Parareptilia
Parareptilia ("near-reptiles") is a subclass or clade of basal sauropsids, typically considered the sister taxon to Eureptilia (the group that likely contains all living reptiles and birds). Parareptiles first arose near the end of the Carboniferous period and achieved their highest diversity during the Permian period. Several ecological innovations were first accomplished by parareptiles among reptiles. These include the first reptiles to return to marine ecosystems (mesosaurs), the first bipedal reptiles (bolosaurids such as Eudibamus), the first reptiles with advanced hearing systems (nycteroleterids and others), and the first large herbivorous reptiles (the pareiasaurs). The only parareptiles to survive into the Triassic period were the procolophonoids, a group of small generalists, omnivores, and herbivores. The largest family of procolophonoids, the procolophonids, rediversified in the Triassic, but subsequently declined and became extinct by the end of the period.[2][3]
| Parareptiles | |
|---|---|
 | |
| A collage of five parareptile fossils. Clockwise from top, Mesosaurus tenuidens (a mesosaur), Delorhynchus cifellii (a probable acleistorhinid), Scutosaurus karpinskii (a pareiasaur), Nyctiphruretus acudens (a nyctiphruretid), Hypsognathus fenneri (a procolophonid) | |
| Scientific classification | |
| Domain: | Eukaryota |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Reptilia |
| Clade: | †Parareptilia Olson, 1947 |
| Orders | |
Compared to most eureptiles, parareptiles retained fairly "primitive" characteristics such as robust, low-slung bodies and large supratemporal bones at the back of the skull. While all but the earliest eureptiles were diapsids, with two openings at the back of the skull, parareptiles were generally more conservative in the extent of temporal fenestration. In its modern usage, Parareptilia was first utilized as a cladistically correct alternative to Anapsida, a term which historically referred to reptiles with solid skulls lacking holes behind the eyes.[4] Nevertheless, not all parareptiles have 'anapsid' skulls, and some do have large holes in the back of the skull. They also had several unique adaptations, such as a large pit on the maxilla, a broad prefrontal-palatine contact, and the absence of a supraglenoid foramen of the scapula.[4][5]
Like many other so-called 'anapsids', parareptiles were historically understudied. Interest in their relationships were reinvigorated in the 1990s, when several studies argued that Testudines (turtles and their kin) were members of Parareptilia.[4] Although this would suggest that Parareptilia was not extinct after all, the origin of turtles is still heavily debated. Many other morphological or genetic analyses find more support for turtles among diapsid eureptiles such as sauropterygians or archosauromorphs, rather than parareptiles.[6][7][8][3]
Description
Skull
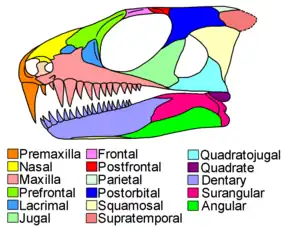
Parareptilian skulls were diverse, from mesosaurs with elongated snouts filled with hundreds of thin teeth, to the snub-nosed, knob-encrusted skulls of pareiasaurs. Parareptile teeth were quite variable in shape and function between different species. However, they were relatively homogenous on the same skull. While most synapsids and many early eureptiles had a caniform region of enlarged fang-like teeth in the front half of the skull, very few parareptiles possessed caniform teeth.[5]
Many amniotes have a row of small pits running along bones at the edge of the mouth, but parareptiles have only a few pits, with one especially large pit near the front of the maxilla.[4][9][7] The rest of the skull was often strongly-textured by pits, ridges, and rugosities in most parareptile groups, occasionally culminating in complex bosses or spines. The maxilla is usually low, while the prefrontal and lacrimal bones in front of the eye are both fairly large. In all parareptiles except mesosaurs, the prefrontal has a plate-like inner branch which forms a broad contact with the palatine bone of the palate.[4][7][5] A prominent hole, the foramen orbitonasale, is present at the intersection of the prefrontal, palatine, and lacrimal. Parareptilian palates also have toothless and reduced ectopterygoid bones, a condition taken to extremes in mesosaurs, which have lost the ectopterygoid entirely.[4][5]
Most parareptiles had large orbits (eye sockets), significantly longer (from front-to-back) than the region of the skull behind the eyes.[5] The jugal bone, which forms the lower and rear edge of the orbit, has a very thin suborbital process (front branch), usually no subtemporal process (lower rear branch), and a thick dorsal process (upper rear branch). The squamosal and quadratojugal bones, which lie behind the jugal, are quite large and are embayed from behind to accommodate the internal ears.[4][5] Parareptiles were traditionally considered to have an ‘anapsid’-type skull, with the jugal, squamosal, and quadratojugal firmly sutured together without any gaps or slits between them. This principle still holds true for some subgroups, such as pareiasaurs. However, a growing number of parareptile taxa are now known to have had an infratemporal fenestra, a large hole or emargination lying among the bones behind the eye. In some taxa, the margins of such openings may include additional bones such as the maxilla or postorbital.[10][11] When seen from above, the rear edge of the skull is straight or has a broad median embayment.[5] From inside to outside, the rear edge of the skull is formed by three pairs of bones: the postparietals, tabulars, and supratemporals. Parareptiles have particularly large supratemporals, which often extend further backwards than the tabulars.[12]
Apart from the long, slender jaws of mesosaurs, most parareptile jaws were short and thick. The jaw joint is formed by the articular (in the lower jaw) and the quadrate (in the upper jaw). In many parareptiles, the jaw joint is shifted forwards on the skull past the rear part of the braincase.[4][7] Jaw muscles attach to the coronoid process, a triangular spur in the rear half of the jaw. Both the tooth-bearing dentary bone and the posterior foramen intermandibularis (a hole on the inner surface of the jaw) reach as far back as the coronoid process.[4][5] The surangular bone, which forms the upper rear part of the jaw, is narrow and plate-like.[13]
Postcranial skeleton
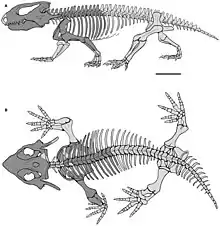
There was some variation in the body shape of parareptiles, with early members of the group having an overall lizard-like appearance, with thin limbs and long tails. The most successful and diverse groups of parareptiles, the pareiasaurs and procolophonids, had massively-built bodies with reduced tails and stout limbs with short digits. This general body shape is shared with other ‘cotylosaurs’ such as captorhinids, diadectomorphs, and seymouriamorphs.[3] Another general ‘cotylosaurian’ feature in parareptiles is the ‘swollen’ appearance of their vertebrae, which have wide and convex upper surfaces.[12]
Parareptiles lacked a supraglenoid foramen on the scapula, a hole which is also absent in varanopids and neodiapsids.[5][11] Most had a fairly short and thick humerus which was expanded near the elbow. Unlike early eureptiles, the outer part of the lower humerus possessed both a small supinator process and an ectepicondylar foramen and groove.[4] The ulna generally has a poorly developed olecranon process, another trait in contrast with the earliest eureptiles.[4][5]
Most parareptiles had an ilium which was fan-shaped and vertically (rather than horizontally) oriented, an unusual trait among early amniotes.[4][9][7] The sacral ribs, which connect the spine to the ilium, were usually slender or fan-shaped, with large gaps between them.[4] The hindlimbs were typically not much longer than the forelimbs, and had thick reptilian ankle bones and short toes. There are some exceptions, such as Eudibamus, an early Permian bolosaurid with very elongated hindlimbs.[14]
History of classification
The name Parareptilia was coined by Olson in 1947 to refer to an extinct group of Paleozoic reptiles, as opposed to the rest of the reptiles or Eureptilia ("true reptiles").[15] Olsen's term was generally ignored, and various taxa now known as parareptiles were generally not placed into exclusive groups with each other. Many were classified as 'cotylosaurs' (a wastebasket taxon of stout-bodied 'primitive' reptiles or reptile-like tetrapods) or 'anapsids' (reptiles without temporal fenestrae, such as modern turtles).
Parareptilia's usage was revived by cladistic studies, to refer to those traditional 'anapsids' that were thought to be unrelated to turtles. Gauthier et al. (1988) provided the first phylogenetic definitions for the names of many amniote taxa and argued that captorhinids and turtles were sister groups, constituting the clade Anapsida (in a much more limited context than typically applied). A name had to be found for a clade of various early-diversing Permian and Triassic reptiles no longer included in the anapsids. Olsen's term "parareptiles" was chosen to refer to this clade, although its instability within their analysis meant that Gauthier et al. (1988) were not confident enough to erect Parareptilia as a formal taxon. Their cladogram is as follows:[16]
| Amniota |
| |||||||||||||||||||||||||||||||||||||||||||||||||||



Laurin & Reisz (1995) found a slightly different topology, in which Reptilia is divided into Parareptilia and Eureptilia. They argued that Testudines (turtles) were members of Parareptilia; in fact, they explicitly defined Parareptilia as "Testudines and all amniotes more closely related to them than to diapsids". Captorhinidae was transferred to Eureptilia, while Parareptilia included turtles alongside many of the taxa named as such by Gauthier et al. (1988). There was one major exception: mesosaurs were placed outside both groups, as the sister taxon to the crown group Reptilia. Mesosaurs were still considered sauropsids, as they were closer to reptiles than to synapsids. The traditional group 'Anapsida' was rejected as a paraphyletic assemblage. The cladogram of Laurin & Reisz (1995) is provided below:[4]
| Amniota |
| |||||||||||||||||||||||||||||||||||||||||||||||||||
_(Varanus_griseus).png.webp)
In contrast, several studies in the mid-to-late 1990s by Olivier Rieppel and Michael deBraga argued that turtles were actually lepidosauromorph diapsids related to the sauropterygians.[17][18][9][6][7] The diapsid affinities of turtles have been supported by molecular phylogenies.[19][20][21][22] The first genome-wide phylogenetic analysis was completed by Wang et al. (2013). Using the draft genomes of Chelonia mydas and Pelodiscus sinensis, the team used the largest turtle data set to date in their analysis and concluded that turtles are likely a sister group of crocodilians and birds (Archosauria).[8] This placement within the diapsids suggests that the turtle lineage lost diapsid skull characteristics, since turtles possess an anapsid skull. This would make Parareptilia a totally extinct group with skull features that resemble those of turtles through convergent evolution. With turtles positioned outside of parareptiles, Tsuji and Müller (2009) redefined Parareptilia as "the most inclusive clade containing Milleretta rubidgei and Procolophon trigoniceps, but not Captorhinus aguti."[3]
The cladogram below follows an analysis by M.S. Lee, in 2013.[23]
| Amniota |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||



The cladogram below follows the analysis of Li et al. (2018).[24]
|
†Parareptilia | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Eureptilia |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

A 2020 study by David P. Ford and Roger B. J. Benson found that Parareptilia was nested within Diapsida as the sister group to Neodiapsida, with the clade containing Neodiapsida and Parareptilia dubbed Neoreptilia, which suggests that parareptiles were ancestrally diapsid. This excluded mesosaurs, which were again found to be basal among the sauropsids.[11] Some studes have found Parareptilia to be paraphyletic, with some parareptiles more closely related to diapsids that to other parareptiles, with Simões et al. (2022) using Neoreptilia for the clade containing Procolophonomorpha+Neodiapsida.[25]
Evolutionary history
The oldest known parareptiles are the bolosaur Erpetonyx and the acleistorhinid Carbonodraco from the Late Carboniferous (Moscovian-Gzhelian) of North America, which represents the only currently known Carboniferous parareptiles, indicating that the initial diversification of the group took place in the Late Carboniferous.[1] Numerous parareptile lineages appeared during the early Permian and the group reached a cosmopolitan distribution. Parareptile diversity declined towards the end of the Permian and procolophonoids, which first appeared during the Late Permian, were the only group of parareptiles to survive the Permian–Triassic extinction event. Procolophonid diversity sharply declined beginning in the Middle Triassic, with the group becoming extinct by the end of the Triassic.[26]
References
- Mann A, McDaniel EJ, McColville ER, Maddin HC (November 2019). "Carbonodraco lundi gen et sp. nov., the oldest parareptile, from Linton, Ohio, and new insights into the early radiation of reptiles". Royal Society Open Science. 6 (11): 191191. Bibcode:2019RSOS....691191M. doi:10.1098/rsos.191191. PMC 6894558. PMID 31827854.
- Botha-Brink J, Smith RM (2012-09-01). "Palaeobiology of Triassic procolophonids, inferred from bone microstructure". Comptes Rendus Palevol. 11 (6): 419–433. doi:10.1016/j.crpv.2012.03.002. ISSN 1631-0683.
- Tsuji LA, Müller J (2009). "Assembling the history of the Parareptilia: phylogeny, diversification, and a new definition of the clade". Fossil Record. 12 (1): 71–81. doi:10.1002/mmng.200800011. ISSN 1860-1014.
- Laurin M, Reisz RR (1995-02-01). "A reevaluation of early amniote phylogeny" (PDF). Zoological Journal of the Linnean Society. 113 (2): 165–223. doi:10.1111/j.1096-3642.1995.tb00932.x. ISSN 0024-4082.
- Müller J, Tsuji LA (September 2007). "Impedance-matching hearing in Paleozoic reptiles: evidence of advanced sensory perception at an early stage of amniote evolution". PLOS ONE. 2 (9): e889. Bibcode:2007PLoSO...2..889M. doi:10.1371/journal.pone.0000889. PMC 1964539. PMID 17849018.
- Rieppel O, deBraga M (December 1996). "Turtles as diapsid reptiles". Nature. 384 (6608): 453–455. Bibcode:1996Natur.384..453R. doi:10.1038/384453a0. ISSN 1476-4687. S2CID 4264378.
- deBraga M, Rieppel O (1997-07-01). "Reptile phylogeny and the interrelationships of turtles". Zoological Journal of the Linnean Society. 120 (3): 281–354. doi:10.1111/j.1096-3642.1997.tb01280.x. ISSN 0024-4082.
- Wang Z, Pascual-Anaya J, Zadissa A, Li W, Niimura Y, Huang Z, et al. (June 2013). "The draft genomes of soft-shell turtle and green sea turtle yield insights into the development and evolution of the turtle-specific body plan". Nature Genetics. 45 (6): 701–706. doi:10.1038/ng.2615. PMC 4000948. PMID 23624526.
- Debraga M, Reisz RR (1996-09-19). "The Early Permian reptile Acleistorhinus pteroticus and its phylogenetic position" (PDF). Journal of Vertebrate Paleontology. 16 (3): 384–395. doi:10.1080/02724634.1996.10011328. ISSN 0272-4634.
- MacDougall MJ, Reisz RR (2014). "The first record of a nyctiphruretid parareptile from the Early Permian of North America, with a discussion of parareptilian temporal fenestration". Zoological Journal of the Linnean Society. 172 (3): 616–630. doi:10.1111/zoj.12180.
- Ford DP, Benson RB (January 2020). "The phylogeny of early amniotes and the affinities of Parareptilia and Varanopidae". Nature Ecology & Evolution. 4 (1): 57–65. doi:10.1038/s41559-019-1047-3. PMID 31900445. S2CID 209673326.
- Tsuji LA, Müller J, Reisz RR (2012-01-01). "Anatomy of Emeroleter levis and the phylogeny of the nycteroleter parareptiles" (PDF). Journal of Vertebrate Paleontology. 32 (1): 45–67. doi:10.1080/02724634.2012.626004. ISSN 0272-4634. S2CID 55268829.
- Haridy Y, Macdougall MJ, Reisz RR (13 December 2017). "The lower jaw of the Early Permian parareptile Delorhynchus, first evidence of multiple denticulate coronoids in a reptile". Zoological Journal of the Linnean Society. 184 (3): 791–803. doi:10.1093/zoolinnean/zlx085. ISSN 0024-4082.
- Berman DS, Reisz RR, Scott D, Henrici AC, Sumida SS, Martens T (November 2000). "Early Permian bipedal reptile". Science. 290 (5493): 969–972. Bibcode:2000Sci...290..969B. doi:10.1126/science.290.5493.969. PMID 11062126.
- Olson EC (1947). Roy SK (ed.). "The family Diadectidae and its bearing on the classification of reptiles". Geology. Fieldiana. Chicago: Chicago Natural History Museum. 11 (1): 1–53. doi:10.5962/bhl.title.3579. Retrieved 15 January 2015.
- Gauthier JA, Kluge AG, Rowe T (1988). "The early evolution of the Amniota". In Benton MJ (ed.). The Phylogeny and Classification of the Tetrapods, Volume 1: Amphibians, Reptiles, Birds. Systematics Association Special Volume. Vol. 35A. Clarendon Press. pp. 103–155.
- Rieppel O (1994). "Osteology of Simosaurus gaillardoti and the relationships of stem-group sauropterygia". Fieldiana Geology. 28 (1462): i-85.
- Rieppel O (1995). "Studies on skeleton formation in reptiles: implications for turtle relationships". Zoology. 98: 298–308.
- Zardoya R, Meyer A (November 1998). "Complete mitochondrial genome suggests diapsid affinities of turtles". Proceedings of the National Academy of Sciences of the United States of America. 95 (24): 14226–14231. Bibcode:1998PNAS...9514226Z. doi:10.1073/pnas.95.24.14226. PMC 24355. PMID 9826682.
- Iwabe N, Hara Y, Kumazawa Y, Shibamoto K, Saito Y, Miyata T, Katoh K (April 2005). "Sister group relationship of turtles to the bird-crocodilian clade revealed by nuclear DNA-coded proteins". Molecular Biology and Evolution. 22 (4): 810–813. doi:10.1093/molbev/msi075. PMID 15625185.
- Roos J, Aggarwal RK, Janke A (November 2007). "Extended mitogenomic phylogenetic analyses yield new insight into crocodylian evolution and their survival of the Cretaceous-Tertiary boundary". Molecular Phylogenetics and Evolution. 45 (2): 663–673. doi:10.1016/j.ympev.2007.06.018. PMID 17719245.
- Katsu Y, Braun EL, Guillette LJ, Iguchi T (2009). "From reptilian phylogenomics to reptilian genomes: analyses of c-Jun and DJ-1 proto-oncogenes". Cytogenetic and Genome Research. 127 (2–4): 79–93. doi:10.1159/000297715. PMID 20234127. S2CID 12116018.
- Lee MS (December 2013). "Turtle origins: insights from phylogenetic retrofitting and molecular scaffolds". Journal of Evolutionary Biology. 26 (12): 2729–2738. doi:10.1111/jeb.12268. PMID 24256520. S2CID 2106400.
- Li, Chun; Fraser, Nicholas C.; Rieppel, Olivier; Wu, Xiao-Chun (August 2018). "A Triassic stem turtle with an edentulous beak". Nature. 560 (7719): 476–479. doi:10.1038/s41586-018-0419-1. ISSN 0028-0836. PMID 30135526. S2CID 52067286.
- Simões, T. R.; Kammerer, C. F.; Caldwell, M. W.; Pierce, S. E. (2022). "Successive climate crises in the deep past drove the early evolution and radiation of reptiles". Science Advances. 8 (33): eabq1898. doi:10.1126/sciadv.abq1898. PMC 9390993. PMID 35984885.
- MacDougall MJ, Brocklehurst N, Fröbisch J (March 2019). "Species richness and disparity of parareptiles across the end-Permian mass extinction". Proceedings. Biological Sciences. 286 (1899): 20182572. doi:10.1098/rspb.2018.2572. PMC 6452079. PMID 30890099.