1998.0121. ISSN 1044-5323. PMID 9695185.
- Clark, Edward A.; Giltiay, Natalia V. (2018). "CD22: A Regulator of Innate and Adaptive B Cell Responses and Autoimmunity". Frontiers in Immunology. 9: 2235. doi:10.3389/fimmu.2018.02235. ISSN 1664-3224. PMC 6173129. PMID 30323814.
- Shah, Nikesh N.; Sokol, Lubomir (2021). "Targeting CD22 for the Treatment of B-Cell Malignancies". ImmunoTargets and Therapy. 10: 225–236. doi:10.2147/ITT.S288546. ISSN 2253-1556. PMC 8275043. PMID 34262884.
- Clinical trial number NCT00074048 for "BL22 Immunotoxin in Treating Patients Previously Treated With Cladribine for Hairy Cell Leukemia" at ClinicalTrials.gov
- http://www.cambridgeantibody.com/__data/assets/pdf_file/10857/CAT-3888,_CAT-8015_and_CAT-5001_Nov06.pdf Archived 2007-02-27 at the Wayback Machine CAT URL Redirects to Medimmune home page
- "Lumoxiti EPAR". European Medicines Agency (EMA). 9 December 2020. Retrieved 16 July 2021..
- "Moxetumomab pasudotox-tdfk FDA Approval". U.S. Food and Drug Administration (FDA). Retrieved 20 April 2020.
- Wynne, Joseph; Wright, David; Stock, Wendy (2019-01-08). "Inotuzumab: from preclinical development to success in B-cell acute lymphoblastic leukemia". Blood Advances. 3 (1): 96–104. doi:10.1182/bloodadvances.2018026211. ISSN 2473-9537. PMC 6325303. PMID 30622147.
- Research, Center for Drug Evaluation and (2019-02-09). "FDA approves inotuzumab ozogamicin for relapsed or refractory B-cell precursor ALL". FDA.
- Poe JC, Fujimoto M, Jansen PJ, Miller AS, Tedder TF (June 2000). "CD22 forms a quaternary complex with SHIP, Grb2, and Shc. A pathway for regulation of B lymphocyte antigen receptor-induced calcium flux". The Journal of Biological Chemistry. 275 (23): 17420–7. doi:10.1074/jbc.M001892200. PMID 10748054.
- Otipoby KL, Draves KE, Clark EA (November 2001). "CD22 regulates B cell receptor-mediated signals via two domains that independently recruit Grb2 and SHP-1". The Journal of Biological Chemistry. 276 (47): 44315–22. doi:10.1074/jbc.M105446200. PMID 11551923.
- Blasioli J, Paust S, Thomas ML (January 1999). "Definition of the sites of interaction between the protein tyrosine phosphatase SHP-1 and CD22". The Journal of Biological Chemistry. 274 (4): 2303–7. doi:10.1074/jbc.274.4.2303. PMID 9890995.
- Greer SF, Justement LB (May 1999). "CD45 regulates tyrosine phosphorylation of CD22 and its association with the protein tyrosine phosphatase SHP-1". Journal of Immunology. 162 (9): 5278–86. doi:10.4049/jimmunol.162.9.5278. PMID 10228003. S2CID 2223820.
- Law CL, Sidorenko SP, Chandran KA, Zhao Z, Shen SH, Fischer EH, Clark EA (February 1996). "CD22 associates with protein tyrosine phosphatase 1C, Syk, and phospholipase C-gamma(1) upon B cell activation". The Journal of Experimental Medicine. 183 (2): 547–60. doi:10.1084/jem.183.2.547. PMC 2192439. PMID 8627166.
- Adachi T, Wienands J, Wakabayashi C, Yakura H, Reth M, Tsubata T (July 2001). "SHP-1 requires inhibitory co-receptors to down-modulate B cell antigen receptor-mediated phosphorylation of cellular substrates". The Journal of Biological Chemistry. 276 (28): 26648–55. doi:10.1074/jbc.M100997200. PMID 11356834.
External links
- CD22+Antigen at the U.S. National Library of Medicine Medical Subject Headings (MeSH)
- Human CD22 genome location and CD22 gene details page in the UCSC Genome Browser.
- Overview of all the structural information available in the PDB for UniProt: P20273 (B-cell receptor CD22) at the PDBe-KB.