Nucleotide salvage
A salvage pathway is a pathway in which a biological product is produced from intermediates in the degradative pathway of its own or a similar substance. The term often refers to nucleotide salvage in particular, in which nucleotides (purine and pyrimidine) are synthesized from intermediates in their degradative pathway.
Nucleotide salvage pathways are used to recover bases and nucleosides that are formed during degradation of RNA and DNA. This is important in some organs because some tissues cannot undergo de novo synthesis. The salvaged products can then be converted back into nucleotides. Salvage pathways are targets for drug development, one family being called antifolates.[1]
A number of other biologically-important substances, like methionine and nicotinate, have their own salvage pathways to recycle parts of the molecule.
Substrates
The nucleotide salvage pathway requires distinct substrates:
Pyrimidines
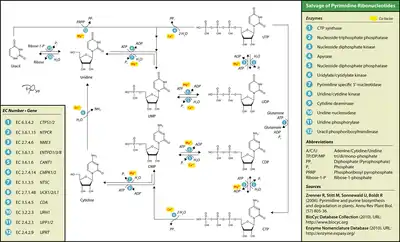
Uridine phosphorylase or pyrimidine-nucleoside phosphorylase substitutes the anomeric-carbon-bonded phosphate of ribose 1-phosphate for the free base uracil, forming the nucleoside uridine. Uridine kinase (aka uridine–cytidine kinase) can then phosphorylate the 5’-carbon of this nucleoside into uridine monophosphate (UMP). UMP/CMP kinase (EC 2.7.4.14) can phosphorylate UMP into uridine diphosphate, which nucleoside diphosphate kinase can phosphorylate into uridine triphosphate.
Thymidine phosphorylase or pyrimidine-nucleoside phosphorylase adds 2-deoxy-alpha-D-ribose 1-phosphate to thymine, with thymine bonding at the anomeric carbon of the deoxyribose, forming the deoxynucleoside thymidine. Thymidine kinase can then phosphorylate the 5’-carbon of this compound into thymidine monophosphate (TMP). Thymidylate kinase can phosphorylate TMP into thymidine diphosphate, which nucleoside diphosphate kinase can phosphorylate into thymidine triphosphate.
The nucleosides cytidine and deoxycytidine can be salvaged along the uracil pathway by cytidine deaminase, which converts them to uridine and deoxyuridine, respectively. Alternatively, uridine–cytidine kinase can phosphorylate them into cytidine monophosphate (CMP) or deoxycytidine monophosphate (dCMP). UMP/CMP kinase can phosphorylate (d)CMP into cytidine diphosphate or deoxycytidine diphosphate, which nucleoside diphosphate kinase can phosphorylate into cytidine triphosphate or deoxycytidine triphosphate.
Purines
Phosphoribosyltransferases add activated ribose-5-phosphate (Phosphoribosyl pyrophosphate, PRPP) to bases, creating nucleoside monophosphates. There are two types of phosphoribosyltransferases: adenine phosphoribosyltransferase (APRT) and hypoxanthine-guanine phosphoribosyltransferase (HGPRT). HGPRT is an important enzyme in Purine pathway metabolism and[2] its deficiency is implicated in Lesch–Nyhan syndrome.
| Nucleobase | Enzyme | Nucleotide |
| hypoxanthine | hypoxanthine/guanine phosphoribosyl transferase (HGPRT) | IMP |
| guanine | hypoxanthine/guanine phosphoribosyl transferase (HGPRT) | GMP |
| adenine | adenine phosphoribosyltransferase (APRT) | AMP |
Folate biosynthesis
Tetrahydrofolic acid and its derivatives are produced by salvage pathways from GTP.[1]
Other salvage pathways
L-methionine salvage is the pathway that regenerates methionine from its downstream products. A version of the pathway uses methylthioadenosine (MTA), forming the so-called MTA cycle with its synthesizing reaction. This sulphur-recycling action is found in humans, and seems to be universal among aerobic life.[3][4]
Nicotinate salvage is the process of regenerating nicotinamide adenine dinucleotide from nicotinic acid. This pathway is important for controlling the level of oxidative stress in cells. The human gene NAPRT encodes the main enzyme in the pathway.[5] Cancer cells, which have increased NAD requirements, tend to upregulate the pathway.[6]
Salvage pathways also exist for ceramide, cobalamin, cell wall components, and tetrahydrobiopterin in various organisms.
References
- Kompis IM, Islam K, Then RL (February 2005). "DNA and RNA synthesis: antifolates". Chemical Reviews. 105 (2): 593–620. doi:10.1021/cr0301144. PMID 15700958.
- Ansari MY, Equbal A, Dikhit MR, Mansuri R, Rana S, Ali V, Sahoo GC, Das P (February 2016). "Establishment of correlation between in-silico and in-vitro test analysis against Leishmania HGPRT to inhibitors". International Journal of Biological Macromolecules. 83: 78–96. doi:10.1016/j.ijbiomac.2015.11.051. PMID 26616453.
- Albers, E (December 2009). "Metabolic characteristics and importance of the universal methionine salvage pathway recycling methionine from 5'-methylthioadenosine". IUBMB Life. 61 (12): 1132–42. doi:10.1002/iub.278. PMID 19946895.
- Sekowska, A; Ashida, H; Danchin, A (January 2019). "Revisiting the methionine salvage pathway and its paralogues". Microbial Biotechnology. 12 (1): 77–97. doi:10.1111/1751-7915.13324. PMC 6302742. PMID 30306718.
- Hara, N; Yamada, K; Shibata, T; Osago, H; Hashimoto, T; Tsuchiya, M (24 August 2007). "Elevation of cellular NAD levels by nicotinic acid and involvement of nicotinic acid phosphoribosyltransferase in human cells". The Journal of Biological Chemistry. 282 (34): 24574–82. doi:10.1074/jbc.M610357200. PMID 17604275.
- Duarte-Pereira, S; Pereira-Castro, I; Silva, SS; Correia, MG; Neto, C; da Costa, LT; Amorim, A; Silva, RM (9 December 2015). "Extensive regulation of nicotinate phosphoribosyltransferase (NAPRT) expression in human tissues and tumors". Oncotarget. 7 (2): 1973–1983. doi:10.18632/ONCOTARGET.6538. ISSN 1949-2553. PMC 4811510. PMID 26675378. Wikidata Q36739012.