Sigma factor
A sigma factor (σ factor or specificity factor) is a protein needed for initiation of transcription in bacteria.[1][2] It is a bacterial transcription initiation factor that enables specific binding of RNA polymerase (RNAP) to gene promoters. It is homologous to archaeal transcription factor B and to eukaryotic factor TFIIB.[3] The specific sigma factor used to initiate transcription of a given gene will vary, depending on the gene and on the environmental signals needed to initiate transcription of that gene. Selection of promoters by RNA polymerase is dependent on the sigma factor that associates with it.[4] They are also found in plant chloroplasts as a part of the bacteria-like plastid-encoded polymerase (PEP).[5]
The sigma factor, together with RNA polymerase, is known as the RNA polymerase holoenzyme. Every molecule of RNA polymerase holoenzyme contains exactly one sigma factor subunit, which in the model bacterium Escherichia coli is one of those listed below. The number of sigma factors varies between bacterial species.[1][6] E. coli has seven sigma factors. Sigma factors are distinguished by their characteristic molecular weights. For example, σ70 is the sigma factor with a molecular weight of 70 kDa.
The sigma factor in the RNA polymerase holoenzyme complex is required for the initiation of transcription, although once that stage is finished, it is dissociated from the complex and the RNAP continues elongation on its own.
Specialized sigma factors
Different sigma factors are utilized under different environmental conditions. These specialized sigma factors bind the promoters of genes appropriate to the environmental conditions, increasing the transcription of those genes.
Sigma factors in E. coli:
- σ70(RpoD) – σA – the "housekeeping" sigma factor or also called as primary sigma factor (Group 1), transcribes most genes in growing cells. Every cell has a "housekeeping" sigma factor that keeps essential genes and pathways operating.[1] In the case of E. coli and other gram-negative rod-shaped bacteria, the "housekeeping" sigma factor is σ70.[1] Genes recognized by σ70 all contain similar promoter consensus sequences consisting of two parts.[1] Relative to the DNA base corresponding to the start of the RNA transcript, the consensus promoter sequences are characteristically centered at 10 and 35 nucleotides before the start of transcription (−10 and −35).
- σ19 (FecI) – the ferric citrate sigma factor, regulates the fec gene for iron transport and metabolism
- σ24 (RpoE) – extreme heat stress response and the extracellular proteins sigma factor
- σ28 (RpoF/FliA) – the flagellar synthesis and chemotaxis sigma factor
- σ32 (RpoH) – the heat shock sigma factor, it is turned on when the bacteria are exposed to heat. Due to the higher expression, the factor will bind with a high probability to the polymerase-core-enzyme. Doing so, other heatshock proteins are expressed, which enable the cell to survive higher temperatures. Some of the enzymes that are expressed upon activation of σ32 are chaperones, proteases and DNA-repair enzymes.
- σ38 (RpoS) – the starvation/stationary phase sigma factor
- σ54 (RpoN) – the nitrogen-limitation sigma factor
There are also anti-sigma factors that inhibit the function of sigma factors and anti-anti-sigma factors that restore sigma factor function.
Structure
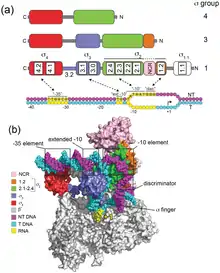
By sequence similarity, most sigma factors are σ70-like (InterPro: IPR000943). They have four main regions (domains) that are generally conserved:
N-terminus --------------------- C-terminus
1.1 2 3 4
The regions are further subdivided. For example, region 2 includes 1.2 and 2.1 through 2.4.
Domain 1.1 is found only in "primary sigma factors" (RpoD, RpoS in E.coli; "Group 1"). It is involved in ensuring the sigma factor will only bind the promoter when it is complexed with the RNA polymerase.[7] Domains 2-4 each interact with specific promoter elements and with RNAP. Region 2.4 recognizes and binds to the promoter −10 element (called the "Pribnow box"). Region 4.2 recognizes and binds to the promoter −35 element.[7]
Not every sigma factor of the σ70 family contains all the domains. Group 2, which includes RpoS, is very similar to Group 1 but lacks domain 1. Group 3 also lacks domain 1, and includes σ28. Group 4, also known as the Extracytoplasmic Function (ECF) group, lack both σ1.1 and σ3. RpoE is a member.[7]
Other known sigma factors are of the σ54/RpoN (InterPro: IPR000394) type. They are functional sigma factors, but they have significantly different primary amino acid sequences.[8]
|
|
|
|
|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
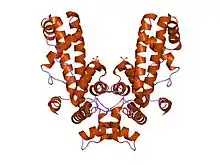


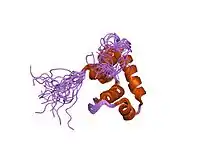
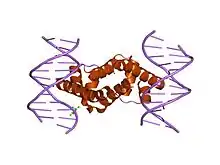
Retention during transcription elongation
The core RNA polymerase (consisting of 2 alpha (α), 1 beta (β), 1 beta-prime (β'), and 1 omega (ω) subunits) binds a sigma factor to form a complex called the RNA polymerase holoenzyme. It was previously believed that the RNA polymerase holoenzyme initiates transcription, while the core RNA polymerase alone synthesizes RNA. Thus, the accepted view was that sigma factor must dissociate upon transition from transcription initiation to transcription elongation (this transition is called "promoter escape"). This view was based on analysis of purified complexes of RNA polymerase stalled at initiation and at elongation. Finally, structural models of RNA polymerase complexes predicted that, as the growing RNA product becomes longer than ~15 nucleotides, sigma must be "pushed out" of the holoenzyme, since there is a steric clash between RNA and a sigma domain. However, σ70 can remain attached in complex with the core RNA polymerase in early elongation[9] and sometimes throughout elongation.[10] Indeed, the phenomenon of promoter-proximal pausing indicates that sigma plays roles during early elongation. All studies are consistent with the assumption that promoter escape reduces the lifetime of the sigma-core interaction from very long at initiation (too long to be measured in a typical biochemical experiment) to a shorter, measurable lifetime upon transition to elongation.
Sigma cycle
It had long been thought that the sigma factor obligatorily leaves the core enzyme once it has initiated transcription, allowing it to link to another core enzyme and initiate transcription at another site. Thus, the sigma factor would cycle from one core to another. However, fluorescence resonance energy transfer was used to show that the sigma factor does not obligatorily leave the core.[9] Instead, it changes its binding with the core during initiation and elongation. Therefore, the sigma factor cycles between a strongly bound state during initiation and a weakly bound state during elongation.
Sigma factor competition
The number of RNAPs in bacterial cells (e.g., E. coli) have been shown to be smaller than the number of sigma factors. Consequently, if a certain sigma factor is overexpressed, not only will increase the expression levels of genes whose promoters have preference for that sigma factor, but it will also reduce the probability that genes with promoters with preference for other sigma factors.[11][12][13][14]
Meanwhile, transcription initiation has two major rate limiting steps: the closed and the open complex formation. However, only the dynamics of the first step depends on the concentration of sigma factors. Interestingly, the fastest is the closed complex formation relative to the open complex formation, the less responsive is a promoter to changes in sigma factors’ concentration (see [14] for a model and empirical data of this phenomenon).
Genes with dual sigma factor preference
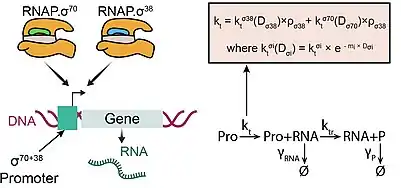
While most genes of E. coli can be recognized by an RNAP with one and only one type of sigma factor (e.g. sigma 70), a few genes (~ 5%) have what is called a “dual sigma factor preference”,[15] that is, they can respond to two different sigma factors, as reported in RegulonDB.[16] The most common ones are those promoters that can respond to both sigma 70 and to sigma 38 (iIlustrated in the figure) . Studies of the dynamics of these genes showed that when the cells enter stationary growth they are almost as induced as those genes that have preference for σ38 alone. This induction level was shown to be predictable from their promoter sequence.[15] A model of their dynamics is shown in the figure. In the future, these promoters may become useful tools in synthetic genetic constructs in E. coli.
See also
- Ekkehard Bautz
References
- Gruber TM, Gross CA (2003). "Multiple sigma subunits and the partitioning of bacterial transcription space". Annual Review of Microbiology. 57: 441–66. doi:10.1146/annurev.micro.57.030502.090913. PMID 14527287.
- Kang JG, Hahn MY, Ishihama A, Roe JH (July 1997). "Identification of sigma factors for growth phase-related promoter selectivity of RNA polymerases from Streptomyces coelicolor A3(2)". Nucleic Acids Research. 25 (13): 2566–73. doi:10.1093/nar/25.13.2566. PMC 146787. PMID 9185565.
- Burton SP, Burton ZF (6 November 2014). "The σ enigma: bacterial σ factors, archaeal TFB and eukaryotic TFIIB are homologs". Transcription. 5 (4): e967599. doi:10.4161/21541264.2014.967599. PMC 4581349. PMID 25483602.
- Ho TD, Ellermeier CD (April 2012). "Extra cytoplasmic function σ factor activation". Current Opinion in Microbiology. 15 (2): 182–8. doi:10.1016/j.mib.2012.01.001. PMC 3320685. PMID 22381678.
- Schweer J, Türkeri H, Kolpack A, Link G (December 2010). "Role and regulation of plastid sigma factors and their functional interactors during chloroplast transcription - recent lessons from Arabidopsis thaliana". European Journal of Cell Biology. 89 (12): 940–6. doi:10.1016/j.ejcb.2010.06.016. PMID 20701995.
- Sharma UK, Chatterji D (September 2010). "Transcriptional switching in Escherichia coli during stress and starvation by modulation of sigma activity". FEMS Microbiology Reviews. 34 (5): 646–57. doi:10.1111/j.1574-6976.2010.00223.x. PMID 20491934.
- Paget MS (June 2015). "Bacterial Sigma Factors and Anti-Sigma Factors: Structure, Function and Distribution". Biomolecules. 5 (3): 1245–65. doi:10.3390/biom5031245. PMC 4598750. PMID 26131973.
- Merrick MJ (December 1993). "In a class of its own--the RNA polymerase sigma factor sigma 54 (sigma N)". Molecular Microbiology. 10 (5): 903–9. doi:10.1111/j.1365-2958.1993.tb00961.x. PMID 7934866. S2CID 84789281.
- Kapanidis AN, Margeat E, Laurence TA, Doose S, Ho SO, Mukhopadhyay J, Kortkhonjia E, Mekler V, Ebright RH, Weiss S (November 2005). "Retention of transcription initiation factor sigma70 in transcription elongation: single-molecule analysis". Molecular Cell. 20 (3): 347–56. doi:10.1016/j.molcel.2005.10.012. PMID 16285917.
- Harden TT, Wells CD, Friedman LJ, Landick R, Hochschild A, Kondev J, Gelles J (January 2016). "Bacterial RNA polymerase can retain σ70 throughout transcription". Proc Natl Acad Sci U S A. 113 (3): 602–7. Bibcode:2016PNAS..113..602H. doi:10.1073/pnas.1513899113. PMC 4725480. PMID 26733675.
- Jishage, M; Ishihama, A (December 1995). "Regulation of RNA polymerase sigma subunit synthesis in Escherichia coli: intracellular levels of sigma 70 and sigma 38". Journal of Bacteriology. 177 (23): 6832–6835. doi:10.1128/jb.177.23.6832-6835.1995. ISSN 0021-9193. PMC 177550. PMID 7592475.
- Jishage, M; Iwata, A; Ueda, S; Ishihama, A (September 1996). "Regulation of RNA polymerase sigma subunit synthesis in Escherichia coli: intracellular levels of four species of sigma subunit under various growth conditions". Journal of Bacteriology. 178 (18): 5447–5451. doi:10.1128/jb.178.18.5447-5451.1996. ISSN 0021-9193. PMC 178365. PMID 8808934.
- Grigorova, Irina L.; Phleger, Naum J.; Mutalik, Vivek K.; Gross, Carol A. (2006-04-04). "Insights into transcriptional regulation and σ competition from an equilibrium model of RNA polymerase binding to DNA". Proceedings of the National Academy of Sciences. 103 (14): 5332–5337. Bibcode:2006PNAS..103.5332G. doi:10.1073/pnas.0600828103. ISSN 0027-8424. PMC 1459355. PMID 16567622.
- Kandavalli, Vinodh K.; Tran, Huy; Ribeiro, Andre S. (October 2016). "Effects of σ factor competition are promoter initiation kinetics dependent". Biochimica et Biophysica Acta (BBA) - Gene Regulatory Mechanisms. 1859 (10): 1281–1288. doi:10.1016/j.bbagrm.2016.07.011. PMID 27452766.
- Baptista, Ines S.C.; Kandavalli, Vinodh; Chauhan, Vatsala; Bahrudeen, Mohamed N.M.; Almeida, Bilena L.B.; Palma, Cristina S.D.; Dash, Suchintak; Ribeiro, Andre S. (April 2022). "Sequence-dependent model of genes with dual σ factor preference". Biochimica et Biophysica Acta (BBA) - Gene Regulatory Mechanisms. 1865 (3): 194812. doi:10.1016/j.bbagrm.2022.194812. PMID 35338024. S2CID 247636833.
- Santos-Zavaleta, Alberto; Salgado, Heladia; Gama-Castro, Socorro; Sánchez-Pérez, Mishael; Gómez-Romero, Laura; Ledezma-Tejeida, Daniela; García-Sotelo, Jair Santiago; Alquicira-Hernández, Kevin; Muñiz-Rascado, Luis José; Peña-Loredo, Pablo; Ishida-Gutiérrez, Cecilia (2019-01-08). "RegulonDB v 10.5: tackling challenges to unify classic and high throughput knowledge of gene regulation in E. coli K-12". Nucleic Acids Research. 47 (D1): D212–D220. doi:10.1093/nar/gky1077. ISSN 0305-1048. PMC 6324031. PMID 30395280.
External links
- Sigma+Factor at the US National Library of Medicine Medical Subject Headings (MeSH)