ARN CRISPR transactivante (ARNtracr)
El ARN CRISPR transactivante o ARNtracr es un fragmento de ARN que interactúa con el sistema de Repeticiones Palindrómicas Cortas Agrupadas y Regularmente Interespaciadas (CRISPR). De CRISPR hay muchos tipos, pero nos centraremos en el tipo 2, en el que el tracrRNA usa una secuencia complementaria a este de manera que se permite la maduración del ARNcr con la ayuda del Cas9. El CRISPR proporciona inmunidad adaptiva o adquirida a bacterias y arqueas y el uso de la tecnología con este sistema está siendo muy útil para la edición del genoma[1][2]. Dentro del ARNtracr encontramos una clasificación que divide en tres grupos las bacterias con un sistema inmunológico CRISPR tipo 2 ya que presentan unos prototipos ligeramente distintos a la estructura original dependiendo de la familia a la que pertenecen[3].
| Género de bacterias | Bacterias | Estructura ARNtracr | |
|---|---|---|---|
| Streptococcus | S. sanguinis, S. oralis, S. mitis, S. anginosus B, S. pyogenes, S. dysagalactiae, S. equí, S. thermophilus CRISPR3, S. salivarius, S. gallolycticus, S. lutetiensis |  |
|
| Streptococcus | S. henryi, S. infantarius, S. orisratti, S. vestibularis, S. intermedius, S. mutans, S. thermophilus CRISPR1, S. anginosus A, S. gordonii, S. parasanguinis |  |
|
| Lactobacillus | L plantarum, L. pentosus, L. reuteri, L. curvatus, L. fermentum, L. paracasei, L. casei, L. rhamnosus, L. brevis, L. mucosae, L. buchneri A, L. gasseri, L. hominis, L. johnsonii, L. jensenii, L. rossiae, L. delbrueckii, L. coryniformis, L. buchneri B, L. otakiensis |  |
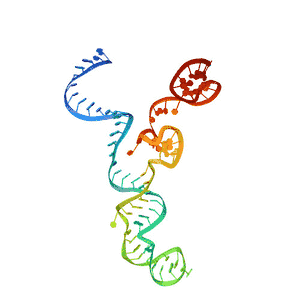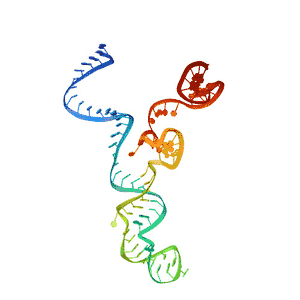
| ARN CRISPR transactivante | |
|---|---|
| Identificadores | |
| Organismo | |
| Símbolo | ARNtracr |
| Entrez | https://www.ncbi.nlm.nih.gov/gene/3665270 |
| PDB | (sgARN) 4ZT9 (sgARN) |
| RefSeq (mRNA) | (sgARN) 31603896 (sgARN) |
| Otros datos | |
Biosíntesis y genética
Adaptación
El bacteriófago se une a la membrana plasmática de una bacteria (p. ej.:Streptococcus pyogenes) e introduce su material genético o ADN bicatenario al citoplasma celular. Esto conlleva toda una serie de transducción de señales que activan las RNasas (Nucleasas) Cas1 y Cas2[4], y detectan una secuencia de nucleótidos específica del ADN vírico bicatenario, también conocido como protoespaciador, gracias a una secuencia PAM (Protospacer adjacent motif) que la precede.
Tras la localización de esta secuencia específica, las RNasas Cas1 y Cas2 llevan a cabo el proceso de integración del material genético vírico al ADN bicatenario de la bacteria (Este proceso también es conocido como fase de adaptación[5][6][7]). Esta secuencia se convierte en un espaciador característico del virus en el ADN bacteriano con una secuencia CRISPR (clustered regularly interspaced short palindromic repeats) adyacente. El conjunto de espaciadores y secuencias CRISPR que se encuentran en el genoma procariota se denomina matriz CRISPR.
Esta matriz CRISPR, en el caso de la bacteria Streptococcus pyogenes, como se trata de un sistema Tipo II-A, estará formada por cinco espaciadores cuyos objetivos serán la endopeptidasa, el superantígeno (speM), la metiltransferasa, la hialuronidasa y una proteína hipotética del fago o bacteriófago[7]. Le sigue, aguas arriba (en sentido 5'), una secuencia líder rica en secuencia AT[8] (Adenina y Timina) que delimita el inicio de la matriz y la separa del operón Cas; formado por los genes codificantes de las proteínas (nucleasas) Cas9, Cas1, Cas2 y Csn2[9]. Seguidamente, se encuentra la secuencia del promotor de transcripción y, finalmente, el gen codificante del ARNtracr; subdividido en una región que codifica para el ARNtracr largo o tracr-L, y en otra que codifica para el ARNtracr corto o tracr-S.
.jpg.webp)
Expresión
Ante la falta de investigación sobre este mecanismo, aún no data un sistema de transcripción concreto que determine qué inicia la biosíntesis del ARNtracr. No obstante, se ha tratado de demostrar la coexistencia de dos tipos de sistemas que pueden llevar a cabo la biosíntesis del ARNtracr.
Por un lado, según un estudio basado en la constante observación de la expresión y el procesamiento de los ARNtracr, a raíz de un sondeo de ARN de varias bacterias (Neisseria meningitidis, Streptococcus mutans y Streptococcus thermophilus), se determinó que los respectivos sistemas CRISPR/Cas9 de tipo II se encuentran constitutivamente activos. Como consecuencia, las bacterias que gozan de este sistema presentan una alta respuesta inmunológica ante bacteriófagos que las infecte[10].
No obstante, por otro lado, sí que existen otros tipos de bacterias dotadas de un represor del promotor de transcripción del locus CRISPR. Este sistema es llevado a cabo por un tipo de ARNtracr salvaje de algunas bacterias (ARNtracr largo o tracr-L) que se pliega en forma de ARNg[6] y se une a una proteína Cas9[10]. Este, al encontrarse libre, mediante la complementariedad de bases nitrogenadas de una pequeña región de la molécula respecto a una corta secuencia PAM del promotor de transcripción, se une al locus CRISPR y silencia la transcripción (silenciamiento génico transcripcional o TGS) del locus. Por esta misma razón, el tracr-L es capaz de regular la síntesis de las proteínas Cas y del resto de genes del locus[6]. De este modo, cuando una célula, en este caso Streptococcus pyogenes, es infectada por un bacteriófago, el represor transcripcional (tracr-L) se libera y permite la activación del promotor para iniciar la transcripción del locus CRISPR. Por esta misma razón, a diferencia del sistema constitutivo de algunas bacterias, la importancia del tracr-L radica en la significativa reducción de probabilidades de padecer enfermedades autoinmunitarias.
Tal y como se representa en la FIGURA 1, la transcripción del operón Cas9 y la matriz CRISPR va en sentido 5'-3', es decir, que la síntesis de ARN es 3'-5'; mientras que la transcripción del gen ARNtracr, que finaliza gracias a la presencia de una Terminación independiente de Rho, es 3'-5', de manera que la biosíntesis del ARNtracr será 5'-3' (en excepción de un gen ARNtracr del sistema CRISPR/Cas9 Tipo II-C)[11].
Como resultado de la transcripción y expresión de genes, se sintetizan un conjunto de ARN's y proteínas que desarrollan la respuesta inmunológica, como ahora el tracr-P (tracr-S procesado o transcrito), las RNasas Cas9, Cas1, Cas2 y Csn2 y el pre-ARNcr (pre-ARN CRISPR).
Localizaciones del gen ARNtracr
Como se ha especificado anteriormente, este caso se centra en el sistema CRISPR/Cas9 Tipo II-A del Streptococcus pyogenes. El estudio de los diferentes sistemas CRISPR ha conllevado la aparición de una gran diversidad de sistemas de respuesta inmunitaria de las diferentes bacterias dotadas del sistema CRISPR/Cas9. Por ende, se presenta una gran la variabilidad de los genes codificantes[11][12]. En otros sistemas CRISPR/Cas Tipo II, como el Tipo II-A del S. thermophilus, el gen codificante del ARNtracr tanto se encuentra aguas arriba del gen Cas9 como justo después de su secuencia. En el caso del Tipo II-B del Francisella Novicida, el gen se encuentra adyacente al operón Cas9 y aguas arriba de la matriz CRISPR[13]. O, en el caso del Tipo II-C del Campylobacter jejuni, que la localización de sus genes ARNtracr se basa en la unión de los modelos de los Tipos II-A y II-B.
Estructura y composición química

El ARNtracr es un pequeño ARN no codificante formado por entre 50 y 171 nucleótidos, por lo que tiene un masa molar de entre 16500-49500 dalton. El número de nucleótidos puede variar tras la maduración del ARNcr (transcrito del gen CRISPR) en la que interviene, ya que puede ser escindido por la RNAsa III la cual regula la maduración del transcrito.[14]
En cuanto a su composición química, el ARNtracr, está formado por la polimerización de nucleótidos mediante la combinación de las bases nitrogenadas A-U, G-C. Se ha establecido un modelo, llamado andamio (scaffold), que establece las bases nitrogenadas que tiene el ARNtracr, aunque puede contener otras dependiendo de la familia de la bacteria. [3]
El modelo base de unión al ARNcr es el siguiente:
5´AAACAGCAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGCUUCG-3’ [15][10]
Alrededor de los primeros 24-25 nucleótidos corresponden a la secuencia de anterrepetición parcialmente complementaria a la secuencia repetidora del ARNcr, a la que se une formando un ARN híbrido conocido como ARN guía o ARNg que se une a la proteína Cas9 para dirigirla hasta el ADN vírico ambas cadenas son necesarias para llevar a cabo esta acción.[10][16]
La estructura del ARNtracr está definida por 3 loops. En el primer loop se encuentra la secuencia de antirrepetición y el linker de unión al siguiente tramo. Esta zona presenta gran afinidad por la proteína Cas9, en concreto por el dominio Hel-I, rica en arginina, y el dominio CTD. El segundo loop mantiene muchos menos contacto con la proteína y tan solo presenta cierta afinidad por los dominios RuvC y CTD. Por último, en el tercer loop no se ha encontrado ningún tipo de unión con la proteína porque para observar su estructura por cristalografía se ha de prescindir de este.[10] En una visión global, se observa, por tanto, que el sgARN se encuentra entre los dos lóbulos de la proteína Cas9.[17]
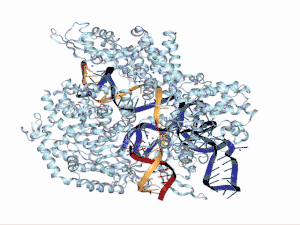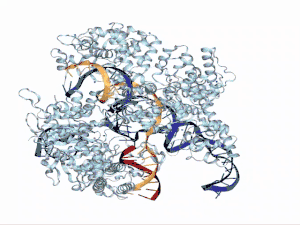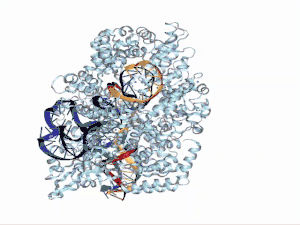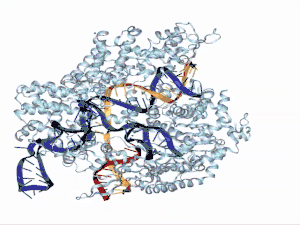
Funciones en el sistema CRISPR/Cas9
El ARNtracr media la interacción entre ARNcr y Cas9, endonucleasa asociada a los sistemas CRISPR del tipo II, convirtiéndose así en uno de los componentes esenciales en el sistema inmunitario procariota. El sistema CRISPR II es el que se ha utilizado mayormente como herramienta de edición genética por su simple mecanismo en el cual participan tres componentes siendo el ARNtracr uno principal de ellos junto con la proteína Cas 9 y el ARNcr. La nucleasa Cas9 utiliza como guía a un ARNcr que se aparea con el ARN transactivante, el cual cumple una función principalmente estructural.[18]
Esta función esencial involucrada en la conformación de la estructura del complejo CRISPR se puede desglosar en tres funciones principales; participa en la adquisición del espaciador, dirige la maduración de otro ARN no codificante (pre-ARNcr) mediante la actividad de la enzima RNAasa III para producir la especie activa (ARNcr maduros). [19][20] Y, por otro lado, ARNtracr también es necesario para la activación y posterior guía de la nucleasa Cas9 para que realice el corte en el ADN diana proveniente del material genético del virus (bacteriófago) inactivándolo y protegiendo a la célula procariota de agentes patógenos invasores.[21]
Mecanismos de reacción en el sistema CRISPR/Cas9
En general, el funcionamiento del sistema CRISPR que proporciona la inmunidad adquirida en bacterias y arqueas, se puede dividir en tres etapas bien diferenciadas: etapa de adquisición, etapa de expresión y etapa de interferencia. [22] Cada uno de los diferentes tipos de sistema CRISPR presenta peculiaridades propias de su biología, pero la ruta de maduración y procesamiento de ARNcr en los sistemas de tipo II, como lo ejemplifica S. pyogenes , difiere significativamente. Se ha demostrado que el proceso requiere una molécula de ARNtracr adicional y depende de la RNasa III de la célula huésped. [23]
Durante la etapa de adquisición en la que se ha captado ADN foráneo del ente invasor (protoespaciador) y se ha incorporado en el locus CRISPR (espaciador) [24] entre las secuencias repetidas y en posición 5´ respecto a la secuencia espaciadora y siempre tras la secuencia líder, el ARNtracr parece estar involucrado en la adquisición del espaciador guiado por la Cas 9, aunque la base mecánica de esta función sigue siendo difícil de alcanzar. [25][26]
A continuación, en la etapa de expresión, la matriz CRISPR se transcribe dando lugar a un largo transcrito primario, ARN precursor-CRISPR (pre-crRNA), molécula larga de ARN que contiene todos los espaciadores adquiridos en la etapa anterior separados por secuencias repetidoras. [27][28] También se transcribe un segundo ARN procedente de un locus genómico situado aguas arriba del locus CRISPR y de los genes cas; el ARNtracr. [29][30]

El ARNtracr tiene una región complementaria a la secuencia de repetición del locus CRISPR, de manera que se une al pre-RNA recién transcrito debido a su complementariedad de bases de la secuencia repetida (homología en unos 25 nucleótidos aproximadamente) formando un complejo dúplex reconocido y posteriormente escindido por la RNAsa III (una enzima que reconoce y corta el RNA de doble cadena) quien va a llevar a cabo el procesamiento generando el extremo 3′ de los ARNcr maduros. Se tratan de secuencias de ARN constituidas por un único espaciador (secuencia corta variable de ARN, homóloga a las secuencias genómicas de fagos) en su extremo 5’ flanqueada por dos secuencias repetidoras. [29] Sin embargo, queda por establecer si otras nucleasas celulares contribuyen al procesamiento del extremo 5′ del crRNA en los sistemas de tipo II para que el ARNcr intermediario se transfigure en su estructura madura. De esta manera, el ARNcr originalmente con dos secuencias repetidoras pasa a tener exclusivamente una, facilitando así su afinidad y unión al ARNtracr y consecuentemente, la guía al ADN invasor para degradarlo. Por otro lado, la RNasa III no es necesaria para el procesamiento de un transcrito de crRNA "mínimo" que incluye un único espaciador. ARNtracr, por lo tanto, dirige la maduración de precr-RNA promoviendo la escisión endonucleotídica dando lugar a ARNcr maduros.
Seguidamente, el híbrido ARNcr:ARNtracr se asocia con una proteína Cas9, creando un complejo de ribonucleoproteína activo (RNP). Aun así, queda por establecer si la contribución de Cas9 se debe al papel activo en el reconocimiento de ARNcr y ARNtracr, o a la capacidad de eliminar estructuras secundarias en las moléculas de ARNcr y ARNtracr monocatenarias. Cabe destacar que el extremo 3' de ARNtracr puede contribuir a la unión de Cas9, demostrando una vez más su importancia en los sistemas CRISPR del tipo II. Una vez que el ARNcr:ARNtracr se une a Cas9, esta se activa y puede escindir las secuencias de ácido nucleico invasoras de tal forma que ARNtracr constituye una pieza fundamental para la activación de la enzima endonucleasa.[28]
Evidencias experimentales tanto in vitro como in vivo demuestran que este ARN dual que dirige a Cas9 para escindir cualquier ADN debe de contener al menos 15 nt (nucleótidos) más allá de la secuencia complementaria de ARNcr y PAM adyacente. Así entonces, el recorte en el extremo 3' de ARNtracr compromete la actividad de escisión del ADN, lo que sugiere que el fragmento de ARNtracr cerca de la unión dúplex puede ser crítico para la interacción con la proteína Cas9[31] para el ensamblaje del complejo catalíticamente activo. Dichos estudios corroboran que el dúplex repetido-antirepetido y el primer tallo-bucle son indispensables para la formación del complejo Cas9-sgRNA, mientras que el enlazador, el segundo y tercer bucle-tallo no son necesarios para la función pero pueden estabilizar el ARN guía.
La etapa de interferencia tiene lugar cuando el genoma de un organismo foráneo reincidente intenta de nuevo infectar la célula procariota. En este estadio, el híbrido de ARN (ARNtracr:ARNcr) es imprescindible para dirigir a la endonucleasa Cas9 hacia la secuencia diana del material genómico invasivo que debe degradar. [32][33][34] Para que se produzca la escisión del material genético extraño y por lo tanto, la protección de delante del agente invasor, deben cumplirse dos requisitos indispensables: que exista hibridación mediada por complementariedad de bases (A=T, G≡C) entre el ARNcr y el ADN diana; y que la secuencia diana a cortar se localice inmediatamente adyacente a un PAM aguas arriba o abajo del protoespaciador. [35] La Cas9 realiza el corte 3 pares de bases antes de la PAM, una secuencia de unos pocos nucleótidos, con una composición de bases y tamaño que varía entre sistemas CRISPR/Cas (por ejemplo, para Cas9 de Streptococcus pyogenes -SpCas9- esta secuencia es 5´NGG3´). [36] Finalmente, si el ADN es complementario al ARNg, se forma un híbrido ARN:ADN, denominado bucle R, y se produce la escisión. [37] El corte del ADN es el resultado de la acción de dos dominios diferentes de la nucleasa Cas9: el dominio HNH corta la cadena complementaria al ARNcr y el dominio RuvC corta la cadena que no es complementaria al ARNcr.[38]
Historia y antecedentes
La historia del tracrARN forma parte de la historia del CRISPR. Este fue descubierto por Francisco Mojica en 1992 al observar una repetición de bases en el genoma de la arquea Haloferax mediterranei. En un primer momento, Mojica, no pudo descifrar a que se debían estas repeticiones, de manera que no fue hasta el año 2003 cuando el científico dilucidó que estas se debían al sistema inmunitario del microorganismo y que estaba presente en todas las bacterias.
Tras este descubrimiento, muchos científicos analizaron diferentes familias de bacterias observando distintos tipos de CRISPR. A grandes rasgos, encontramos tres tipos: I,II,II. El ARNtracr tan solo se encuentra en el tipo II. Esta molécula fue descubierta por Emmanuelle Marie Charpentier en colaboración con Jörg Vogel del Grupo de Biología del RNA en el Instituto de Biología Molecular de la Infección en la Universidad de Würzburg, en Würzburg, Alemania, en un estudio que trataba de identificar ARNs microbianos, tras observar uno especialmente virulento en el Streptococcus pyogenes, se utilizó un programa bioinformático que buscaba regiones intergénicas que pudieran regular funciones de la bacteria y que no codificaban para ninguna proteína. En su estudio observó una región cerca del CRISPR que podría llevar esto a cabo. Tras observarla más detenidamente, pudo ver que se trataba del tercer ARN no codificante más abundante y que su función era participar en la maduración del ARNcr y su combinación con este hace que tan solo se necesite una proteína, la Cas 9, para llevar a cabo la respuesta inmunitaria de la bacteria. Fue denominado tracrARN.
En 2011, Charpentier presentó su descubrimiento en una conferencia. Allí conoció a Jennifer Doudna, bióloga estadounidense, quien, junto a Charpentier, ideó el método CRISPR-Cas9 para insertar genes en el genoma de seres vivos, reprogramando el ARNcr para que fuera específico al segmento que se quería cortar. Por ende, finalmente crearon una estructura quimera fusionando las cadenas de ARNcr y ARNtracr (ARNg).
Debido a sus descubrimientos, a ambas científicas se les otorgó el premio nobel de química 2020. [39][40][41][42][43]
Aplicaciones
El ARNtracr interactuando con el CRISPR adaptado es una herramienta biotecnológica de gran potencia. Sus utilidades destacan sobre todo en el campo de la edición genética y diagnóstico molecular. Además de poseer una gran concisión y simplicidad en su diseño, tiene un bajo coste, la cual cosa lo convierte en un método verdaderamente óptimo para en la investigación y edición del genoma. Algunas de las aplicaciones serían:
Diagnóstico molecular
Gracias a que se puede utilizar para reconocer ácidos nucleicos, se pueden detectar mutaciones en los nucleótidos porque se hallaría la diferencia entre la secuencia del sistema CRISPR y la secuencia diana debido a su elevado grado de especificidad. Asimismo, sería posible la detección de patógenos cuando están infectando a través del análisis de su material genético (ARN o ADN) y se podrían diagnosticar enfermedades que dan llugar a nivel molecular con mayor exactitud y seguridad, como sería el caso del cáncer[44][45][46].
Sistema LEOPARD ( "Leveraging Engineered tracrRNAs and On-target DNAs for PArallel RNA Detection") para el tratamiento de la COVID-19

Otro ejemplo sería el sistema LEOPARD[47][48] que, mediante el uso de los ARNtracr y el ADN, el ARN paralelo es detectado. Está programado para que cuando se detecte cualquier ARN, el ADN correspondiente sea cortado por Cas9. De esta manera, este método de diagnóstico CRISPR permite detectar múltiples biomarcadores al mismo tiempo. Es este método el que distinguió el síndrome respiratorio agudo severo coronavirus 2 (SARS-CoV-2). En resumen, el ARNtracr es reprogramado para decidir los ARN que serán los ARN guía y, dependiendo de qué ADN corta, se puede determinar los ARN que estaban presentes en la muestra.
Bloqueo de la expresión del gen VIH1
También es un método muy esperanzador para enfermedades para las que no se ha encontrado aún una cura definitiva, como es el caso del VIH. A pesar de haber desarrollado terapias para controlar la replicación del virus VIH-1, el virus no desaparece sino que se mantiene en un estado de latencia en el genoma de la célula huésped. Por consiguiente, continúa presente la amenaza de que al cabo de un tiempo resurja. La solución que se ofrece con el CRISPR/Cas9 es que este, potencialmente, pueda modificar el genoma del VIH-1 así como impedir su expresión.
Se ha puesto a prueba la capacidad del CRISPR/Cas9 para anular la expresión del virus VIH-1 editando el ADN proviral integrado. Se dirigen los componentes CRISPR/Cas9 hacia el LTR, que son secuencias terminales genéticas de su genoma que regulan la integración del ácido nucleico viral en los cromosomas de la célula huésped y que, por lo tanto, influyen en la virulencia de este, y se consiguió inhibir de forma importante la expresión del virus mediada por LTR y también suprimir provirus del genoma celular[49][50].
Cáncer
Se está estudiando un método que tiene como objetivo mejorar la capacidad de las células T humanas para combatir el cáncer. Se basa en quitar dos de los genes que codifican las cadenas del receptor endógeno de las células T (TCR), que son el TCRα y TCRβ, para así reducir al máximo el emparejamiento equívoco de TCR y lograr una mejor expresión de un transgen TCR sintético específico para el cáncer. Asimismo, se elimina otro gen que codifica la proteína de muerte celular programada con el fin de aumentar la inmunidad antitumoral. La transferencia de estas células T modificadas genéticamente a los pacientes con cáncer ha permitido conseguir en un injerto duradero con ediciones en los tres loci genómicos. Estas células T modificadas persistieron durante un periodo de hasta 9 meses, cosa que propone que la inmunogenicidad es mínima en estas condiciones y evidencia la viabilidad de la edición del gen CRISPR para la inmunoterapia contra el cáncer[51].
Referencias
- Faure, Guilhem; Shmakov, Sergey A.; Makarova, Kira S.; Wolf, Yuri I.; Crawley, Alexandra B.; Barrangou, Rodolphe; Koonin, Eugene V. (2019-04). «Comparative genomics and evolution of trans-activating RNAs in Class 2 CRISPR-Cas systems». RNA biology 16 (4): 435-448. ISSN 1555-8584. PMC 6546382. PMID 30103650. doi:10.1080/15476286.2018.1493331. Consultado el 30 de octubre de 2022.
- Workman, Rachael E.; Pammi, Teja; Nguyen, Binh T. K.; Graeff, Leonardo W.; Smith, Erika; Sebald, Suzanne M.; Stoltzfus, Marie J.; Euler, Chad W. et al. (4 de febrero de 2021). «A natural single-guide RNA repurposes Cas9 to autoregulate CRISPR-Cas expression». Cell (en inglés) 184 (3): 675-688.e19. ISSN 0092-8674. doi:10.1016/j.cell.2020.12.017. Consultado el 30 de octubre de 2022.
- Briner, Alexandra E.; Donohoue, Paul D.; Gomaa, Ahmed A.; Selle, Kurt; Slorach, Euan M.; Nye, Christopher H.; Haurwitz, Rachel E.; Beisel, Chase L. et al. (23 de octubre de 2014). «Guide RNA Functional Modules Direct Cas9 Activity and Orthogonality». Molecular Cell (en inglés) 56 (2): 333-339. ISSN 1097-2765. doi:10.1016/j.molcel.2014.09.019. Consultado el 31 de octubre de 2022.
- Rocha, Taibo; Eugenia, María (2016). Caracterización y análisis de la variación de Loci CRISPRs en Streptococcus thermophilus. Consultado el 21 de octubre de 2022.
- Vol. 25 (2022). doi:10.22201/fesz.23958723e.2022. Consultado el 23 de octubre de 2022.
- «Shibboleth Authentication Request». login.sire.ub.edu. doi:10.1146/annurev-genet-071719-022559. Consultado el 23 de octubre de 2022.
- «Shibboleth Authentication Request». login.sire.ub.edu. doi:10.1080/15476286.2019.1582974?needaccess=true. Consultado el 21 de octubre de 2022.
- «ORIGEN Y DESARROLLO DEL SISTEMA CRISPR/Cas. De 1987 a 2002. 1ª parte». www.fernandogalangalan.com. Consultado el 21 de octubre de 2022.
- «Shibboleth Authentication Request». login.sire.ub.edu. doi:10.1038/471588a. Consultado el 23 de octubre de 2022.
- Deltcheva, Elitza; Chylinski, Krzysztof; Sharma, Cynthia M.; Gonzales, Karine; Chao, Yanjie; Pirzada, Zaid A.; Eckert, Maria R.; Vogel, Jörg et al. (31 de marzo de 2011). «CRISPR RNA maturation by trans-encoded small RNA and host factor RNase III». Nature 471 (7340): 602-607. ISSN 0028-0836. PMC 3070239. PMID 21455174. doi:10.1038/nature09886. Consultado el 21 de octubre de 2022.
- Karvelis, Tautvydas; Gasiunas, Giedrius; Miksys, Algirdas; Barrangou, Rodolphe; Horvath, Philippe; Siksnys, Virginijus (1 de mayo de 2013). «crRNA and tracrRNA guide Cas9-mediated DNA interference in Streptococcus thermophilus». RNA Biology 10 (5): 841-851. ISSN 1547-6286. PMC 3737341. PMID 23535272. doi:10.4161/rna.24203. Consultado el 23 de octubre de 2022.
- Liao, Chunyu; Beisel, Chase L. (23 de noviembre de 2021). «The tracrRNA in CRISPR Biology and Technologies». Annual Review of Genetics (en inglés) 55 (1): 161-181. ISSN 0066-4197. doi:10.1146/annurev-genet-071719-022559. Consultado el 23 de octubre de 2022.
- Bikard, David; Marraffini, Luciano A. (1 de noviembre de 2013). «Control of gene expression by CRISPR-Cas systems». F1000Prime Reports 5: 47. ISSN 2051-7599. PMC 3816762. PMID 24273648. doi:10.12703/P5-47. Consultado el 23 de octubre de 2022.
- Deltcheva, Elitza; Chylinski, Krzysztof; Sharma, Cynthia M.; Gonzales, Karine; Chao, Yanjie; Pirzada, Zaid A.; Eckert, Maria R.; Vogel, Jörg et al. (31 de marzo de 2011). «CRISPR RNA maturation by trans-encoded small RNA and host factor RNase III». Nature 471 (7340): 602-607. ISSN 0028-0836. PMC 3070239. PMID 21455174. doi:10.1038/nature09886. Consultado el 24 de octubre de 2022.
- Xie, Kabin; Yang, Yinong (1 de noviembre de 2013). «RNA-Guided Genome Editing in Plants Using a CRISPR–Cas System». Molecular Plant (en inglés) 6 (6): 1975-1983. ISSN 1674-2052. PMID 23956122. doi:10.1093/mp/sst119. Consultado el 24 de octubre de 2022.
- «Shibboleth Authentication Request». login.sire.ub.edu. Consultado el 24 de octubre de 2022.
- Martin Jinek, Fuguo Jiang, David W. Taylor, Samuel H. Sternberg, Emine Kaya, Enbo Ma2, Carolin Anders, Michael Hauer, Kaihong Zhou, Steven Lin, Matias Kaplan, Anthony T. Iavarone, Emmanuelle Charpentier, Eva Nogales, and Jennifer A. Doudna (14 de marzo de 2014). «Structures of Cas9 Endonucleases Reveal RNA-Mediated Conformational Activation». Science. Consultado el 17 de octubre de 2022.
- Faure, Guilhem; Shmakov, Sergey A.; Makarova, Kira S.; Wolf, Yuri I.; Crawley, Alexandra B.; Barrangou, Rodolphe; Koonin, Eugene V. (14 de agosto de 2018). «Comparative genomics and evolution of trans-activating RNAs in Class 2 CRISPR-Cas systems». RNA Biology 16 (4): 435-448. ISSN 1547-6286. doi:10.1080/15476286.2018.1493331. Consultado el 29 de octubre de 2022.
- Emmanuelle, Deltcheva, Elitza Chylinski, Krzysztof Sharma, Cynthia M Gonzales, Karine Chao, Yanjie Pirzada, Zaid A Eckert, Maria R Vogel, Jörg Charpentier, (2011). CRISPR RNA maturation by trans-encoded small RNA and host factor RNase III. Umeå universitet, Umeå Centre for Microbial Research (UCMR). OCLC 1234029490. Consultado el 30 de octubre de 2022.
- Marraffini, Luciano (22 de octubre de 2013). «Faculty Opinions recommendation of CRISPR RNA maturation by trans-encoded small RNA and host factor RNase III.». Faculty Opinions – Post-Publication Peer Review of the Biomedical Literature. Consultado el 30 de octubre de 2022.
- A., Haurwitz, Rachel E. Jinek, Martin Wiedenheft, Blake Zhou, Kaihong Doudna, Jennifer (10 de septiembre de 2010). Sequence- and structure-specific RNA processing by a CRISPR endonuclease. OCLC 847380945. Consultado el 28 de octubre de 2022.
- Rath, Devashish; Amlinger, Lina; Rath, Archana; Lundgren, Magnus (2015-10). «The CRISPR-Cas immune system: biology, mechanisms and applications». Biochimie 117: 119-128. ISSN 1638-6183. PMID 25868999. doi:10.1016/j.biochi.2015.03.025. Consultado el 30 de octubre de 2022.
- Deltcheva, Elitza; Chylinski, Krzysztof; Sharma, Cynthia M.; Gonzales, Karine; Chao, Yanjie; Pirzada, Zaid A.; Eckert, Maria R.; Vogel, Jörg et al. (2011-03). «CRISPR RNA maturation by trans-encoded small RNA and host factor RNase III». Nature 471 (7340): 602-607. ISSN 0028-0836. doi:10.1038/nature09886. Consultado el 28 de octubre de 2022.
- Xiong, Xin; Chen, Meng; Lim, Wendell A.; Zhao, Dehua; Qi, Lei S. (31 de agosto de 2016). «CRISPR/Cas9 for Human Genome Engineering and Disease Research». Annual Review of Genomics and Human Genetics (en inglés) 17 (1): 131-154. ISSN 1527-8204. doi:10.1146/annurev-genom-083115-022258. Consultado el 30 de octubre de 2022.
- «Overview of the CRISPR-Cas system and design of sgRNAs (single guide RNAs) for genomic DNA editing in yeast».
- Jansen, Ruud.; Embden, Jan. D. A. van; Gaastra, Wim.; Schouls, Leo. M. (2002-03). «Identification of genes that are associated with DNA repeats in prokaryotes». Molecular Microbiology 43 (6): 1565-1575. ISSN 0950-382X. doi:10.1046/j.1365-2958.2002.02839.x. Consultado el 30 de octubre de 2022.
- «Principles and medical applications of gene editing by CRISPR / Cas».
- Thurtle-Schmidt, Deborah M.; Lo, Te-Wen (30 de enero de 2018). «Molecular biology at the cutting edge: A review on CRISPR/CAS9 gene editing for undergraduates». Biochemistry and Molecular Biology Education 46 (2): 195-205. ISSN 1470-8175. doi:10.1002/bmb.21108. Consultado el 29 de octubre de 2022.
- Munawar, Nayla; Ahmad, Aftab (2021). CRISPR/Cas System: An Introduction. Springer Singapore. pp. 1-35. ISBN 978-981-15-7141-1. Consultado el 29 de octubre de 2022.
- Khan, Sehrish; Mahmood, Muhammad Shahid; Rahman, Sajjad ur; Zafar, Hassan; Habibullah, Sultan; khan, Zulqarnain; Ahmad, Aftab (28 de marzo de 2018). «CRISPR/Cas9: the Jedi against the dark empire of diseases». Journal of Biomedical Science 25 (1). ISSN 1423-0127. doi:10.1186/s12929-018-0425-5. Consultado el 29 de octubre de 2022.
- Karvelis, Tautvydas; Gasiunas, Giedrius; Miksys, Algirdas; Barrangou, Rodolphe; Horvath, Philippe; Siksnys, Virginijus (27 de marzo de 2013). «crRNA and tracrRNA guide Cas9-mediated DNA interference inStreptococcus thermophilus». RNA Biology 10 (5): 841-851. ISSN 1547-6286. doi:10.4161/rna.24203. Consultado el 29 de octubre de 2022.
- Pingoud, Alfred (26 de julio de 2012). «Faculty Opinions recommendation of A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity.». Faculty Opinions – Post-Publication Peer Review of the Biomedical Literature. Consultado el 28 de octubre de 2022.
- Emmanuelle, Jinek, Martin Chylinski, Krzysztof Fonfara, Ines Hauer, Michael Doudna, Jennifer A Charpentier, (2012). A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Umeå universitet, Molekylär Infektionsmedicin, Sverige (MIMS). OCLC 1233892747. Consultado el 28 de octubre de 2022.
- Terns, Michael P; Terns, Rebecca M (2011-06). «CRISPR-based adaptive immune systems». Current Opinion in Microbiology 14 (3): 321-327. ISSN 1369-5274. doi:10.1016/j.mib.2011.03.005. Consultado el 30 de octubre de 2022.
- Jiang, Fuguo; Doudna, Jennifer A. (22 de mayo de 2017). «CRISPR–Cas9 Structures and Mechanisms». Annual Review of Biophysics (en inglés) 46 (1): 505-529. ISSN 1936-122X. doi:10.1146/annurev-biophys-062215-010822. Consultado el 30 de octubre de 2022.
- Friedl, Peter; Weigelin, Bettina (2014-12). «A Swiss Army Knife for CTLs». Immunity 41 (6): 873-875. ISSN 1074-7613. doi:10.1016/j.immuni.2014.12.008. Consultado el 30 de octubre de 2022.
- Emmanuelle, Jinek, Martin Chylinski, Krzysztof Fonfara, Ines Hauer, Michael Doudna, Jennifer A Charpentier, (2012). A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Umeå universitet, Molekylär Infektionsmedicin, Sverige (MIMS). OCLC 1233892747. Consultado el 30 de octubre de 2022.
- Lone, Bilal Ahmad; Karna, Shibendra Kumar Lal; Ahmad, Faiz; Shahi, Nerina; Pokharel, Yuba Raj (18 de septiembre de 2018). «CRISPR/Cas9 System: A Bacterial Tailor for Genomic Engineering». Genetics Research International 2018: 1-17. ISSN 2090-3154. doi:10.1155/2018/3797214. Consultado el 29 de octubre de 2022.
- Lüthy, Isabel A.; Lamb, Caroline A.; Lüthy, Isabel A.; Lamb, Caroline A. (2020-12). «Premio Nobel de Química 2020 a la edición génica con tecnología CRISPR/Cas9». Medicina (Buenos Aires) 80 (6): 738-740. ISSN 0025-7680. Consultado el 24 de octubre de 2022.
- «The Nobel Prize in Chemistry 2020». NobelPrize.org (en inglés estadounidense). Consultado el 24 de octubre de 2022.
- «TRABAJO FIN DE GRADO CRISPR COMO HERRAMINETA DE EDICIÓN GENÉTICA Y SUS APLICACIONES EN LA SALUD HUMANA».
- Las tijeras genéticas: premio nobel de química del 2020. Acta Herediana. doi:10.20453/ah.v64i1.3917. Consultado el 24 de octubre de 2022.
- «Premio Nobel de Química 2020 para las inventoras de las "tijeras genéticas"». www.nationalgeographic.com.es. 7 de octubre de 2020. Consultado el 24 de octubre de 2022.
- Zhang, Huimin; Qin, Chunhong; An, Changming; Zheng, Xiwang; Wen, Shuxin; Chen, Wenjie; Liu, Xianfang; Lv, Zhenghua et al. (1 de octubre de 2021). «Application of the CRISPR/Cas9-based gene editing technique in basic research, diagnosis, and therapy of cancer». Molecular Cancer 20 (1): 126. ISSN 1476-4598. PMC 8484294. PMID 34598686. doi:10.1186/s12943-021-01431-6. Consultado el 30 de octubre de 2022.
- rdu, Editor (12 de septiembre de 2021). «CRISPR-Cas: la nueva herramienta para diagnosticar enfermedades infecciosas». RDU UNAM. doi:10.22201/cuaieed.16076079e.2021.22.5.8. Consultado el 30 de octubre de 2022.
- Strich, Jeffrey R.; Chertow, Daniel S. (2019-04). «CRISPR-Cas Biology and Its Application to Infectious Diseases». Journal of Clinical Microbiology 57 (4): e01307-18. ISSN 1098-660X. PMC 6440769. PMID 30429256. doi:10.1128/JCM.01307-18. Consultado el 30 de octubre de 2022.
- «Shibboleth Authentication Request». login.sire.ub.edu. doi:10.1126/science.abe7106. Consultado el 30 de octubre de 2022.
- Soh, Jun Hui; Balleza, Enrique; Abdul Rahim, Muhammad Nadjad; Chan, Hsi-Min; Mohd Ali, Siswand; Chuah, Jacqueline Kai Chin; Edris, Sherif; Atef, Ahmed et al. (1 de noviembre de 2022). «CRISPR-based systems for sensitive and rapid on-site COVID-19 diagnostics». Trends in Biotechnology (en inglés) 40 (11): 1346-1360. ISSN 0167-7799. doi:10.1016/j.tibtech.2022.06.002. Consultado el 30 de octubre de 2022.
- «Shibboleth Authentication Request». login.sire.ub.edu. Consultado el 30 de octubre de 2022.
- Xiao, Qiaoqiao; Guo, Deyin; Chen, Shuliang (2019). «Application of CRISPR/Cas9-Based Gene Editing in HIV-1/AIDS Therapy». Frontiers in Cellular and Infection Microbiology 9: 69. ISSN 2235-2988. PMC 6439341. PMID 30968001. doi:10.3389/fcimb.2019.00069. Consultado el 30 de octubre de 2022.
- Stadtmauer, Edward A.; Fraietta, Joseph A.; Davis, Megan M.; Cohen, Adam D.; Weber, Kristy L.; Lancaster, Eric; Mangan, Patricia A.; Kulikovskaya, Irina et al. (28 de febrero de 2020). «CRISPR-engineered T cells in patients with refractory cancer». Science (New York, N.Y.) 367 (6481): eaba7365. ISSN 1095-9203. PMID 32029687. doi:10.1126/science.aba7365. Consultado el 30 de octubre de 2022.
Enlaces externos
- Más información sobre el ARNtracr
- RCSB PDB - 6K4P: Crystal structure of xCas9 in complex with sgRNA and DNA (TGG PAM)
- Tenacibaculum maritimum strain Tur45 class II type CRISPR array repeat - Nucleotide - NCBI (nih.gov)
- 5VZL: cryo-EM structure of the Cas9-sgRNA-AcrIIA4 anti-CRISPR complex
- NC_002737.2: Streptococcus pyogenes M1 GAS, complete sequence (nih.gov)
- CRISPR
- Fases del sistema CRISPR/Cas
- Terminación independiente de Rho
- Predicción de estructura secundaria de ARN mediante SHAPE
- Sondeo de ADN o ARN
- RNAsaIII
- Inmunidad adaptativa o adquirida