Perivascular space
A perivascular space, also known as a Virchow–Robin space, is a fluid-filled space surrounding certain blood vessels in several organs, including the brain,[1] potentially having an immunological function, but more broadly a dispersive role for neural and blood-derived messengers.[2] The brain pia mater is reflected from the surface of the brain onto the surface of blood vessels in the subarachnoid space. In the brain, perivascular cuffs are regions of leukocyte aggregation in the perivascular spaces, usually found in patients with viral encephalitis.
| Perivascular space | |
|---|---|
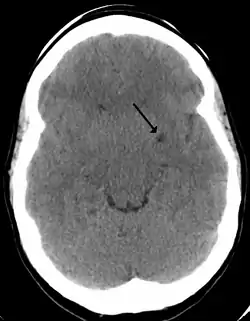 A perivascular space as seen on CT | |
| Anatomical terms of neuroanatomy |
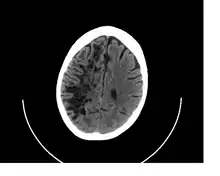
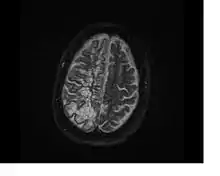
Perivascular spaces vary in dimension according to the type of blood vessel. In the brain where most capillaries have an imperceptible perivascular space, select structures of the brain, such as the circumventricular organs, are notable for having large perivascular spaces surrounding highly permeable capillaries, as observed by microscopy. The median eminence, a brain structure at the base of the hypothalamus, contains capillaries with wide perivascular spaces.[3]
In humans, perivascular spaces surround arteries and veins can usually be seen as areas of dilatation on MRI images. While many normal brains will show a few dilated spaces, an increase in these spaces may correlate with the incidence of several neurodegenerative diseases, making the spaces a topic of research.[4]
Structure
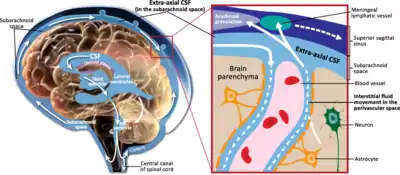
Perivascular spaces are gaps containing interstitial fluid that span between blood vessels and their host organ, such as the brain, which they penetrate and serve as extravascular channels through which solutes can pass.[2][5] Like the blood vessels around which they form, perivascular spaces are found in both the brain subarachnoid space and the subpial space.[6]
Perivascular spaces surrounding arteries in the cerebral cortex and the basal ganglia are separated from the subpial space by one or two layers of leptomeninges, respectively, as well as the pia mater.[7] By virtue of the leptomeningeal cell layer, the perivascular spaces belonging to the subarachnoid space are continuous with those of the subpial space. The direct communication between the perivascular spaces of the subarachnoid space and the subpial space is unique to the brain’s arteries, as no leptomeningeal layers surround the brain’s veins.[7][6] Use of the scanning electron microscope has determined that the spaces surrounding blood vessels in the subarachnoid space are not continuous with the subarachnoid space because of the presence of pia mater cells joined by desmosomes.[8]
Perivascular spaces, especially around fenestrated capillaries, are found in many organs, such as the thymus, liver, kidneys, spleen, bones, and pineal gland.[9][10][11][12] Particularly within the brain circumventricular organs – subfornical organ, area postrema, and median eminence – large perivascular spaces are present around fenestrated capillaries, indicating that the spaces serve a dispersive role for brain- or bloodborne messengers.[2]
Perivascular spaces may be enlarged to a diameter of five millimeters in healthy humans and do not imply disease. When enlarged, they can disrupt the function of the brain regions into which they project.[5] Dilation can occur on one or both sides of the brain.[7]
Dilated perivascular spaces are categorized into three types:[7]
- Type 1 are located on the lenticulostriate arteries projecting into the basal ganglia
- Type 2 are located in the cortex following the path of the medullary arteries
- Type 3 are located in the midbrain
Perivascular spaces are most commonly located in the basal ganglia and white matter of the cerebrum, and along the optic tract.[13] The ideal method used to visualize perivascular spaces is T2-weighted MRI. The MR images of other neurological disorders can be similar to those of the dilated spaces. These disorders are:[7]
- cystic neoplasms
- lacunar infarctions
- cystic periventricular leukomalacia
- cryptococcosis
- multiple sclerosis
- mucopolysaccharidoses
- neurocysticercosis
- arachnoid cysts
- neuroepithelial cysts
Perivascular spaces are distinguished on an MRI by several key features. The spaces appear as distinct round or oval entities with a signal intensity visually equivalent to that of cerebrospinal fluid in the subarachnoid space.[7][14][15] In addition, a perivascular space has no mass effect and is located along the blood vessel around which it forms.[14]
Function
One of the most basic roles of the perivascular space is the regulation of fluid movement in the central nervous system and its drainage.[13] The spaces ultimately drain fluid from neuronal cell bodies to the cervical lymph nodes.[5] In particular, the “tide hypothesis” suggests that the cardiac contraction creates and maintains pressure waves to modulate the flow to and from the subarachnoid space and the perivascular space.[16] By acting as a sort of sponge, they are essential for signal transmission and the maintenance of extracellular fluid.[16]
Another function is as an integral part of the blood–brain barrier (BBB).[17] While the BBB is often described as the tight junctions between the endothelial cells, this is an oversimplification that neglects the intricate role that perivascular spaces take in separating the venous blood from the parenchyma of the brain. Often, cell debris and foreign particles, which are impermeable to the BBB will get through the endothelial cells, only to be phagocytosed in the perivascular spaces. This holds true for many T and B cells, as well as monocytes, giving this small fluid filled space an important immunological role.[17]
Perivascular spaces also play an important role in immunoregulation; they not only contain interstitial and cerebrospinal fluid, but they also have a constant flux of macrophages, which is regulated by blood-borne mononuclear cells, but do not pass the basement membrane of the glia limitans.[17] Similarly, as part of its role in signal transmission, perivascular spaces contain vasoactive neuropeptides (VNs), which, aside from regulating blood pressure and heart rate, have an integral role in controlling microglia.[18] VNs serve to prevent inflammation by activating the enzyme adenylate cyclase which then produces cAMP. The production of cAMP aids in the modulation of auto-reactive T cells by regulatory T cells. .[19] The perivascular space is susceptible space for VN compromise and when their function is reduced in the space, immune response is adversely affected and the potential for degradation increases.[18][19] When inflammation by T cells begins, astrocytes begin to undergo apoptosis, due to their CD95 receptor, to open up the glia limitans and let T cells into the parenchyma of the brain.[17] Because this process is aided by the perivascular macrophages, these tend to accumulate during neuroinflammation and cause dilation of the spaces.[18]
Clinical significance
The clinical significance of perivascular spaces comes primarily from their tendency to dilate. The importance of dilation is hypothesized to be based on changes in shape rather than size.[13] Enlarged spaces have been observed most commonly in the basal ganglia, specifically on the lenticulostriate arteries. They have also been observed along the paramedial mesencephalothalamic artery and the substantia nigra in the mesencephalon, the brain region below the insula, the dentate nucleus in the cerebellum, and the corpus callosum, as well as the brain region directly above it, the cingulate gyrus.[5] Upon the clinical application of MRI, it was shown in several studies that perivascular space dilation and lacunar strokes are the most commonly observed histological correlates of signaling abnormalities.[13]
Senescence
Dilation is most commonly and closely associated with aging. Dilation of perivascular spaces has been shown to correlate best with age, even when accompanying factors including hypertension, dementia, and white matter lesions are considered.[20] In the elderly, such dilation has been correlated with many symptoms and conditions that often affect the arterial walls, including vascular hypertension, arteriosclerosis, reduced cognitive capacity, dementia, and low post-mortem brain weight.[13] In addition to dilation among the elderly, dilation in young, healthy individuals can also be observed. This occurrence is rare and there has been no observed association in such cases with reduced cognitive function or white matter abnormalities.[13] When dilated VRS are observed in the corpus callosum, there is generally no neurological deficit associated. They are often observed in this region as cystic lesions with cerebrospinal-like fluid.[21]
Symptoms of dilation
Extreme dilation has been associated with several specific clinical symptoms. In cases of severe dilation in only one hemisphere, symptoms reported include a non-specific fainting attack, hypertension, positional vertigo, headache, early recall disturbances, and hemifacial tics. Symptoms associated with severe bilateral dilation include ear pain (which was reported to have resolved on its own), dementia, and seizures. This data was compiled from case studies of individuals with severe VRS dilation.[13] Considering the anatomical abnormality presented in such cases, these findings were considered surprising in that the symptoms were relatively mild. In most cases, there is in fact no mass effect associated with some VRS dilation. An exception to the mildness of clinical symptoms associated with VRS dilation is when there is extreme dilation in the lower mesencephalon at the junction between the substantia nigra and cerebral peduncle. In such cases, mild to moderate obstructive hydrocephalus was reported in most patients. Associated symptoms ranged from headaches to symptoms more severe than those just discussed in the cases of dilation in the cerebral hemispheres.[13] Other general symptoms associated with VRS dilation include headaches, dizziness, memory impairment, poor concentration, dementia, visual changes, oculomotor abnormality, tremors, seizures, limb weakness, and ataxia.[5]
Associated disorders
Dilation is a typical characteristic of several diseases and disorders. These include diseases from metabolic and genetic disorders such as mannosidosis, myotonic dystrophy, Lowe syndrome, and Coffin–Lowry syndrome. Dilation is also a common characteristic of diseases or disorders of vascular pathologies, including CADASIL (cerebral autosomal dominant arteriopathy with subcortical infarcts and leukoencephalopathy), hereditary infantile hemiparesis, retinal arteriolar tortuosity and leukoencephalopathy, migraines, and vascular dementia. A third group of disorders typically associated with VRS dilation is neuroectodermal syndromes. This includes polycystic brains associated with ectodermal dysplasia, frontonasal dysplasia, and Joubert syndrome. There is a fourth miscellaneous group of disorders typically associated with dilation that includes autism in children, megalencephalopathy, secondary Parkinson’s disease, recent-onset multiple sclerosis, and chronic alcoholism. Because dilation can be associated with several diseases but also observed in healthy patients, it is always important in the evaluation of VRS to study the tissue around the dilation via MRI and to consider the entire clinical context.[13]
Current research
Causes of dilated VRS
Much of the current research concerning Virchow–Robin spaces relates to their known tendency to dilate. Research is presently being performed in order to determine the exact cause of dilation in these perivascular spaces. Current theories include mechanical trauma resulting from cerebrospinal fluid pulsation, elongation of ectactic penetrating blood vessels, and abnormal vascular permeability leading to increased fluid exudation. Further research has implicated shrinkage or atrophy of surrounding brain tissue, perivascular demyelination, coiling of the arteries as they age, altered permeability of the arterial wall and obstruction of lymphatic drainage pathways.[13] In addition, insufficient fluid draining and injury to ischemic perivascular tissue resulting in an ex vacuo effect have been suggested as possible causes for dilated VRS.[5]
Association of dilated VRS and other diseases
Recent and ongoing research has found associations between enlarged VRS and several disorders.
Dementia
At one point in time, dilated Virchow–Robin spaces were so commonly noted in autopsies of persons with dementia, they were believed to cause the disease. However, additional research is currently being performed in order to confirm or refute a direct connection between dilation of VRS and dementia.[15]
Analysis of VRS may distinguish dementia caused by arteriosclerotic microvascular disease from dementia caused by neurodegenerative disease. A 2005 study has evidenced that a substantial amount of VRS in the substantia innominata, lentiform nucleus, and the caudate nucleus of the basal ganglia may implicate dementia due to arteriosclerotic microvascular disease, in particular Ischemic Vascular Dementia, as opposed dementia due to neurodegenerative disease, specifically Alzheimer’s disease and frontotemporal dementia. Thus, perhaps VRS dilation can be used to distinguish between diagnoses of vascular dementias and degenerative dementias.[22]
Alzheimer's disease
Some studies have assessed the spatial distribution and prevalence of VRS in people with Alzheimer's disease versus those without the disease. Researchers have found that while VRS appear to be correlated with natural aging, MR imaging reveals a greater prevalence of VRS in those with Alzheimer's.[23]
Cerebral amyloid angiopathy (CAA), a blood vessel failure often associated with Alzheimer's disease, utilizes dilated VRS to spread inflammation to the parenchyma. Because the VRS often have an extra membrane in gray matter, the ischemic CAA response is often observed in white matter.[24]
It has been hypothesized that the structure of VRS in the cerebral cortex may contribute to the development of Alzheimer’s disease. In contrast to VRS of the basal ganglia, VRS in the cerebral cortex are surrounded by only one layer of leptomeninges. As such, VRS in the cerebral cortex may drain β-amyloid in interstitial fluid less effectively than VRS in the basal ganglia. The less-effective drainage may lead to the development of the β-amyloid plaques that characterize Alzheimer’s disease. In support of this hypothesis, studies have noted the greater frequency of β-amyloid plaques in the cerebral cortex than in the basal ganglia of Alzheimer’s disease patients.[8]
Stroke
Because dilated perivascular spaces are so closely correlated with cerebrovascular disease, there is much current research on their use as a diagnostic tool. In a recent study of 31 subjects, abnormal dilation, along with irregular CSF pulsation, were correlated with those subjects having three or more risk factors for strokes. Therefore, perivascular spaces are a possible novel biomarker for hemorrhagic strokes.[25]
CADASIL syndrome (cerebral autosomal dominant arteriopathy with subcortical infarcts and leukoencephalopathy syndrome) is a hereditary stroke condition due to a Notch 3 gene mutation on Chromosome 19. Studies have noted that in comparison to family members lacking the affected haplotype that leads to the condition, an increased number of dilated spaces is observed in individuals with CADASIL. These perivascular spaces are localized primarily in the putamen and temporal subcortical white matter and they appear to correlate with age of the individual with the condition rather than severity of the disease itself.[26]
There has been a high risk of stroke associated with dilated perivascular spaces in the elderly according to the Framingham Stroke Risk Score.[15] In contrast, other studies have concluded that the dilation of these spaces is a normal phenomenon in aging with no association with arterosclerosis. This remains, therefore, an important point of research in the field.[13]
Multiple sclerosis
Similar to the research concerning a potential connection between perivascular spaces and Alzheimer's, MRI scans of people recently diagnosed with multiple sclerosis (MS) have been studied. Larger, more prevalent spaces have been observed in those with MS.[27] Additional studies with similar findings have suggested that the inflammatory cells which contribute to the demyelination that characterizes MS also attack the perivascular spaces. Studies using advanced MRI techniques will be necessary to determine if the perivascular spaces can be implicated as a potential marker of the disease.[28]
Autism
Dilated perivascular spaces are common among the elderly and uncommon in children. Studies have noted the association between both developmental delay and non-syndromic autism and enlarged or dilated perivascular spaces.[29][30] Non-syndromic autism categorizes autistic patients for which there is no known cause.[29]
History
The appearance of perivascular spaces was first noted in 1843 by Durant-Fardel.[7] In 1851, Rudolph Virchow was the first to provide a detailed description of these microscopic spaces between the outer and inner/middle lamina of the brain vessels. Charles-Philippe Robin confirmed these findings in 1859 and was the first to describe the perivascular spaces as channels that existed in normal anatomy. The spaces were called Virchow-Robin spaces and are still also known as such. The immunological significance was discovered by Wilhelm His, Sr. in 1865 based on his observations of the flow of interstitial fluid over the spaces to the lymphatic system.[13]
For many years after Virchow-Robin spaces were first described, it was thought that they were in free communication with the cerebrospinal fluid in the subarachnoid space. It was later shown with the use of electron microscopy that the pia mater serves as separation between the two. Upon the application of MRI, measurements of the differences of signal intensity between the perivascular spaces and cerebrospinal fluid supported these findings.[13] As research technologies continued to expand, so too did information regarding their function, anatomy and clinical significance.
References
- Norrving, Bo (2016). "Lacunar Syndromes, Lacunar Infarcts, and Cerebral Small-vessel Disease". Stroke. Elsevier. pp. 449–465.e4. doi:10.1016/b978-0-323-29544-4.00027-x. ISBN 978-0-323-29544-4.
Perivascular spaces are fluid-filled spaces that follow a typical course of a vessel penetrating/transversing the brain through gray or white matter.89
- Gross PM, Weindl A (1987). "Peering through the windows of the brain (Review)". Journal of Cerebral Blood Flow and Metabolism. 7 (6): 663–72. doi:10.1038/jcbfm.1987.120. PMID 2891718.
{{cite journal}}: CS1 maint: uses authors parameter (link) - Shaver, SW; Pang, JJ; Wainman, DS; Wall, KM; Gross, PM (1992). "Morphology and function of capillary networks in subregions of the rat tuber cinereum". Cell and Tissue Research. 267 (3): 437–48. doi:10.1007/bf00319366. PMID 1571958. S2CID 27789146.
- Esiri, MM; Gay, D (1990). "Immunological and neuropathological significance of the Virchow–Robin space". Journal of the Neurological Sciences. 100 (1–2): 3–8. doi:10.1016/0022-510X(90)90004-7. PMID 2089138. S2CID 39929713.
- Fayeye, Oluwafikay; Pettorini, Benedetta Ludovica; Foster, Katharine; Rodrigues, Desiderio (2010). "Mesencephalic enlarged Virchow–Robin spaces in a 6-year-old boy: a case-based update". Child's Nervous System. 26 (9): 1155–1160. doi:10.1007/s00381-010-1164-4. PMID 20437240. S2CID 6794261.
- Zhang, E.T.; Inman, C.B.; Weller, R.O. (1990). "Interrelationships of the pia mater and the perivascular (Virchow–Robin) spaces in the human cerebrum". Journal of Anatomy. 170: 111–123. PMC 1257067. PMID 2254158.
- Kwee, Robert M.; Kwee, Thomas C. (2007). "Virchow–Robin Spaces at MR Imaging". RadioGraphics. 27 (4): 1071–1086. doi:10.1148/rg.274065722. PMID 17620468.
- Pollock, H.; Hutchings, M.; Weller, R.O.; Zhang, E.T. (1997). "Perivascular spaces in the basal ganglia of the human brain :their relationship to lacunes". Journal of Anatomy. 191 (3): 337–346. doi:10.1046/j.1469-7580.1997.19130337.x. PMC 1467691. PMID 9418990.
- Kendall, M. D (1989). "The morphology of perivascular spaces in the thymus". Thymus. 13 (3–4): 157–64. PMID 2694455.
- Edelman, E. R; Nugent, M. A; Karnovsky, M. J (1993). "Perivascular and intravenous administration of basic fibroblast growth factor: Vascular and solid organ deposition". Proceedings of the National Academy of Sciences of the United States of America. 90 (4): 1513–1517. Bibcode:1993PNAS...90.1513E. doi:10.1073/pnas.90.4.1513. PMC 45904. PMID 8434012.
- Morrison, S. J; Scadden, D. T (2014). "The bone marrow niche for haematopoietic stem cells". Nature. 505 (7483): 327–334. Bibcode:2014Natur.505..327M. doi:10.1038/nature12984. PMC 4514480. PMID 24429631.
- Hatton, G. I (1988). "Pituicytes, glia and control of terminal secretion" (PDF). The Journal of Experimental Biology. 139: 67–79. doi:10.1242/jeb.139.1.67. PMID 3062122.
- Groeschel, S.; Chong, WK.; Surtees, R.; Hanefeld, F. (2006). "Virchow–Robin spaces on magnetic resonance images: normative data, their dilatation, and a review of the literature" (PDF). Neuroradiology. 48 (10): 745–754. doi:10.1007/s00234-006-0112-1. PMID 16896908. S2CID 31816844.
- Ogawa, Toshihide; Okudera, Toshio; Fukasawa, Hitoshi; Hashimoto, Manabu; Inugami, Atsushi; Fujita, Hideaki; Hatazawa, Jun; Noguchi, Kyo; Uemura, Kazuo; Nakajima, Shigeyoshi; Yasui, Nobuyuki (1995). "Unusual Widening of Virchow–Robin Spaces: MR Appearance". American Journal of Neuroradiology. 16 (6): 1238–1242. PMC 8337819. PMID 7677015.
- Mills, S.; Cain, J.; Purandare, N.; Jackson, A. (2007). "Biomarkers of cerebrovascular disease in dementia". British Journal of Radiology. 80: S128–S145. doi:10.1259/bjr/79217686. PMID 18445743.
- Agnati, L.F.; Genedani, S.; Lenzi, P.L.; Leo, G.; Mora, F.; Ferré, S.; Fuxe, K. (2005). "Energy gradients for the homeostatic control of brain ECF composition and for VT signal migration: introduction of the tide hypothesis". Journal of Neural Transmission. 112 (1): 45–63. doi:10.1007/s00702-004-0180-5. PMID 15599604. S2CID 24193688.
- Bechmann, Ingo; Galea, Ian; Perry, V Hugh (2007). "What is the blood–brain barrier (not)?" (PDF). Trends in Immunology. 28 (1): 5–11. doi:10.1016/j.it.2006.11.007. PMID 17140851.
- Pantoni, Leonardo (2010). "Cerebral small vessel disease: from pathogenesis and clinical characteristics to therapeutic challenges". Lancet Neurol. 9 (7): 689–701. doi:10.1016/S1474-4422(10)70104-6. PMID 20610345. S2CID 27010799.
- Staines, D.R.; Brenu, E.W.; Marshall-Gradisnik, S. (2008). "Postulated role of vasoactive neuropeptide-related immunopathology of the blood brain barrier and Virchow–Robin spaces in the aetiology of neurological-related conditions". Mediators of Inflammation. 2008: 1–5. doi:10.1155/2008/792428. PMC 2643053. PMID 19229345.
- Davis, Patricia C.; Mirra, Suzanne S.; Alazraki, Naomi (1994-06-01). "The brain in older persons with and without dementia: Findings on MR, PET, and SPECT images". American Journal of Roentgenology. 162 (6): 1267–78. doi:10.2214/ajr.162.6.8191980. PMID 8191980.
- Uchino, A.; Takase, Y.; Nomiyama, K.; Egashira, R.; Kudo, S. (2005). "Acquired lesions of the corpus callosum: MR imaging". European Radiology. 16 (4): 905–14. doi:10.1007/s00330-005-0037-9. PMID 16284771. S2CID 10484243.
- Patankar, Toufail F.; Mitra, Dipayan; Varma, Anoop; Snowden, Julie; Neary, David; Jackson, Alan (2005). "Dilation of the Virchow–Robin Space Is a Sensitive Indicator of Cerebral Microvascular Disease: Study in Elderly Patients with Dementia". American Journal of Neuroradiology. 26 (6): 1512–1520. PMC 8149063. PMID 15956523.
- Chen, W.; Song, X.; Zhang, Y. (2011). "Assessment of the Virchow–Robin Spaces in Alzheimer Disease, Mild Cognitive Impairment, and Normal Aging, Using High-Field MR Imaging". American Journal of Neuroradiology. 32 (8): 1490–5. doi:10.3174/ajnr.A2541. PMC 7964361. PMID 21757525.
- Schrag, M; McAuley, G; Pomakian, J; Jiffry, A; Tung, S; Mueller, C; Vinters, HV; Haacke, EM; Holshouser, B; Kido, D; Kirsch, WM (2010). "Correlation of hypointesities in susceptibility-weighted images to tissue histology in dementia patients with cerebral amyloid angiopathy: a postmortem MRI study". European Radiology. 119 (3): 291–302. doi:10.1007/s00401-009-0615-z. PMC 2916065. PMID 19937043.
- Selvarajah, J; Scott, M; Stivaros, S; Hulme, S; Georgiou, R; Rothwell, N; Tyrrell, P; Jackson, A (2008). "Potential surrogate markers of cerebral microvascular angiopathy in asymptomatic subjects at risk of stroke". European Radiology. 19 (4): 1011–1018. doi:10.1007/s00330-008-1202-8. PMID 18987865. S2CID 417275.
- Cumurciuc, R.; Guichard, J.-P.; Reizine, D.; Gray, F.; Bousser, M. G.; Chabriat, H. (2006). "Dilation of Virchow–Robin spaces in CADASIL". European Journal of Neurology. 13 (2): 187–190. doi:10.1111/j.1468-1331.2006.01113.x. PMID 16490051. S2CID 23224811.
- Etemadifar, M.; Hekmatnia, A.; Tayari, N.; Kazemi, M.; Ghazavi, A.; Akbari, M.; Maghzi, A. (2011). "Features of Virchow–Robin spaces in newly diagnosed multiple sclerosis patients". European Journal of Radiology. 80 (2): 104–8. doi:10.1016/j.ejrad.2010.05.018. PMID 20650586.
- Achiron, A.; Faibel, M. (2002). "Sandlike appearance of Virchow–Robin spaces in early multiple sclerosis: a novel neuroradiologic marker". American Journal of Neuroradiology. 23 (3): 376–80. PMC 7975312. PMID 11901003.
- Boddaert, Nathalie; Zilbovicius, Mônica; Philipe, Anne; Robel, Laurence; Bourgeois, Marie; Barthélemy, Catherine; Seidenwurm, David; Meresse, Isabelle; Laurier, Laurence; Desguerre, Isabelle; Bahi-Buisson, Nadia; Brunelle, Francis; Munnich, Arnold; Samson, Yves; Mouren, Marie-Christine; Chabane, Nadia (2009). "MRI Findings in 77 Children with Non-Syndromic Autistic Disorder". PLOS ONE. 4 (2): e4415. Bibcode:2009PLoSO...4.4415B. doi:10.1371/journal.pone.0004415. PMC 2635956. PMID 19204795.
- Zeegers, Mijke; Van Der Grond, Jeroen; Durston, Sarah; Jan Nievelstein, Rutger; Witkamp, Theo; Van Daalen, Emma; Buitelaar, Jan; Van Engeland, Herman (2006). "Radiological findings in autistic and developmentally delayed children". Brain and Development. 28 (8): 495–9. doi:10.1016/j.braindev.2006.02.006. PMID 16616445. S2CID 41735789.